Endotoxin Translocation Is Increased in Broiler Chickens Fed a Fusarium Mycotoxin-Contaminated Diet

 ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Growth Performance
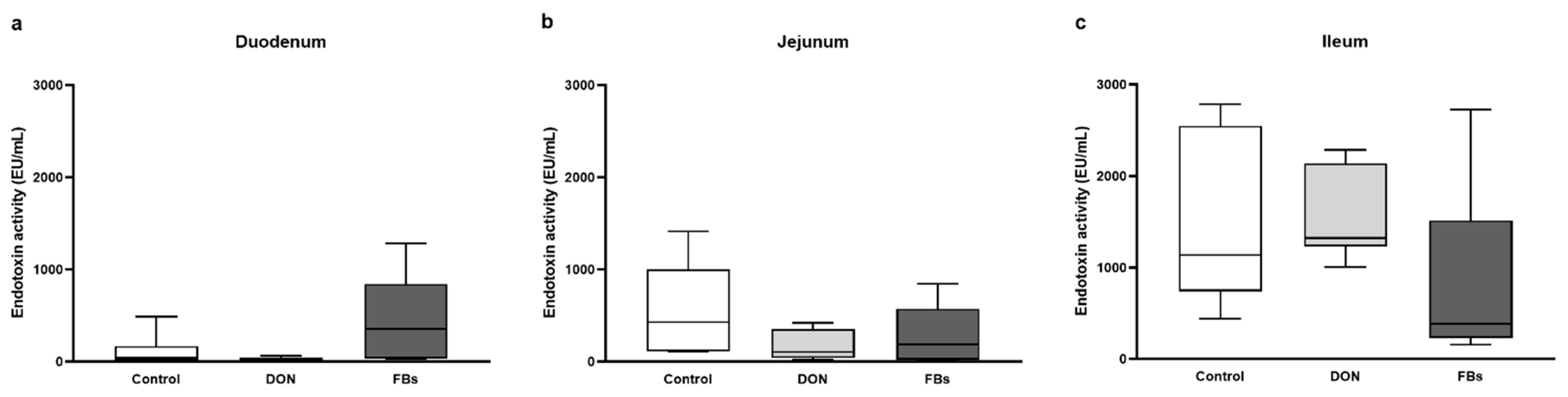
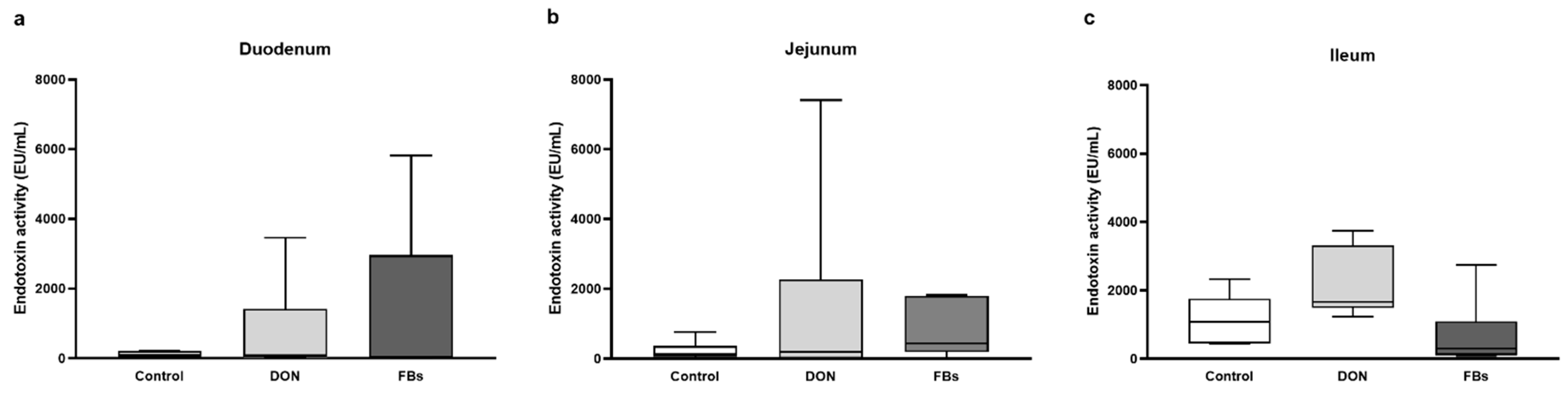
2.2. Thermoneutral Conditions—Endotoxin Activity of Intestinal Digesta
2.3. Thermoneutral Conditions—Gene Expression of Intestinal Tissue
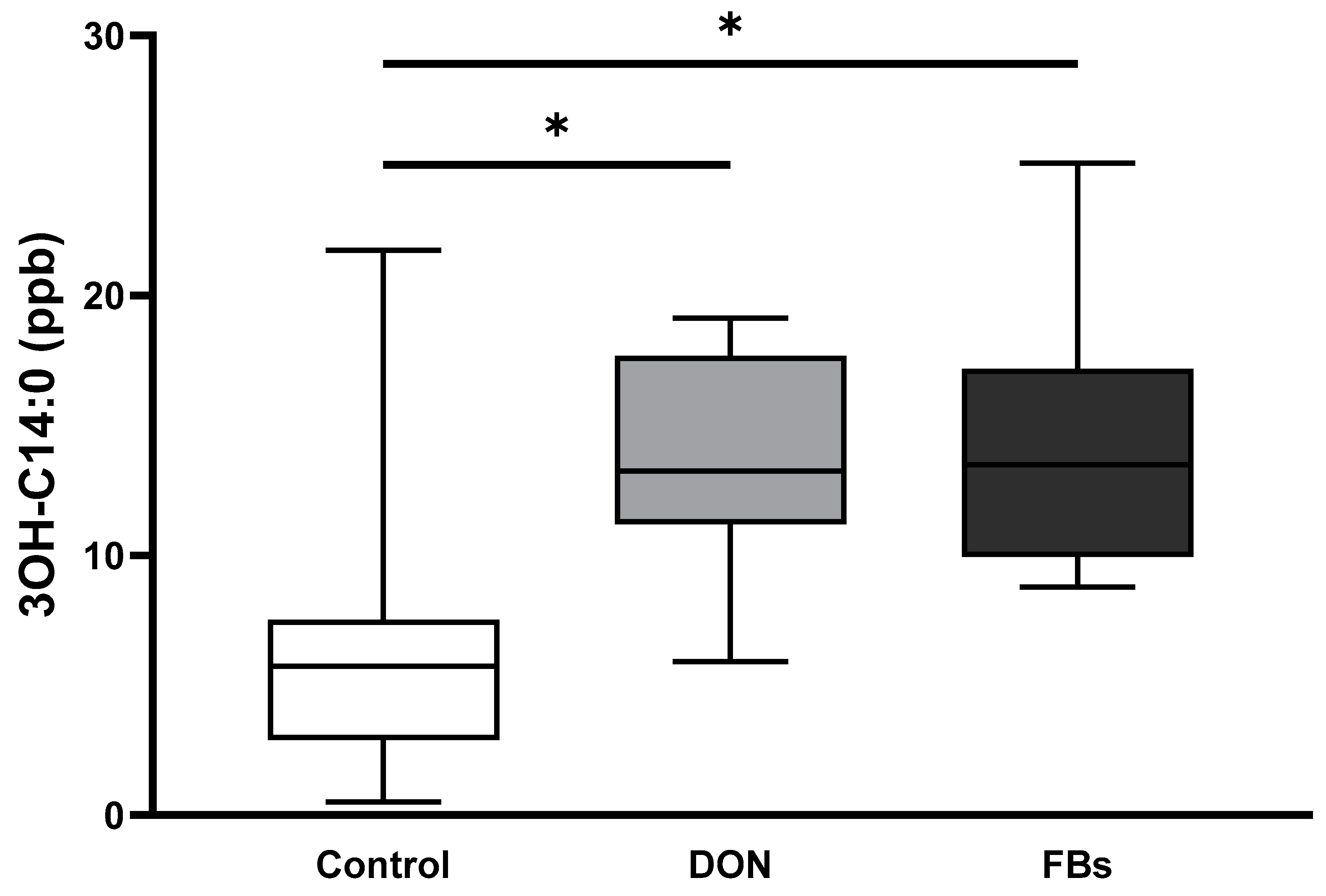
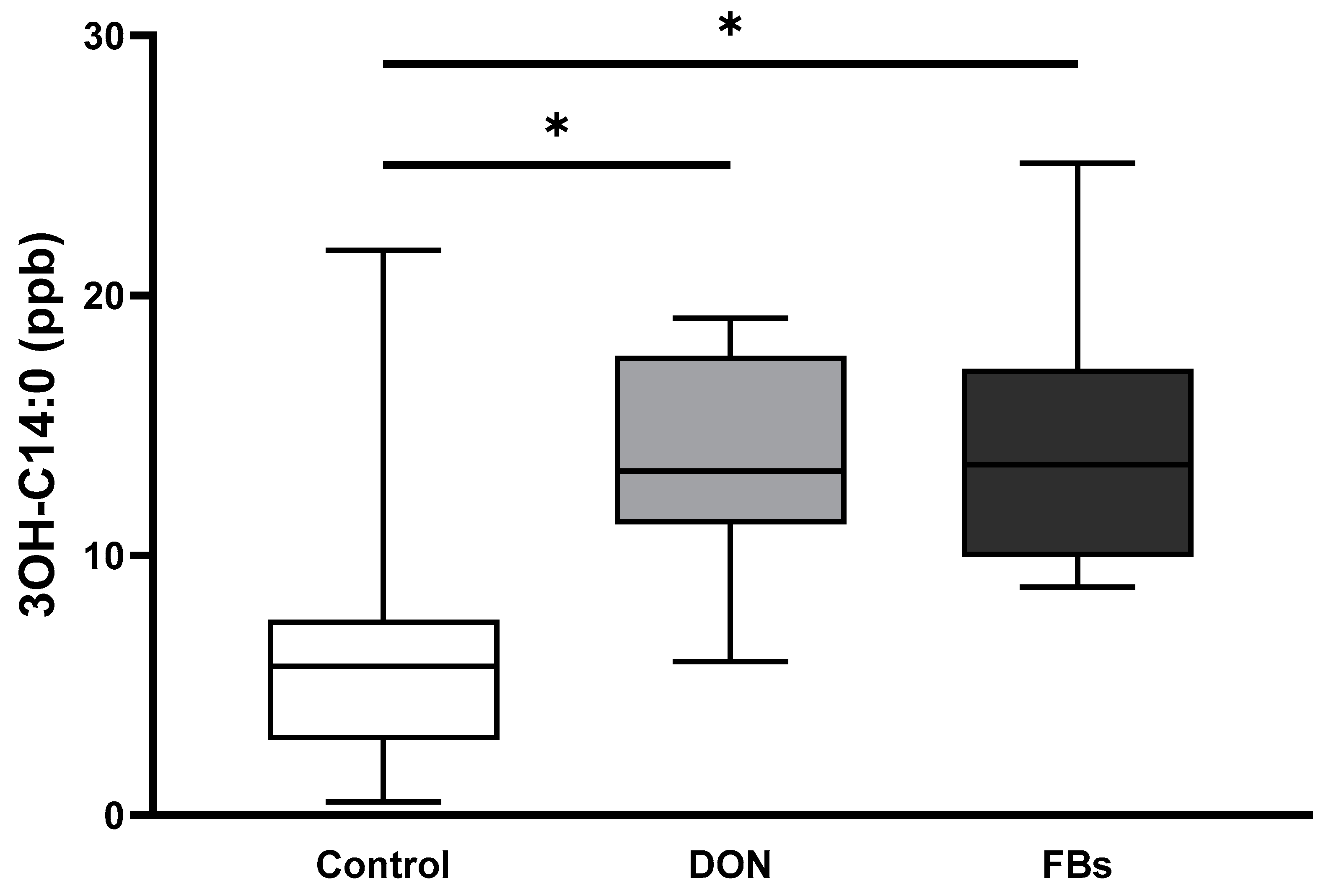
2.4. Thermoneutral Conditions—Endotoxin Marker (3-OH C14:0)
2.5. Heat Stress Conditions—Endotoxin Activity of Intestinal Digesta
2.6. Heat Stress Conditions—Gene Expression of Intestinal Tissue
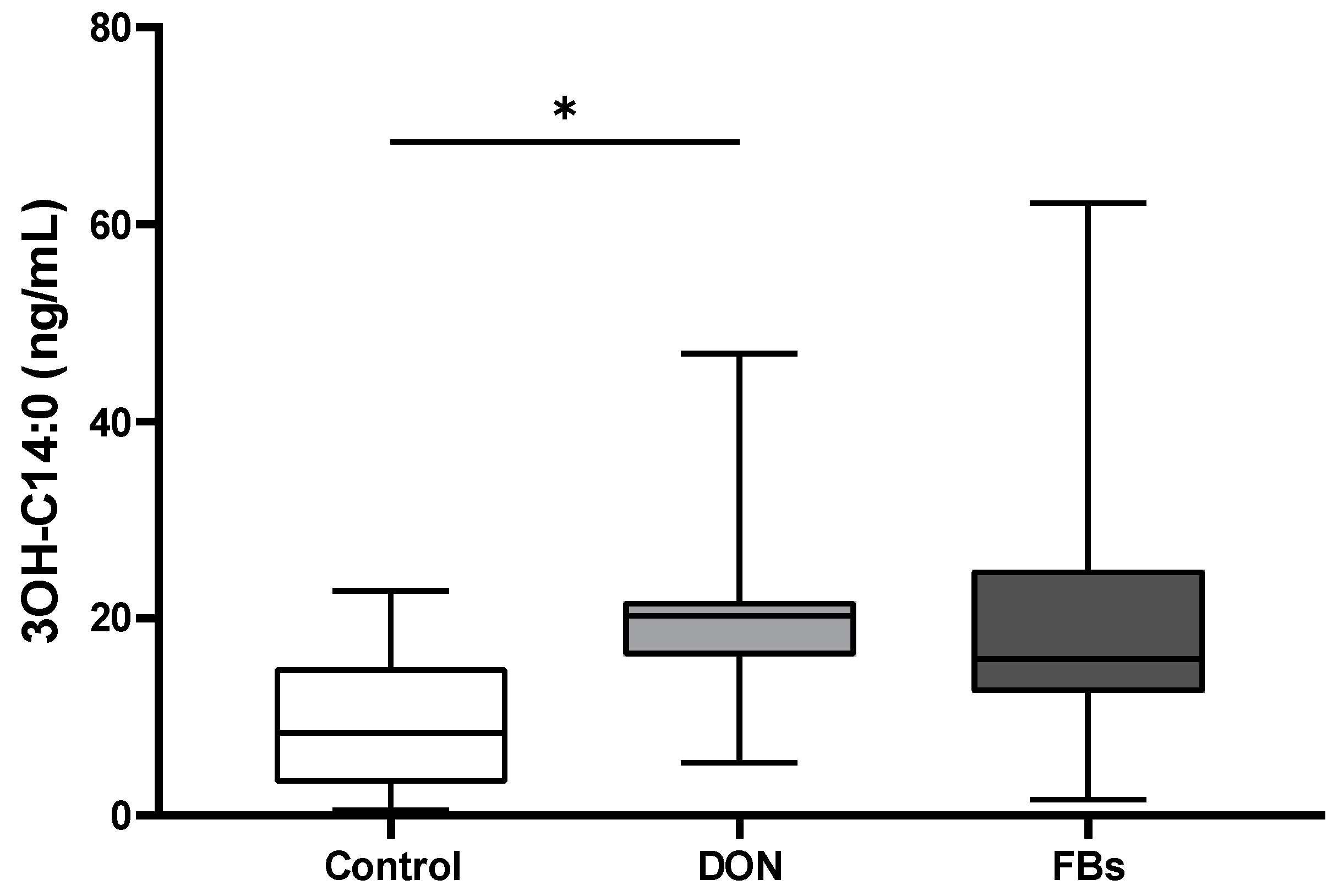
2.7. Heat Stress Conditions—3-OH C14:0 Plasma Concentration
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethic Statement
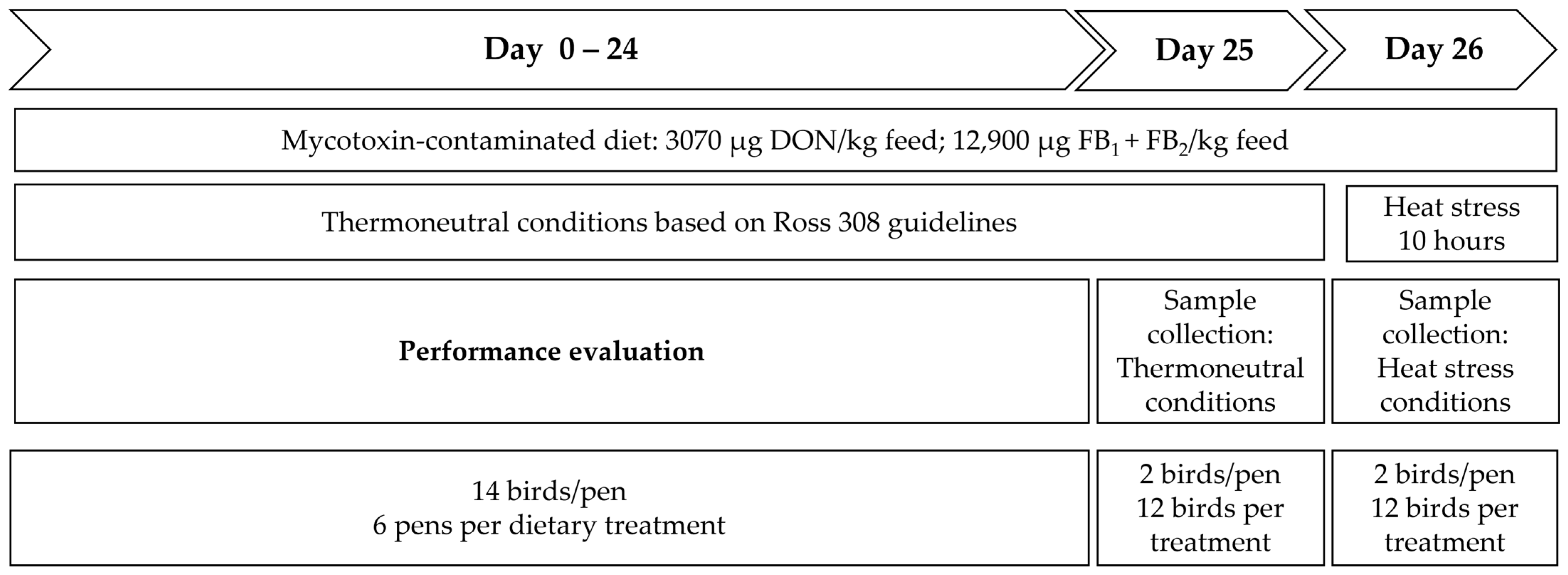
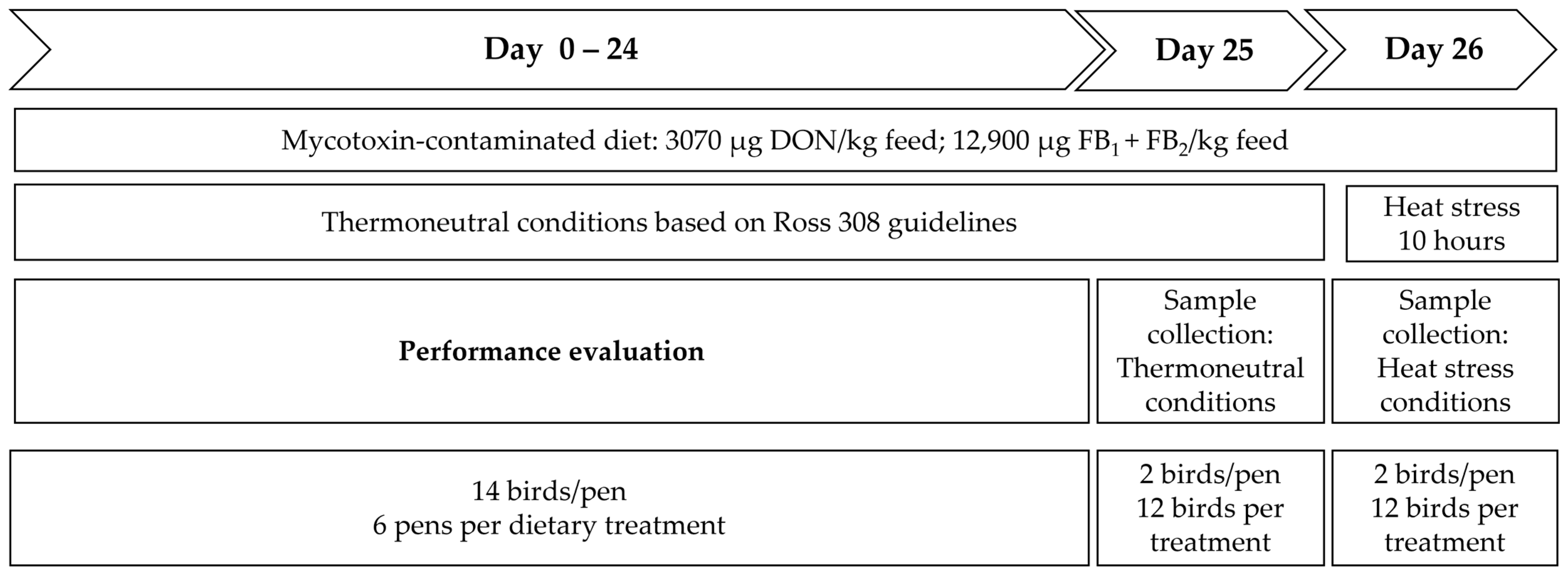
5.2. Animals and Trial Set-Up
5.3. Sampling
5.4. Analysis
5.4.1. Endotoxin Analysis Gastrointestinal Tract
5.4.2. RNA Extraction and Gene Expression Analysis of the Intestinal Tissue
5.4.3. Plasma Endotoxin Marker (3-OH C14:0)
6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kolawole, O.; Graham, A.; Donaldson, C.; Owens, B.; Abia, W.A.; Meneely, J.; Alcorn, M.J.; Connolly, L.; Elliott, C.T. Low Doses of Mycotoxin Mixtures below EU Regulatory Limits Can Negatively A Ff Ect the Performance of Broiler Chickens: A Longitudinal Study. Toxins 2020, 12, 433. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.D.; Doupovec, B.; Schatzmayr, D.; Murugesan, G.R.; Bortoluzzi, C.; Villegas, A.M.; Applegate, T.J. The Impact of Deoxynivalenol, Fumonisins, and Their Combination on Performance, Nutrient, and Energy Digestibility in Broiler Chickens. Poult. Sci. 2019, 99, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Dohnal, I.; Shanmugasundaram, R.; Eicher, S.D.; Selvaraj, R.K.; Schatzmayr, G.; Applegate, T.J. Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction. Toxins 2016, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Paraskeuas, V.; Griela, E.; Bouziotis, D.; Fegeros, K.; Antonissen, G.; Mountzouris, K.C. Effects of Deoxynivalenol and Fumonisins on Broiler Gut Cytoprotective Capacity. Toxins 2021, 13, 729. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; del Mazo, J.K.C.J.; Grasl-Kraupp, B.; Hogstrand, C.; Leblanc, J.C.; Nielsen, E.; Ntzani, E.; et al. Assessment of Information as Regards the Toxicity of Deoxynivalenol for Horses and Poultry. EFSA J. 2023, 21. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Leblanc, J.C.; Nielsen, E.; et al. Assessment of Information as Regards the Toxicity of Fumonisins for Pigs, Poultry and Horses. EFSA J. 2022, 20. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Tabler, T.W.; Greene, E.S.; Orlowski, S.K.; Hiltz, J.Z.; Anthony, N.B.; Dridi, S. Intestinal Barrier Integrity in Heat-Stressed Modern Broilers and Their Ancestor Wild Jungle Fowl. Front. Vet. Sci. 2020, 7, 249. [Google Scholar] [CrossRef]
- Rostagno, M.H. Effects of Heat Stress on the Gut Health of Poultry. J. Anim. Sci. 2020, 98, skaa090. [Google Scholar] [CrossRef]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken Hepatic Response to Chronic Heat Stress Using Integrated Transcriptome and Metabolome Analysis. PLoS ONE 2017, 12, e0181900. [Google Scholar] [CrossRef]
- Kang, D.; Shim, K. Early Heat Exposure Effects on Proteomic Changes of The Broiler Liver under Acute Heat Stress. Animals 2021, 11, 1338. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide Endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- De Boever, S.; Croubels, S.; Meyer, E.; Sys, S.; Beyaert, R.; Ducatelle, R.; De Backer, P. Characterization of an Intravenous Lipopolysaccharide Inflammation Model in Broiler Chickens. Avian Pathol. 2009, 38, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Berczi, I.; Bertok, L.; Bereznai, T. Comparative Studies on the Toxicity of Escherichia Coli Lipopolysaccharide Endotoxin in Various Animal Species. Can. J. Microbiol. 1966, 12, 1070–1071. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Nighot, M.; Al-Sadi, R.; Alhmoud, T.; Nighot, P.; Ma, T.Y. Lipopolysaccharide Regulation of Intestinal Tight Junction Permeability Is Mediated by TLR4 Signal Transduction Pathway Activation of FAK and MyD88. J. Immunol. 2015, 195, 4999–5010. [Google Scholar] [CrossRef] [PubMed]
- Nighot, M.; Rawat, M.; Al-Sadi, R.; Castillo, E.F.; Nighot, P.; Ma, T.Y. Lipopolysaccharide-Induced Increase in Intestinal Permeability Is Mediated by TAK-1 Activation of IKK and MLCK/MYLK Gene. Am. J. Pathol. 2019, 189, 797–812. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zhang, Q.; Wang, C.C.; Wu, H.; Jiao, L.; Hong, Q.; Hu, C. LPS Challenge Increased Intestinal Permeability, Disrupted Mitochondrial Function and Triggered Mitophagy of Piglets. Innate Immun. 2018, 24, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Nazar, F.N.; Estevez, I. The Immune-Neuroendocrine System, a Key Aspect of Poultry Welfare and Resilience. Poult. Sci. 2022, 101, 101919. [Google Scholar] [CrossRef]
- Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N.; et al. Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets And IPEC-J2 Cells. Toxins 2018, 15, 541. [Google Scholar] [CrossRef]
- Kahlert, S.; Renner, L.; Kluess, J.; Frahm, J.; Tesch, T.; Bannert, E.; Kersten, S.; Dänicke, S.; Rothkötter, H.J. Effects of Deoxynivalenol-Feed Contamination on Circulating LPS in Pigs. Innate Immun. 2019, 5, 168–175. [Google Scholar] [CrossRef]
- Skiepko, N.; Przybylska-Gornowicz, B.; Gajęcka, M.; Gajęcki, M.; Lewczuk, B. Effects of Deoxynivalenol and Zearalenone on The Histology and Ultrastructure of Pig Liver. Toxins 2020, 20, 463. [Google Scholar] [CrossRef] [PubMed]
- Munford, R.S. Invited Review Detoxifying Endotoxin: Time, Place and Person. J. Endotoxin. Res. 2005, 11, 69–84. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.J.; Fernandez, M.V.S.; Baumgard, L.H. Glucose Requirements of an Activated Immune System in Lactating Holstein Cows. J. Dairy. Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Mayorga, E.J.; Sanz-Fernandez, M.V.; Abuajamieh, M.; Baumgard, L.H. Estimating Glucose Requirements of an Activated Immune System in Growing Pigs. J. Anim. Sci. 2017, 95, 5020–5029. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Wang, F.; Ma, H.; Ren, Z.; Yang, X.; Yang, X. Study on the Interactive Effect of Deoxynivalenol and Clostridium Perfringens on the Jejunal Health of Broiler Chickens. Poult. Sci. 2020, 100, 100807. [Google Scholar] [CrossRef]
- Lucke, A.; Böhm, J.; Zebeli, Q.; Metzler-Zebeli, B.U. Dietary Deoxynivalenol Contamination and Oral Lipopolysaccharide Challenge Alters the Cecal Microbiota of Broiler Chickens. Front. Microbiol. 2018, 9, 804. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Croubels, S.; Pasmans, F.; Ducatelle, R.; Eeckhaut, V.; Devreese, M.; Verlinden, M.; Haesebrouck, F.; Eeckhout, M.; De Saeger, S.; et al. Fumonisins Affect the Intestinal Microbial Homeostasis in Broiler Chickens, Predisposing to Necrotic Enteritis. Vet. Res. 2015, 46, 98. [Google Scholar] [CrossRef]
- Humer, E.; Kröger, I.; Neubauer, V.; Schedle, K.; Reisinger, N.; Zebeli, Q. Supplementing Phytogenic Compounds or Autolyzed Yeast Modulates Ruminal Biogenic Amines and Plasma Metabolome in Dry Cows Experiencing Subacute Ruminal Acidosis. J. Dairy. Sci. 2018, 101, 9559–9574. [Google Scholar] [CrossRef]
- Huo, W.; Zhu, W.; Mao, S. Effects of Feeding Increasing Proportions of Corn Grain on Concentration of Lipopolysaccharide in the Rumen Fluid and the Subsequent Alterations in Immune Responses in Goats. Asian-Australas. J. Anim. Sci. 2013, 26, 1437–1445. [Google Scholar] [CrossRef]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. Alfalfa Pellet-Induced Subacute Ruminal Acidosis in Dairy Cows Increases Bacterial Endotoxin in the Rumen without Causing Inflammation. J. Dairy. Sci. 2009, 92, 1712–1724. [Google Scholar] [CrossRef]
- Emsenhuber, C.; Reisinger, N.; Mayer, E.; Schatzmayr, G. Endotoxin Concentrations in the Gut Increase from the Duodenum to the Ileum in Healthy Broiler Chickens. In Proceedings of the EEAP Annual Meeting, Belfast, UK, 29 August–2 September 2016; p. 246, Session 20 Nutrition physiology. [Google Scholar]
- Reisinger, N.; Emsenhuber, C.; Doupovec, B.; Mayer, E.; Schatzmayr, G.; Nagl, V.; Grenier, B. Endotoxin Translocation and Gut Inflammation Are Increased in Broiler Chickens Receiving an Oral Lipopolysaccharide (LPS) Bolus during Heat Stress. Toxins 2020, 12, 622. [Google Scholar] [CrossRef]
- Liu, W.; Huang, M.; Balasubramanian, B.; Jha, R. Heat Stress Affects Jejunal Immunity of Yellow-Feathered Broilers and Is Potentially Mediated by the Microbiome. Front. Physiol. 2022, 13, 913696. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Bai, L.; Qu, Q.; Zhou, S.; Yang, M.; Guo, S.; Li, Q.; Liu, C. Impact of Gut Microbiota Structure in Heat-Stressed Broilers. Poult. Sci. 2018, 98, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Sarmikasoglou, E.; Vinyard, J.R.; Khan, M.S.; Jiranantasak, T.; Ravelo, A.; Lobo, R.R.; Fan, P.; Jeong, K.C.; Tuanyok, A.; Faciola, A. Ruminal Lipid A Analysis by Matrix-Assisted Laser Desorption / Ionization Time-of-Flight Mass Spectrometry. Polysaccharides 2021, 2, 817–824. [Google Scholar] [CrossRef]
- Lucke, A.; Böhm, J.; Zebeli, Q.; Metzler-Zebeli, B. Dietary Deoxynivalenol and Oral Lipopolysaccharide Challenge Differently Affect Intestinal Innate Immune Response and Barrier Function in Broiler Chickens. J. Anim. Sci. 2018, 96, 5134–5143. [Google Scholar] [CrossRef]
- Azizi, T.; Daneshyar, M.; Allymehr, M.; Shalizar, A.; Khalilvandi, H.; Tukmechi, A. The Impact of Deoxynivalenol Contaminated Diet on Performance, Immune Response, Intestine Morphology and Jejunal Gene Expression in Broiler Chicken. Toxicon 2021, 199, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.J.; De Baere, S.; Mountzouris, K.C.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins Deoxynivalenol and Fumonisins Alter the Extrinsic Component of Intestinal Barrier in Broiler Chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Böhm, J.; Zebeli, Q. Impacts of the Feed Contaminant Deoxynivalenol on the Intestine of Monogastric Animals. Poultry and Swine. J. Appl. Toxicol. 2014, 35, 327–337. [Google Scholar] [CrossRef]
- Lichte, P.; Grigoleit, J.S.; Steiner, E.M.; Kullmann, J.S.; Schedlowski, M.; Oberbeck, R.; Kobbe, P. Low Dose LPS Does Not Increase TLR4 Expression on Monocytes in a Human in Vivo Model. Cytokine 2013, 63, 74–80. [Google Scholar] [CrossRef]
- Jiang, K.; Guo, S.; Zhang, T.; Yang, Y.; Zhao, G.; Shaukat, A.; Wu, H.; Deng, G. Downregulation of TLR4 by miR-181a Provides Negative Feedback Regulation to Lipopolysaccharide-Induced Inflammation. Front. Pharmacol. 2018, 26, 142. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porc. Health Manag. 2016, 14, 21. [Google Scholar] [CrossRef]
- Bouhet, S.; Le Dorze, E.; Peres, S.; Fairbrother, J.M.; Oswald, I.P. Mycotoxin fumonisin B1 selectively down-regulates the basal IL-8 expression in pig intestine: In vivo and in vitro studies. Food Chem. Toxicol. 2006, 44, 1768–1773. [Google Scholar] [CrossRef]
- Becker, C.; Reiter, M.; Pfaffl, M.W.; Meyer, H.H.D.; Bauer, J.; Meyer, K.H.D. Expression of Immune Relevant Genes in Pigs under the Influence of Low Doses of Deoxynivalenol (DON). Mycotoxin Res. 2011, 27, 287–293. [Google Scholar] [CrossRef]
- Pais de Barros, J.; Gautier, T.; Sali, W.; Adrie, C.; Choubley, H.; Charron, E.; Lalande, C.; Le Guern, N.; Deckert, V.; Monchi, M.; et al. Quantitative Lipopolysaccharide Analysis Using HPLC / MS / MS and Its Combination with the Limulus Amebocyte Lysate Assay. J. Lipid Res. 2015, 56, 1363–1369. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, X.; Hsieh, J.C.F.; Monson, M.S.; Zhang, J.; Shu, D.; Nie, Q.; Persia, M.E.; Rothschild, M.F.; Lamont, S.J. Transcriptome Response of Liver and Muscle in Heat-Stressed Laying Hens. Genes. 2021, 12, 255. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Hsieh, J.C.F.; Schmidt, C.J.; Zhu, Q.; Lamont, S.J. Liver Transcriptome Response to Hyperthermic Stress in Three Distinct Chicken Lines. BMC Genom. 2016, 17, 955. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. QBase Relative Quantification Framework and Software for Management and Automated Analysis of Real-Time Quantitative PCR Data. Genome Biol. 2008, 8, R19. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Body Weight Gain (g) | ||
|---|---|---|---|---|
| Day 1 | Day 1–12 | Day 12–24 | Day 1–24 | |
| Control | 43 ± 0 | 270 ± 8 a | 770 ± 21 | 1040 ± 29 a |
| DON | 43 ± 0 | 235 ± 8 b | 742 ± 12 | 977 ± 16 ab |
| FBs | 43 ± 1 | 243 ± 7 ab | 707 ± 22 | 950 ± 25 b |
| Gut Barrier Function | |||||||||||||||
| Mid-Duodenum | Mid-Jejunum | Mid-Ileum | |||||||||||||
| CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | |
| Control | 0.72 ± 0.318 | 0.72 ± 0.142 | 0.83 ± 0.375 | 0.93 ± 0.1851 | 0.88± 0.279 | 0.76 ± 0.172 | 0.72 ± 0.177 a | 0.83 ± 0.523 a | 0.96 ± 0.814 | 0.89 ± 0.404 a | 0.86 ± 0.096 | 0.85 ± 0.042 | 1.03 ± 0.533 | 0.90 ± 0.158 | 0.83 ± 0.079 x |
| DON | 1.45 ± 0.906 | 0.93 ± 0.184 | 1.092 ± 0.299 | 1.068 ± 0.332 | 0.87 ± 0.172 | 0.60 ± 0.445 | 0.51 ± 0.254 ab | 0.51 ± 0.206 ab | 1.00 ± 0.581 | 0.30 ± 0.109 ab | 0.86 ± 0.431 | 0.83 ± 0.153 | 0.87 ± 0.451 | 0.95 ± 0.198 | 1.13 ± 0.265 y |
| FBs | 1.14 ± 0.622 | 0.99 ± 0.562 | 0.80± 0.260 | 0.74 ± 0.108 | 0.71 ± 0.308 | 0.47 ± 0.216 | 0.27 ± 0.166 b | 0.24 ± 0.167 b | 0.89 ± 1.065 | 0.22 ± 0.100 b | 0.99 ± 0.296 | 1.11 ± 0.378 | 1.32 ± 0.371 | 1.05 ± 0.887 | 1.17 ± 0.449 xy |
| p-value | 0.1946 | 0.2029 | 0.2377 | 0.0629 | 0.4439 | 0.3820 | 0.0101 | 0.0184 | 0.7996 | 0.0022 | 0.3346 | 0.5585 | 0.2427 | 0.8017 | 0.0803 |
| Inflammatory Response | |||||||||||||||
| Mid-Duodenum | Mid-Jejunum | Mid-Ileum | |||||||||||||
| TLR2 | IL6 | IL8 | TLR2 | TLR4 | IL1b | IL6 | IL8 | TLR2 | TLR4 | IL1b | IL6 | IL8 | |||
| Control | 0.54 ± 0.214 | 0.30 ± 0.118 | 0.98 ± 1.079 | 0.60 ± 0.310 | 1.14 ± 0.269 a | 0.52 ± 0.334 | 0.49 ± 0.294 | 0.65 ± 0.534 | 0.75 ± 0.287 | 1.01 ± 0.252 | 0.768 ± 0.634 | 1.02 ± 0.609 | 0.57 ± 0.155 | ||
| DON | 1.84 ± 1.993 | 5.00 ± 7.366 | 0.59 ± 0.196 | 0.97 ± 0.666 | 0.39 ± 0.092 ab | 1.24 ± 1.541 | 1.45 ± 1.393 | 0.88 ± 0.911 | 1.04 ± 0.676 | 0.97 ± 0.220 | 0.88 ± 0.607 | 1.69 ± 2.364 | 1.30 ± 0.756 | ||
| FBs | 2.173 ± 2.761 | 4.18 ± 6.982 | 1.25 ± 1.356 | 0.40 ± 0.237 | 0.23 ± 0.128 b | 0.77 ± 1.047 | 0.88 ± 0.683 | 0.46 ± 0.320 | 0.83 ± 0.467 | 1.02 ± 0.362 | 0.39 ± 0.160 | 0.66 ± 0.412 | 0.83 ± 0.641 | ||
| p-value | 0.1664 | 0.0519 | 0.9690 | 0.1390 | 0.0002 | 0.4018 | 0.3493 | 0.8508 | 0.8850 | 0.9006 | 0.1743 | 0.7251 | 0.4786 | ||
| Gut Barrier Function | |||||||||||||||
| Mid-Duodenum | Mid-Jejunum | Mid-Ileum | |||||||||||||
| CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | CLDN1 | CLDN5 | OCLN | ZO1 | ZO2 | |
| Control | 0.69 ± 0.287 a | 0.748 ± 0.379 | 0.80 ± 0.208 | 1.30 ± 0.672 | 1.27 ± 0.736 | 1.43 ± 2.078 | 1.45 ± 1.205 a | 0.97 ± 0.399 x | 1.32 ± 0.625 | 1.28 ± 0.572 a | 0.70 ± 0.225 | 0.76 ± 0.286 x | 0.82 ± 0.396 | 0.90 ± 0.326 | 0.86 ± 0.205 x |
| DON | 0.88 ± 0.427 ab | 0.96 ± 0.656 | 0.90 ± 0.200 | 0.97 ± 0.260 | 1.19 ± 0.488 | 0.50 ± 0.113 | 0.55 ± 0.243 ab | 0.65 ± 0.321 xy | 1.84 ± 0.587 | 0.65 ± 0.395 ab | 0.78 ± 0.174 | 1.04 ± 0.271 xy | 1.14 ± 0.441 | 1.15 ± 0.376 | 1.02 ± 0286 xy |
| FBs | 1.44 ± 0.594 b | 1.21 ± 0.103 | 1.58 ± 0.780 | 1.10 ± 0.348 | 1.39 ± 0.272 | 0.46 ± 0.242 | 0.28 ± 0.090 b | 0.40 ± 0.216 y | 1.03 ± 0.905 | 0.51 ± 0.108 b | 1.08 ± 0.776 | 1.15 ± 0.391 y | 1.33 ± 0.741 | 1.07 ± 0.626 | 1.74 ± 1.035 y |
| p-value | 0.0295 | 0.1680 | 0.1630 | 0.6374 | 0.3405 | 0.6183 | 0.0039 | 0.0528 | 0.6580 | 0.0211 | 0.5929 | 0.0574 | 0.2931 | 0.6219 | 0.0509 |
| Inflammatory Response | |||||||||||||||
| Mid-Duodenum | Mid-Jejunum | Mid-Ileum | |||||||||||||
| TLR2 | IL6 | IL8 | TLR2 | TLR4 | IL1b | IL6 | IL8 | TLR2 | TLR4 | IL1b | IL6 | IL8 | |||
| Control | 0.567 ± 0.338 a | 0.32 ± 0.148 a | 2.26 ± 2.371 | 1.51 ± 1.801 x | 0.95 ± 0.426 a | 1.43 ± 1.067 a | 3.29 ± 5.793 | 2.62 ± 2.018 a | 0.905 ± 1.173 | 0.69 ± 0.148 a | 0.67 ± 0.395 | 1.30 ± 1.916 | 1.22 ± 0.485 | ||
| DON | 0.812 ± 0.541 ab | 1.18 ± 1.723 ab | 3.52 ± 3.712 | 0.48 ± 0.189 xy | 0.67 ± 0.374 ab | 0.51 ± 0.302 ab | 0.57 ± 0.48 | 0.65 ± 0.265 b | 0.570 ± 0.580 | 1.04 ± 0.232 ab | 2.83 ± 3.92 | 2.01 ± 1.950 | 1.67 ± 0.862 | ||
| FBs | 1.68 ± 0.862 b | 3.65 ± 2.909 b | 1.18 ± 0.761 | 0.35 ± 0.228 y | 0.40 ± 0.099 b | 0.26 ± 0.180 b | 0.56 ± 0.676 | 0.89 ± 0.463 ab | 0.91 ± 1.173 | 1.84 ± 1.210 b | 0.52 ± 0.378 | 0.68 ± 0.384 | 1.96 ± 1.081 | ||
| p-value | 0.0174 | 0.0208 | 0.4570 | 0.0802 | 0.0074 | 0.0168 | 0.9823 | 0.0312 | 0.5487 | 0.0003 | 0.1790 | 0.8040 | 0.3022 | ||
| Selected Genes | NCBI Reference Sequence |
|---|---|
| Gut barrier function | |
| Claudin 1 (CLDN1) | NM_001013611 |
| Claudin 5 (CLDN5) | NM_204201 |
| Occludin (OCLN) | XM_025144248 |
| Zonula occludens 1 (ZO1) | XM_015278981 |
| Zonula occludens 2 (ZO2) | XM_025144669 |
| Inflammation | |
| Toll-like receptor 2 (TLR2) | XM_0469144 |
| Toll-like receptor 4 (TLR4) | NM_001030693 |
| Interleukin 1-beta (IL1β) | XM_046931582 |
| Interleukin 6 (IL6) | NM_204628 |
| Interleukin 8 (IL8) | NM_205498 |
| Housekeeping genes | |
| Beta actin (ACTB) | NM_205518 |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | NM_204305 |
| Glucose 6-phosphate dehydrogenase (G6PD) | M11100 |
| Ribosomal protein L7 (RPL7) | XM_046910609 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reisinger, N.; Doupovec, B.; Czabany, T.; Van Immerseel, F.; Croubels, S.; Antonissen, G. Endotoxin Translocation Is Increased in Broiler Chickens Fed a Fusarium Mycotoxin-Contaminated Diet. Toxins 2024, 16, 167. https://doi.org/10.3390/toxins16040167
Reisinger N, Doupovec B, Czabany T, Van Immerseel F, Croubels S, Antonissen G. Endotoxin Translocation Is Increased in Broiler Chickens Fed a Fusarium Mycotoxin-Contaminated Diet. Toxins. 2024; 16(4):167. https://doi.org/10.3390/toxins16040167
Chicago/Turabian StyleReisinger, Nicole, Barbara Doupovec, Tibor Czabany, Filip Van Immerseel, Siska Croubels, and Gunther Antonissen. 2024. "Endotoxin Translocation Is Increased in Broiler Chickens Fed a Fusarium Mycotoxin-Contaminated Diet" Toxins 16, no. 4: 167. https://doi.org/10.3390/toxins16040167
APA StyleReisinger, N., Doupovec, B., Czabany, T., Van Immerseel, F., Croubels, S., & Antonissen, G. (2024). Endotoxin Translocation Is Increased in Broiler Chickens Fed a Fusarium Mycotoxin-Contaminated Diet. Toxins, 16(4), 167. https://doi.org/10.3390/toxins16040167