Venom Composition of Neglected Bothropoid Snakes from the Amazon Rainforest: Ecological and Toxinological Implications

 , , , , , , ,
, , , , , , ,  ,
, 
Abstract
1. Introduction
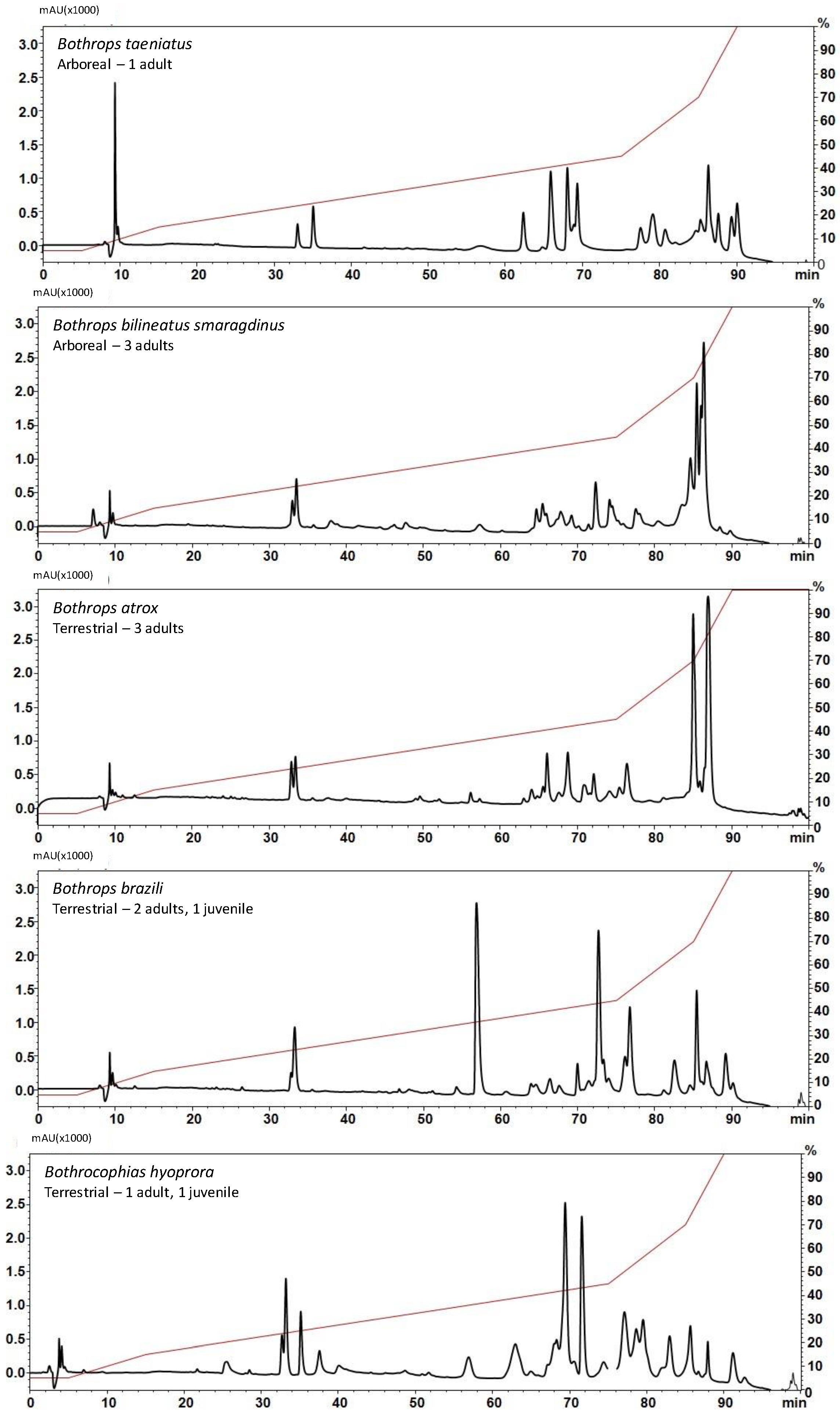
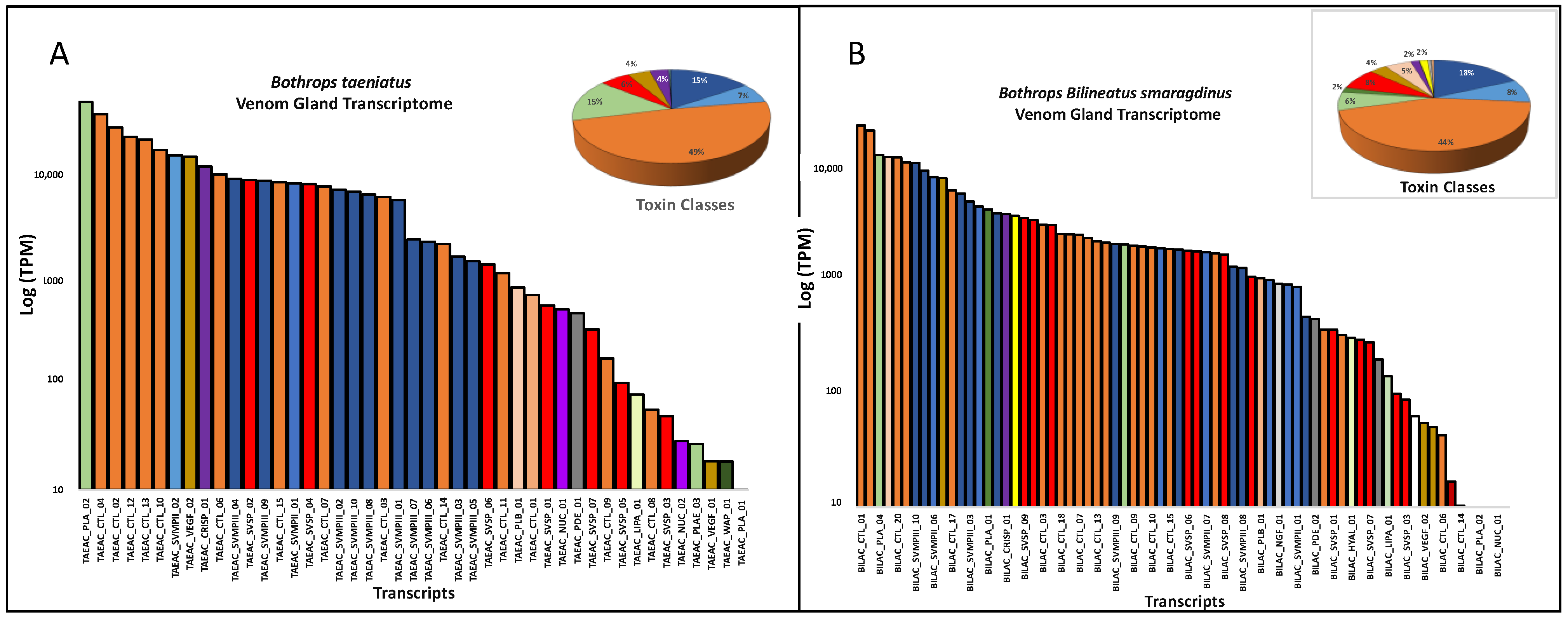
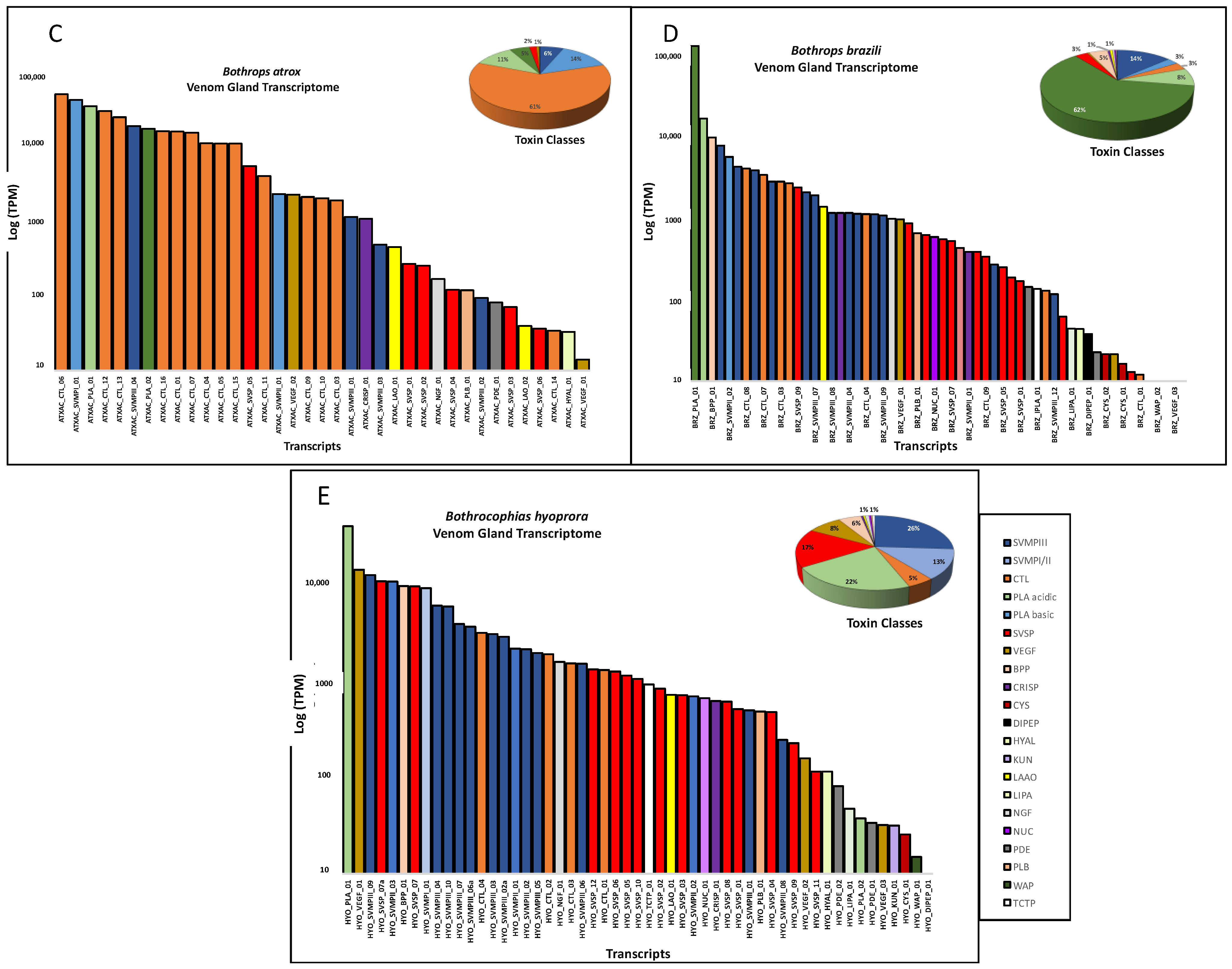
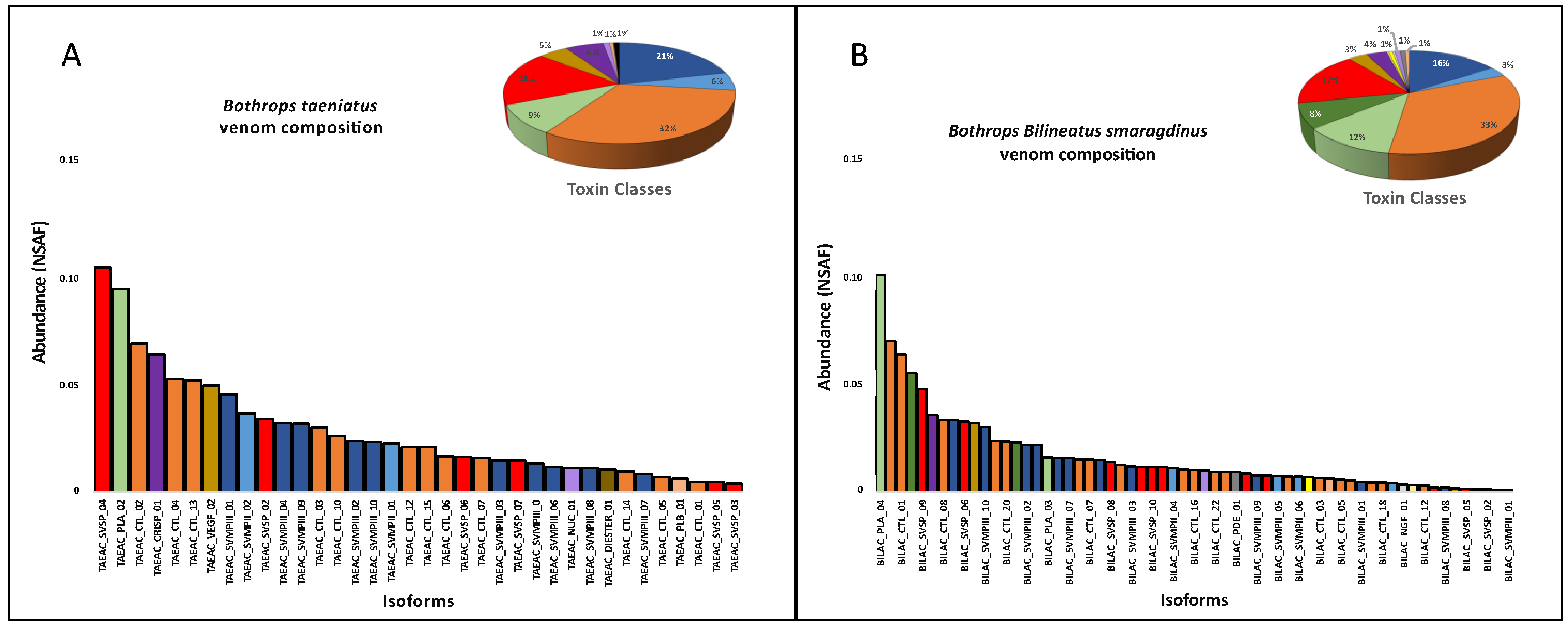
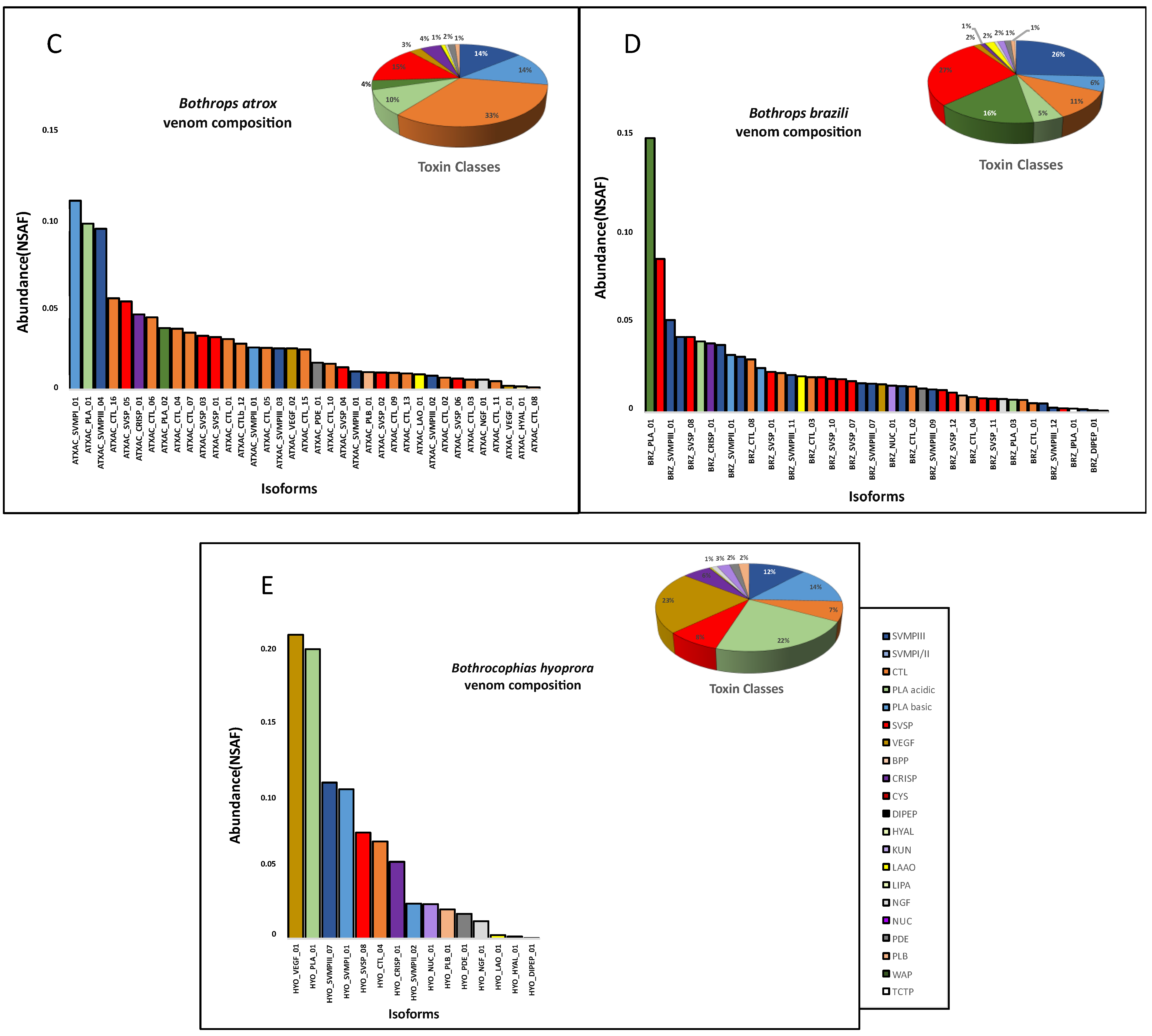
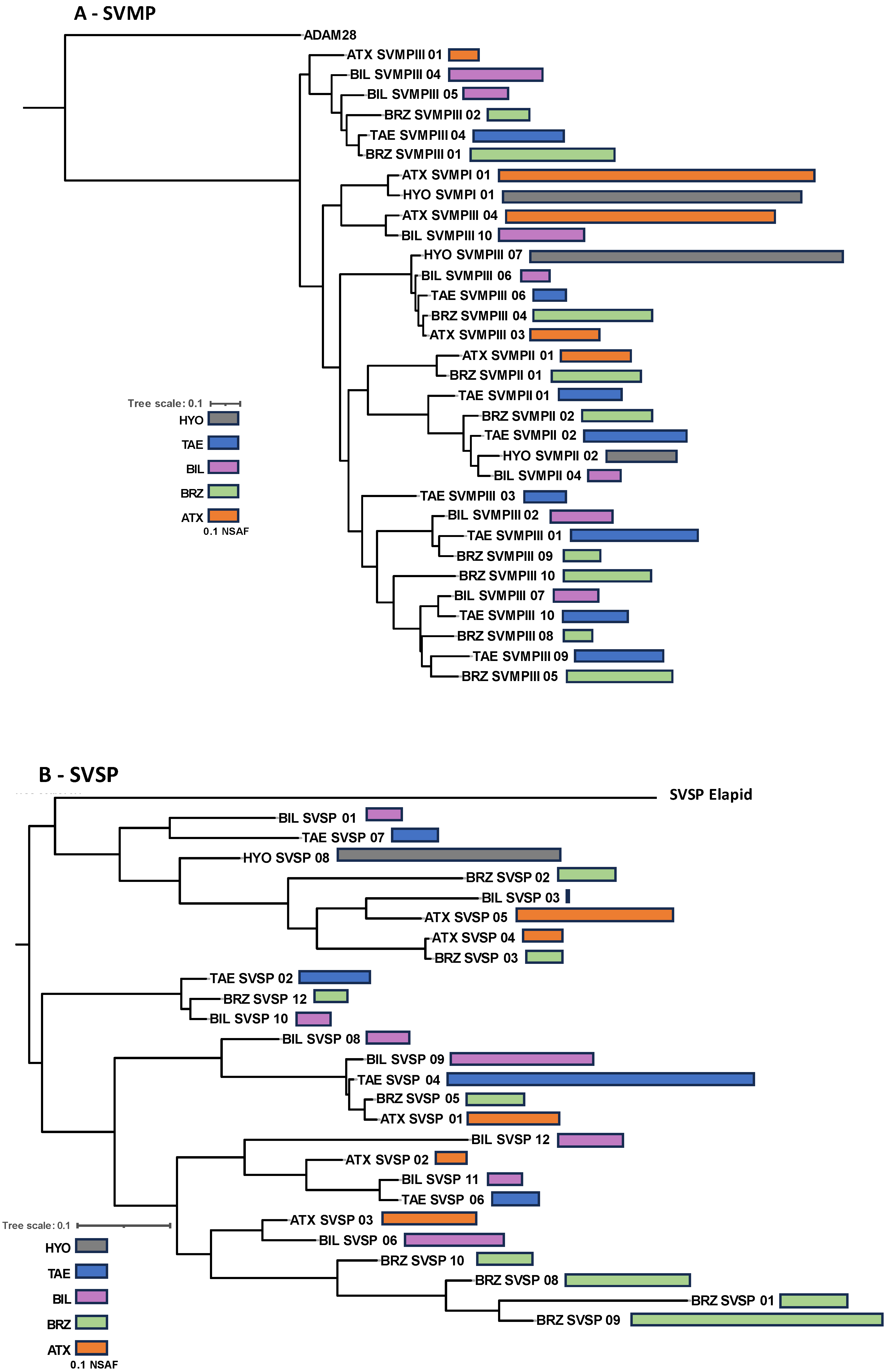
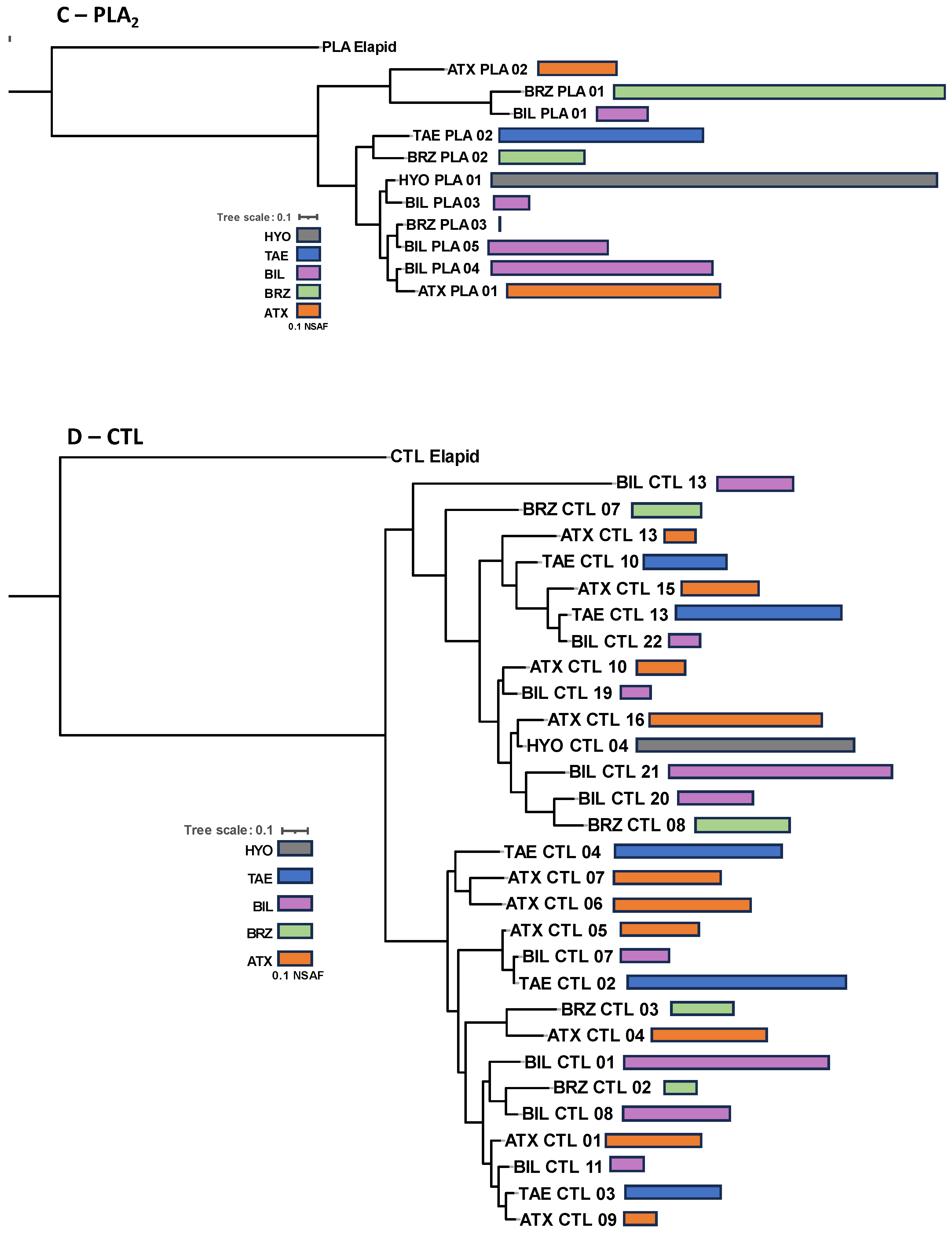
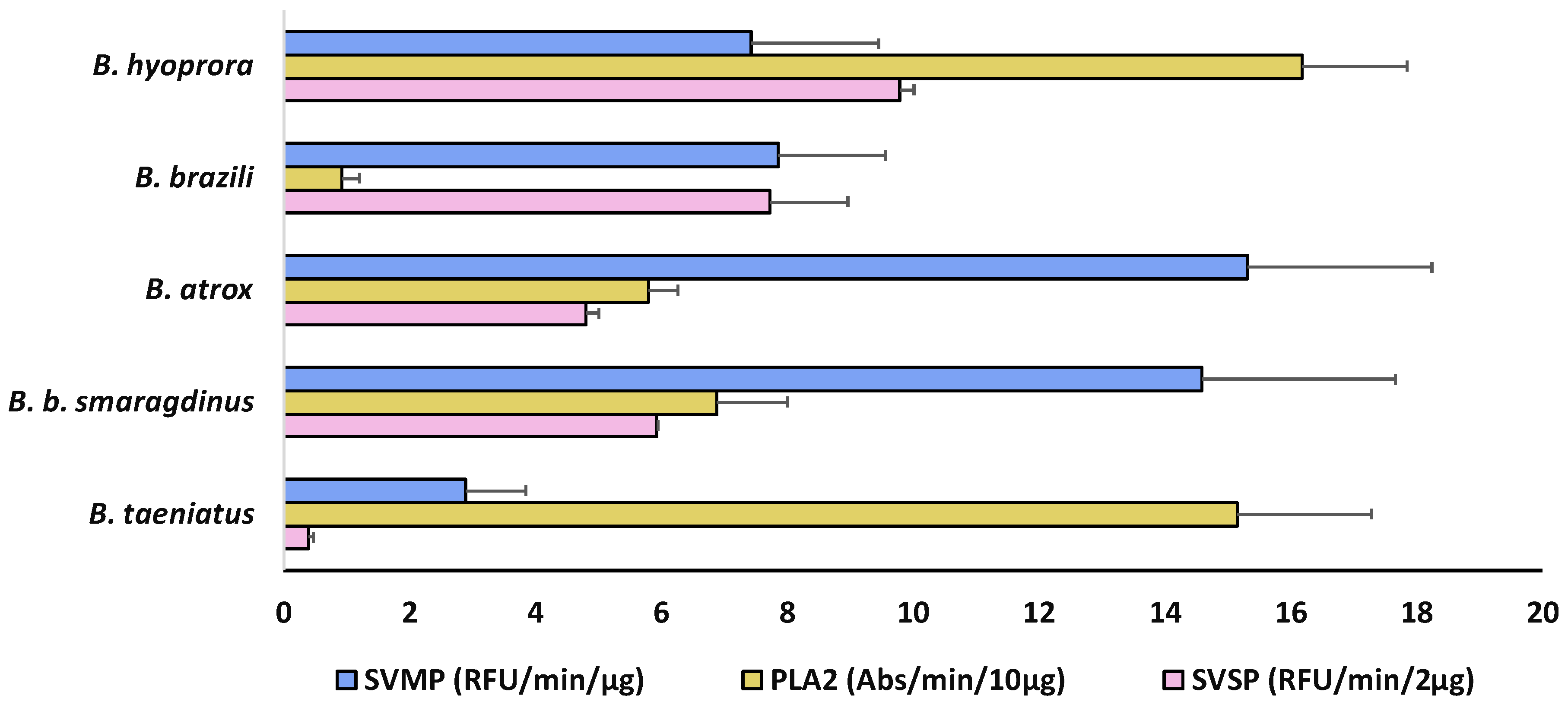
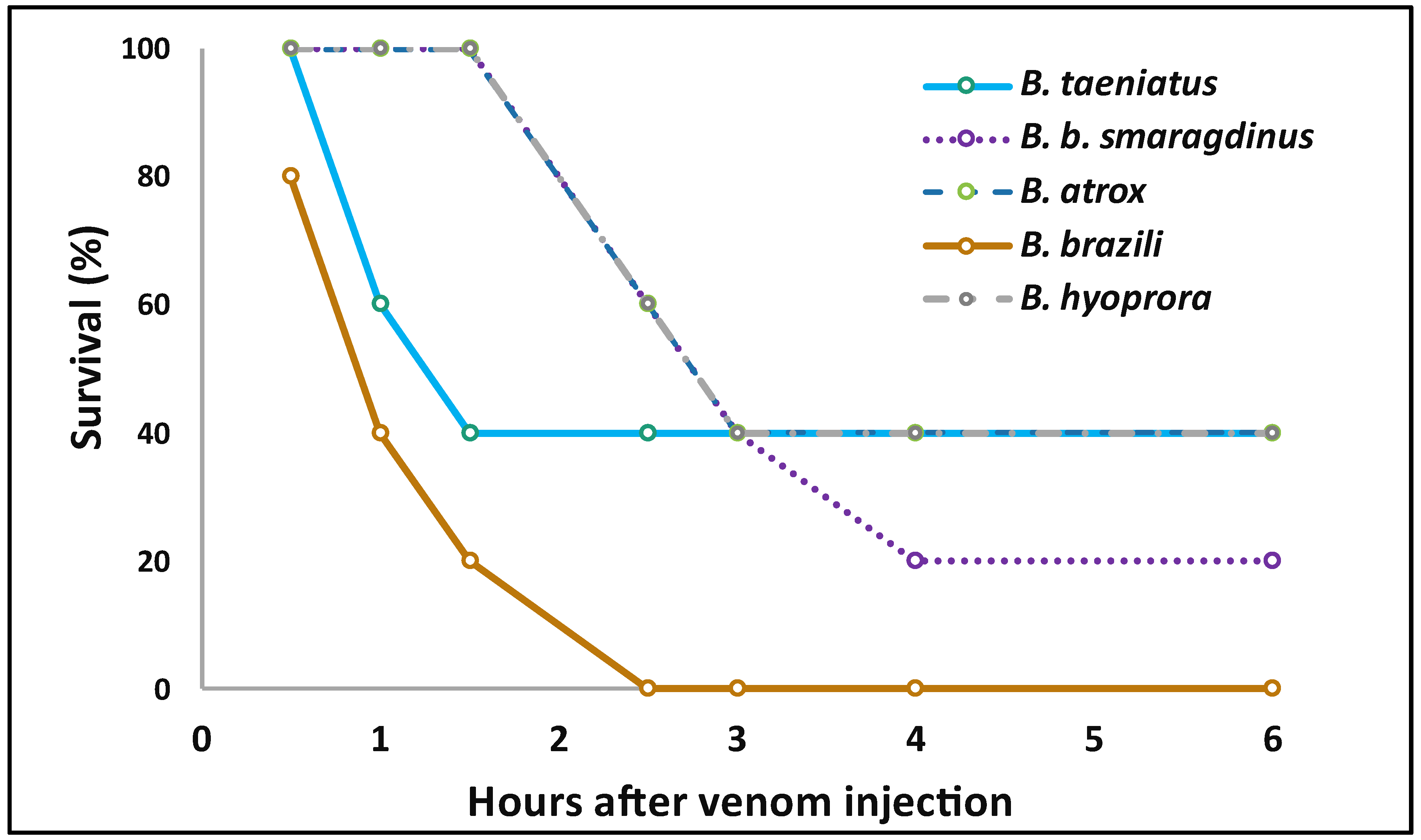
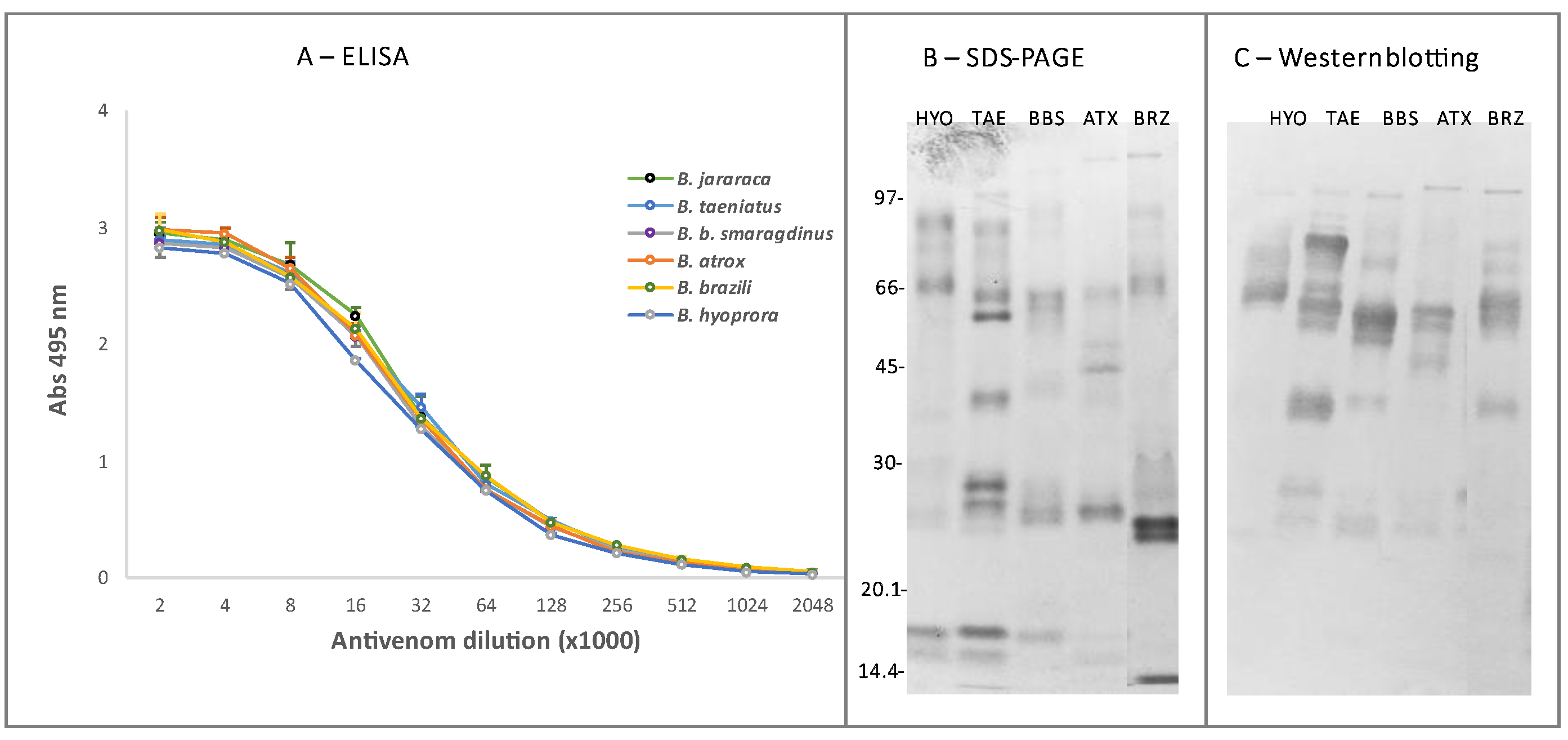
2. Results and Discussion
3. Conclusions
4. Materials and Methods
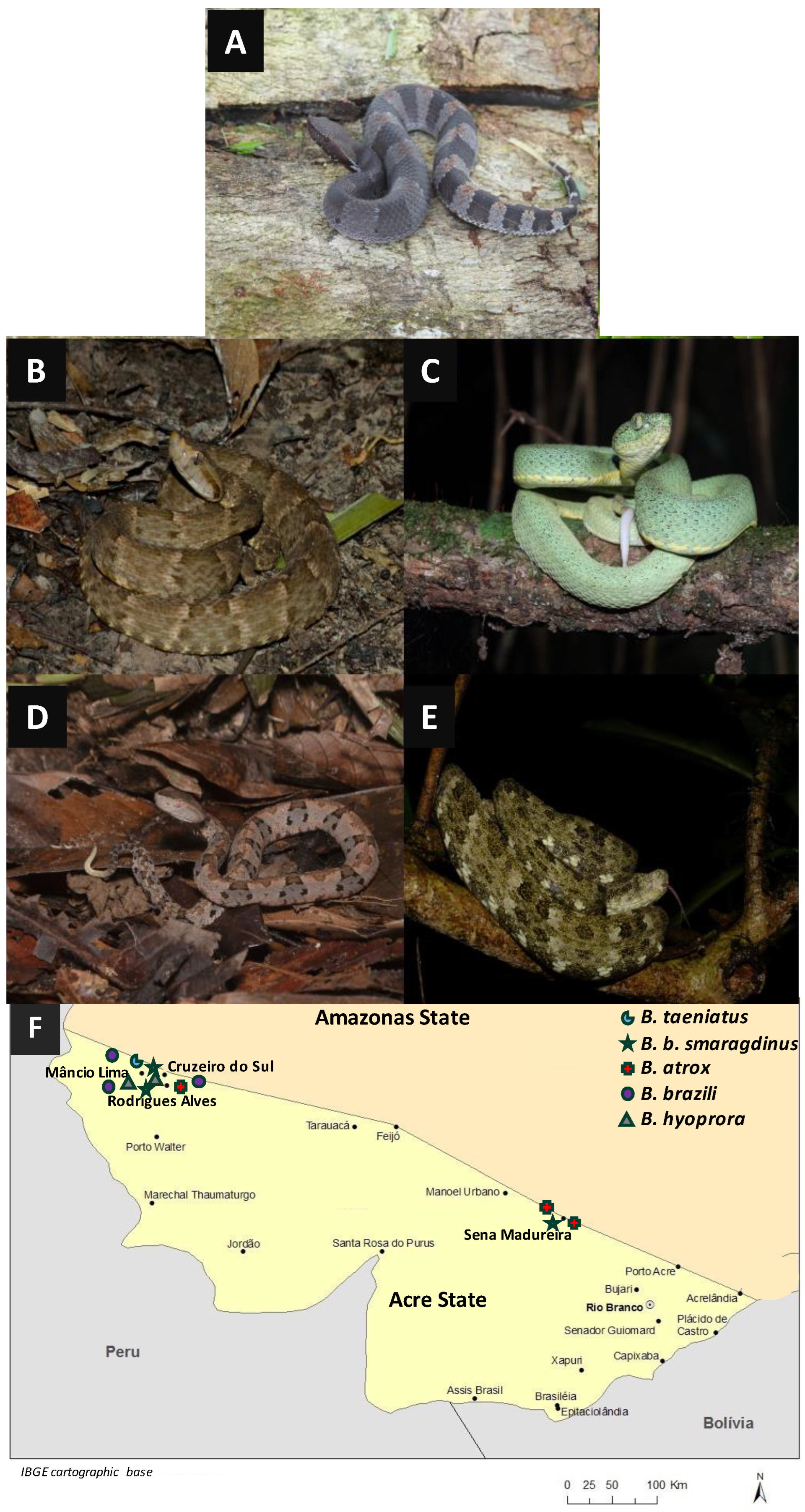
4.1. Snakes and Venoms
4.2. Venom Fractionation by Reverse Phase Chromatography
4.3. Transcriptome
4.3.1. RNA Extraction and Analysis; cDNA Library Construction and Sequencing
4.3.2. Transcriptome Assembly and Annotation
4.4. Proteomic Analysis of Venoms
4.4.1. Analysis via In-Solution Trypsin Digestion and LC−MS/MS
4.4.2. Identification and Relative Quantification
4.5. Sequence Alignments and Gene Tree Analyses
4.6. Enzymatic Assays
4.7. Reactivity with Antivenoms
4.8. Lethality Test
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guedes, T.; Entiauspe, O.; Costa, H. Lista de répteis do Brasil: Atualização de 2022. Herpetol. Bras. 2023, 12, 56–161. [Google Scholar]
- Bernarde, P.; Turci, L.; Machado, R. Serpentes do Alto Juruá, Acre—Amazônia Brasileira; EDUFAC, Ed.; EDUFAC: Rio Branco, AC, Brazil, 2017. [Google Scholar]
- Silveira, M.; Torezan, J.; Daly, D. Vegetação e Diversidade Arbórea da Região do Alto Juruá. In Enciclopédia da Floresta: O Alto Juruá: Práticas e Conhecimento das Populações; Cunha, M., Almeida, M., Eds.; Companhia das Letras: São Paulo, Brazil, 2002; p. 735. [Google Scholar]
- Ab’Saber, A.N. The Paleoclimate and Paleoecology of Brazilian Amazonia. In Biological Diversification in the Tropics; Prance, G.T., Ed.; Columbia University Press: New York, NY, USA, 1982; pp. 41–59. [Google Scholar]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of the Western Hemisphere; Comstock Pub. Associates: Ithaca, NY, USA, 2004. [Google Scholar]
- Daltry, J.C.; Wüster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef]
- Cipriani, V.; Debono, J.; Goldenberg, J.; Jackson, T.N.W.; Arbuckle, K.; Dobson, J.; Koludarov, I.; Li, B.; Hay, C.; Dunstan, N.; et al. Correlation between ontogenetic dietary shifts and venom variation in Australian brown snakes (Pseudonaja). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 197, 53–60. [Google Scholar] [CrossRef]
- Sousa, L.F.; Portes-Junior, J.A.; Nicolau, C.A.; Bernardoni, J.L.; Nishiyama, M.Y., Jr.; Amazonas, D.R.; Freitas-de-Sousa, L.A.; Mourao, R.H.V.; Chalkidis, H.M.; Valente, R.H.; et al. Functional proteomic analyses of Bothrops atrox venom reveals phenotypes associated with habitat variation in the Amazon. J. Proteom. 2017, 159, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.J.; Wray, K.P.; Seavy, M.; McGivern, J.J.; Sanader, D.; Rokyta, D.R. Phenotypic integration in the feeding system of the eastern diamondback rattlesnake (Crotalus adamanteus). Mol. Ecol. 2015, 24, 3405–3420. [Google Scholar] [CrossRef]
- Schonour, R.B.; Huff, E.M.; Holding, M.L.; Claunch, N.M.; Ellsworth, S.A.; Hogan, M.P.; Wray, K.; McGivern, J.; Margres, M.J.; Colston, T.J.; et al. Gradual and Discrete Ontogenetic Shifts in Rattlesnake Venom Composition and Assessment of Hormonal and Ecological Correlates. Toxins 2020, 12, 659. [Google Scholar] [CrossRef]
- Hatakeyama, D.M.; Tasima, L.J.; Bravo-Tobar, C.A.; Serino-Silva, C.; Tashima, A.K.; Rodrigues, C.F.B.; Aguiar, W.D.S.; Galizio, N.D.C.; de Lima, E.O.V.; Kavazoi, V.K.; et al. Venom complexity of Bothrops atrox (common lancehead) siblings. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200018. [Google Scholar] [CrossRef]
- Alape-Girón, A.; Sanz, L.; Escolano, J.; Flores-Díaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancehead pitviper Bothrops asper: Geographic, individual, and ontogenetic variations. J. Proteome Res. 2008, 7, 3556–3571. [Google Scholar] [CrossRef]
- Durban, J.; Pérez, A.; Sanz, L.; Gómez, A.; Bonilla, F.; Rodríguez, S.; Chacón, D.; Sasa, M.; Angulo, Y.; Gutiérrez, J.M.; et al. Integrated “omics” profiling indicates that miRNAs are modulators of the ontogenetic venom composition shift in the Central American rattlesnake, Crotalus simus simus. BMC Genom. 2013, 14, 234. [Google Scholar] [CrossRef]
- López-Lozano, J.L.; de Sousa, M.V.; Ricart, C.A.; Chávez-Olortegui, C.; Flores Sanchez, E.; Muniz, E.G.; Bührnheim, P.F.; Morhy, L. Ontogenetic variation of metalloproteinases and plasma coagulant activity in venoms of wild Bothrops atrox specimens from Amazonian rain forest. Toxicon 2002, 40, 997–1006. [Google Scholar] [CrossRef]
- Salazar, A.M.; Rodriguez-Acosta, A.; Girón, M.E.; Aguilar, I.; Guerrero, B. A comparative analysis of the clotting and fibrinolytic activities of the snake venom (Bothrops atrox) from different geographical areas in Venezuela. Thromb. Res. 2007, 120, 95–104. [Google Scholar] [CrossRef]
- Patiño, R.S.P.; Salazar-Valenzuela, D.; Medina-Villamizar, E.; Mendes, B.; Proaño-Bolaños, C.; da Silva, S.L.; Almeida, J.R. Bothrops atrox from Ecuadorian Amazon: Initial analyses of venoms from individuals. Toxicon 2021, 193, 63–72. [Google Scholar] [CrossRef]
- Dagda, R.K.; Gasanov, S.; De La Oiii, Y.; Rael, E.D.; Lieb, C.S. Genetic Basis for Variation of Metalloproteinase-Associated Biochemical Activity in Venom of the Mojave Rattlesnake (Crotalus scutulatus scutulatus). Biochem. Res. Int. 2013, 2013, 251474. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Neto, A.; Caldeira, C.A.; Souza, G.H.; Zaqueo, K.D.; Kayano, A.M.; Silva, R.S.; Zuliani, J.P.; Soares, A.M.; Stábeli, R.G.; Calderon, L.A. ESI-MS/MS identification of a bradykinin-potentiating peptide from Amazon Bothrops atrox snake venom using a hybrid Qq-oaTOF mass spectrometer. Toxins 2013, 5, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Guércio, R.A.; Shevchenko, A.; López-Lozano, J.L.; Paba, J.; Sousa, M.V.; Ricart, C.A. Ontogenetic variations in the venom proteome of the Amazonian snake Bothrops atrox. Proteome Sci. 2006, 4, 11. [Google Scholar] [CrossRef]
- Kohlhoff, M.; Borges, M.H.; Yarleque, A.; Cabezas, C.; Richardson, M.; Sanchez, E.F. Exploring the proteomes of the venoms of the Peruvian pit vipers Bothrops atrox, B. barnetti and B. pictus. J. Proteom. 2012, 75, 2181–2195. [Google Scholar] [CrossRef]
- Núñez, V.; Cid, P.; Sanz, L.; De La Torre, P.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of Bothrops atrox venoms from Colombia and the Amazon regions of Brazil, Perú and Ecuador suggest the occurrence of geographic variation of venom phenotype by a trend towards paedomorphism. J. Proteom. 2009, 73, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Pérez, A.; Quesada-Bernat, S.; Diniz-Sousa, R.; Calderón, L.A.; Soares, A.M.; Calvete, J.J.; Caldeira, C.A.S. Venomics and antivenomics of the poorly studied Brazil’s lancehead, Bothrops brazili (Hoge, 1954), from the Brazilian State of Pará. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190103. [Google Scholar] [CrossRef]
- Sanz, L.; Quesada-Bernat, S.; Pérez, A.; De Morais-Zani, K.; Sant’Anna, S.S.; Hatakeyama, D.M.; Tasima, L.J.; De Souza, M.B.; Kayano, A.M.; Zavaleta, A.; et al. Danger in the Canopy. Comparative Proteomics and Bioactivities of the Venoms of the South American Palm Pit Viper. J. Proteome Res. 2020, 19, 3518–3532. [Google Scholar] [CrossRef]
- Pereañez, J.A.; Preciado, L.M.; Fernández, J.; Camacho, E.; Lomonte, B.; Castro, F.; Cañas, C.A.; Galvis, C.; Castaño, S. Snake venomics, experimental toxic activities and clinical characteristics of human envenomation by Bothrocophias myersi (Serpentes: Viperidae) from Colombia. J. Proteom. 2020, 220, 103758. [Google Scholar] [CrossRef]
- Salazar-Valenzuela, D.; Mora-Obando, D.; Fernández, M.L.; Loaiza-Lange, A.; Gibbs, H.L.; Lomonte, B. Proteomic and toxicological profiling of the venom of Bothrocophias campbelli, a pitviper species from Ecuador and Colombia. Toxicon 2014, 90, 15–25. [Google Scholar] [CrossRef]
- Lomonte, B.; Díaz, C.; Chaves, F.; Fernández, J.; Ruiz, M.; Salas, M.; Zavaleta, A.; Calvete, J.J.; Sasa, M. Comparative characterization of Viperidae snake venoms from Perú reveals two compositional patterns of phospholipase A. Toxicon X 2020, 7, 100044. [Google Scholar] [CrossRef] [PubMed]
- Angulo, Y.; Escolano, J.; Lomonte, B.; Gutiérrez, J.M.; Sanz, L.; Calvete, J.J. Snake venomics of Central American pitvipers: Clues for rationalizing the distinct envenomation profiles of Atropoides nummifer and Atropoides picadoi. J. Proteome Res. 2008, 7, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Borges, A.; Segura, A.; Flores-Díaz, M.; Alape-Girón, A.; Gutiérrez, J.M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sánchez, E.; et al. Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management. J. Proteom. 2009, 72, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Escolano, J.; Fernández, J.; Sanz, L.; Angulo, Y.; Gutiérrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of the arboreal neotropical pitvipers Bothriechis lateralis and Bothriechis schlegelii. J. Proteome Res. 2008, 7, 2445–2457. [Google Scholar] [CrossRef] [PubMed]
- Amazonas, D.R.; Portes-Junior, J.A.; Nishiyama-Jr, M.Y.; Nicolau, C.A.; Chalkidis, H.M.; Mourão, R.H.V.; Grazziotin, F.G.; Rokyta, D.R.; Gibbs, H.L.; Valente, R.H.; et al. Molecular mechanisms underlying intraspecific variation in snake venom. J. Proteom. 2018, 181, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Durban, J.; Juárez, P.; Angulo, Y.; Lomonte, B.; Flores-Diaz, M.; Alape-Girón, A.; Sasa, M.; Sanz, L.; Gutiérrez, J.M.; Dopazo, J.; et al. Profiling the venom gland transcriptomes of Costa Rican snakes by 454 pyrosequencing. BMC Genom. 2011, 12, 259. [Google Scholar] [CrossRef]
- Margres, M.J.; McGivern, J.J.; Wray, K.P.; Seavy, M.; Calvin, K.; Rokyta, D.R. Linking the transcriptome and proteome to characterize the venom of the eastern diamondback rattlesnake (Crotalus adamanteus). J. Proteom. 2014, 96, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Rokyta, D.R.; Lemmon, A.R.; Margres, M.J.; Aronow, K. The venom-gland transcriptome of the eastern diamondback rattlesnake (Crotalus adamanteus). BMC Genom. 2012, 13, 312. [Google Scholar] [CrossRef]
- Hofmann, E.P.; Rautsaw, R.M.; Strickland, J.L.; Holding, M.L.; Hogan, M.P.; Mason, A.J.; Rokyta, D.R.; Parkinson, C.L. Comparative venom-gland transcriptomics and venom proteomics of four Sidewinder Rattlesnake (Crotalus cerastes) lineages reveal little differential expression despite individual variation. Sci. Rep. 2018, 8, 15534. [Google Scholar] [CrossRef]
- Calvete, J.J.; Domont, G.B. Omic technologies to fight the neglect. J. Proteom. 2011, 74, 1483–1484. [Google Scholar] [CrossRef]
- Neiva, M.; Arraes, F.B.; de Souza, J.V.; Rádis-Baptista, G.; Prieto da Silva, A.R.; Walter, M.E.; Brigido, M.e.M.; Yamane, T.; López-Lozano, J.L.; Astolfi-Filho, S. Transcriptome analysis of the Amazonian viper Bothrops atrox venom gland using expressed sequence tags (ESTs). Toxicon 2009, 53, 427–436. [Google Scholar] [CrossRef]
- Mason, A.J.; Margres, M.J.; Strickland, J.L.; Rokyta, D.R.; Sasa, M.; Parkinson, C.L. Trait differentiation and modular toxin expression in palm-pitvipers. BMC Genom. 2020, 21, 147. [Google Scholar] [CrossRef]
- Barros, S.F.; Friedlanskaia, I.; Petricevich, V.L.; Kipnis, T.L. Local inflammation, lethality and cytokine release in mice injected with Bothrops atrox venom. Mediat. Inflamm. 1998, 7, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, R.A.; Remold, H.; Kipnis, T.L. Purification and variability in thrombin-like activity of Bothrops atrox venom from different geographic regions. Toxicon 1998, 36, 257–267. [Google Scholar] [CrossRef]
- Costal-Oliveira, F.; Stransky, S.; Guerra-Duarte, C.; Naves de Souza, D.L.; Vivas-Ruiz, D.E.; Yarlequé, A.; Sanchez, E.F.; Chávez-Olórtegui, C.; Braga, V.M.M. L-amino acid oxidase from Bothrops atrox snake venom triggers autophagy, apoptosis and necrosis in normal human keratinocytes. Sci. Rep. 2019, 9, 781. [Google Scholar] [CrossRef]
- Furtado, J.L.; Oliveira, G.A.; Pontes, A.S.; Setúbal, S.a.S.; Xavier, C.V.; Lacouth-Silva, F.; Lima, B.F.; Zaqueo, K.D.; Kayano, A.M.; Calderon, L.A.; et al. Activation of J77A.1 macrophages by three phospholipases A2 isolated from Bothrops atrox snake venom. Biomed. Res. Int. 2014, 2014, 683123. [Google Scholar] [CrossRef] [PubMed]
- Moretto Del-Rei, T.H.; Sousa, L.F.; Rocha, M.M.T.; Freitas-de-Sousa, L.A.; Travaglia-Cardoso, S.R.; Grego, K.; Sant’Anna, S.S.; Chalkidis, H.M.; Moura-da-Silva, A.M. Functional variability of Bothrops atrox venoms from three distinct areas across the Brazilian Amazon and consequences for human envenomings. Toxicon 2019, 164, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Silva, D.; Zelanis, A.; Travaglia-Cardoso, S.R.; Nishiyama, M.Y.; Serrano, S.M.T. Venom Profiling of the Insular Species. J. Proteome Res. 2021, 20, 1341–1358. [Google Scholar] [CrossRef]
- Freitas-de-Sousa, L.A.; Colombini, M.; Lopes-Ferreira, M.; Serrano, S.M.T.; Moura-da-Silva, A.M. Insights into the Mechanisms Involved in Strong Hemorrhage and Dermonecrosis Induced by Atroxlysin-Ia, a PI-Class Snake Venom Metalloproteinase. Toxins 2017, 9, 1341–1358. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.F.; Schneider, F.S.; Yarleque, A.; Borges, M.H.; Richardson, M.; Figueiredo, S.G.; Evangelista, K.S.; Eble, J.A. The novel metalloproteinase atroxlysin-I from Peruvian Bothrops atrox (Jergón) snake venom acts both on blood vessel ECM and platelets. Arch. Biochem. Biophys. 2010, 496, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Alencar, L.R.V.; Quental, T.B.; Grazziotin, F.G.; Alfaro, M.L.; Martins, M.; Venzon, M.; Zaher, H. Diversification in vipers: Phylogenetic relationships, time of divergence and shifts in speciation rates. Mol. Phylogenetics Evol. 2016, 105, 50–62. [Google Scholar] [CrossRef]
- Freitas-de-Sousa, L.A.; Amazonas, D.R.; Sousa, L.F.; Sant’Anna, S.S.; Nishiyama, M.Y., Jr.; Serrano, S.M.T.; Junqueira-de-Azevedo, I.L.M.; Chalkidis, H.M.; Moura-da-Silva, A.M.; Mourao, R.H.V. Comparison of venoms from wild and long-term captive Bothrops atrox snakes and characterization of Batroxrhagin, the predominant class PIII metalloproteinase from the venom of this species. Biochimie 2015, 118, 60–70. [Google Scholar] [CrossRef]
- Lomonte, B.; Rangel, J. Snake venom Lys49 myotoxins: From phospholipases A2 to non-enzymatic membrane disruptors. Toxicon 2012, 60, 520–530. [Google Scholar] [CrossRef]
- Bourke, L.A.; Zdenek, C.N.; Tanaka-Azevedo, A.M.; Silveira, G.P.M.; Sant’Anna, S.S.; Grego, K.F.; Rodrigues, C.F.B.; Fry, B.G. Clinical and Evolutionary Implications of Dynamic Coagulotoxicity Divergences in. Toxins 2022, 14, 297. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.F.; Cardoso, S.T.; Soares, O.E.; Pereira, A.P.; Fernandes, D.S.; Tambourgi, D.V.; Sant’Anna, O.A. Antigenic cross-reactivity and immunogenicity of Bothrops venoms from snakes of the Amazon region. Toxicon 2010, 55, 881–887. [Google Scholar] [CrossRef]
- Martins, M.; Marques, O.A.V.; Sazima, I.; Schuett, G.; Höggren, M.; Green, H.W. Ecological and phylogenetic correlates of feeding habits in Neotropicalpitvipers of the genus Bothrops. Biol. Vipers 2002, 307, 328. [Google Scholar]
- Segura, A.; Castillo, M.C.; Núñez, V.; Yarlequé, A.; Gonçalves, L.R.; Villalta, M.; Bonilla, C.; Herrera, M.; Vargas, M.; Fernández, M.; et al. Preclinical assessment of the neutralizing capacity of antivenoms produced in six Latin American countries against medically-relevant Bothrops snake venoms. Toxicon 2010, 56, 980–989. [Google Scholar] [CrossRef]
- Mackessy, S.P. Evolutionary trends in venom composition in the western rattlesnakes (Crotalus viridis sensu lato): Toxicity vs. tenderizers. Toxicon 2010, 55, 1463–1474. [Google Scholar] [CrossRef]
- Kini, R.M. Serine proteases affecting blood coagulation and fibrinolysis from snake venoms. Pathophysiol. Haemost. Thromb. 2005, 34, 200–204. [Google Scholar] [CrossRef]
- Chacón, F.; Oviedo, A.; Escalante, T.; Solano, G.; Rucavado, A.; Gutiérrez, J.M. The lethality test used for estimating the potency of antivenoms against Bothrops asper snake venom: Pathophysiological mechanisms, prophylactic analgesia, and a surrogate in vitro assay. Toxicon 2015, 93, 41–50. [Google Scholar] [CrossRef]
- Cañas, C.A. Brainstem ischemic stroke after to Bothrops atrox snakebite. Toxicon 2016, 120, 124–127. [Google Scholar] [CrossRef]
- Kallel, H.; Mayence, C.; Houcke, S.; Mathien, C.; Mehdaoui, H.; Gutiérrez, J.M.; Megarbane, B.; Hommel, D.; Resiere, D. Severe snakebite envenomation in French Guiana: When antivenom is not available. Toxicon 2018, 146, 87–90. [Google Scholar] [CrossRef] [PubMed]
- França, F.O.; Barbaro, K.C.; Fan, H.W.; Cardoso, J.L.; Sano-Martins, I.S.; Tomy, S.C.; Lopes, M.H.; Warrell, D.A.; Theakston, R.D.; Butantan Institute Antivenom Study Group. Envenoming by Bothrops jararaca in Brazil: Association between venom antigenaemia and severity at admission to hospital. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Torrez, P.Q.; Duarte, M.R.; França, F.O.; Figueiredo, L.; Abati, P.; Campos, L.R.; Pardal, P.P.; Quiroga, M.; Mascheretti, M.; Boulos, M. First report of an accident with the speckled forest pit viper (Bothriopsis taeniata) in Brazil. Rev. Soc. Bras. Med. Trop. 2009, 42, 342–344. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Porto, B.N.; Telli, C.A.; Dutra, T.P.; Alves, L.S.; Bozza, M.T.; Fin, C.A.; Thiesen, F.V.; Renner, M.F. Biochemical and biological characterization of the venoms of Bothriopsis bilineata and Bothriopsis taeniata (Serpentes: Viperidae). Toxicon 2007, 50, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Arlinghaus, F.T.; Eble, J.A. C-type lectin-like proteins from snake venoms. Toxicon 2012, 60, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Zingali, R.B.; Ferreira, M.S.; Assafim, M.; Frattani, F.S.; Monteiro, R.Q. Bothrojaracin, a Bothrops jararaca snake venom-derived (pro)thrombin inhibitor, as an anti-thrombotic molecule. Pathophysiol. Haemost. Thromb. 2005, 34, 160–163. [Google Scholar] [CrossRef]
- Gartner, T.K.; Stocker, K.; Williams, D.C. Thrombolectin: A lectin isolated from Bothrops atrox venom. FEBS Lett. 1980, 117, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Tani, A.; Ogawa, T.; Nose, T.; Nikandrov, N.N.; Deshimaru, M.; Chijiwa, T.; Chang, C.C.; Fukumaki, Y.; Ohno, M. Characterization, primary structure and molecular evolution of anticoagulant protein from Agkistrodon actus venom. Toxicon 2002, 40, 803–813. [Google Scholar] [CrossRef]
- Butantan, I. Soro Antibotrópico Pentavalente. Available online: https://butantan.gov.br/assets/arquivos/soros-e-vacinas/soros/Soro%20antibotrópico-crotálico.pdf (accessed on 25 November 2023).
- Muniz, E.G.; Sano-Martins, I.S.; Saraiva, M.D.G.G.; Monteiro, W.M.; Magno, E.S.; Oliveira, S.S. Biological characterization of the Bothrops brazili snake venom and its neutralization by Brazilian Bothrops antivenom produced by the Butantan Institute. Toxicon 2023, 223, 107010. [Google Scholar] [CrossRef]
- Muniz, E.G.; Maria, W.S.; Estevão-Costa, M.I.; Buhrnheim, P.; Chávez-Olórtegui, C. Neutralizing potency of horse antibothropic Brazilian antivenom against Bothrops snake venoms from the Amazonian rain forest. Toxicon 2000, 38, 1859–1863. [Google Scholar] [CrossRef]
- Calvete, J.J.; Juárez, P.; Sanz, L. Snake venomics. Strategy and applications. J. Mass Spectrom. 2007, 42, 1405–1414. [Google Scholar] [CrossRef]
- Bernardoni, J.L.; Sousa, L.F.; Wermelinger, L.S.; Lopes, A.S.; Prezoto, B.C.; Serrano, S.M.T.; Zingali, R.B.; Moura-da-Silva, A.M. Functional Variability of Snake Venom Metalloproteinases: Adaptive Advantages in Targeting Different Prey and Implications for Human Envenomation. PLoS ONE 2014, 9, e109651. [Google Scholar] [CrossRef]
- Sousa, L.F.; Nicolau, C.A.; Peixoto, P.S.; Bernardoni, J.L.; Oliveira, S.S.; Portes-Junior, J.A.; Mourao, R.H.V.; Lima-dos-Santos, I.; Sano-Martins, I.S.; Chalkidis, H.M.; et al. Comparison of Phylogeny, Venom Composition and Neutralization by Antivenom in Diverse Species of Bothrops Complex. PLoS Negl. Trop. Dis. 2013, 7, e2442. [Google Scholar] [CrossRef]
- Holding, M.L.; Strickland, J.L.; Rautsaw, R.M.; Hofmann, E.P.; Mason, A.J.; Hogan, M.P.; Nystrom, G.S.; Ellsworth, S.A.; Colston, T.J.; Borja, M.; et al. Phylogenetically diverse diets favor more complex venoms in North American pitvipers. Proc. Natl. Acad. Sci. USA 2021, 118, e2015579118. [Google Scholar] [CrossRef]
- Barua, A.; Mikheyev, A.S. Many Options, Few Solutions: Over 60 My Snakes Converged on a Few Optimal Venom Formulations. Mol. Biol. Evol. 2019, 36, 1964–1974. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Nachtigall, P.G.; Freitas-de-Sousa, L.A.; Mason, A.J.; Moura-da-Silva, A.M.; Grazziotin, F.G.; Junqueira-de-Azevedo, I.L.M. Differences in PLA. Toxins 2022, 14, 237. [Google Scholar] [CrossRef]
- Krueger, F. Trim Galore!: A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files. 2015. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 10 November 2022).
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Chang, Z.; Li, G.; Liu, J.; Zhang, Y.; Ashby, C.; Liu, D.; Cramer, C.L.; Huang, X. Bridger: A new framework for de novo transcriptome assembly using RNA-seq data. Genome Biol. 2015, 16, 30. [Google Scholar] [CrossRef]
- Bushmanova, E.; Antipov, D.; Lapidus, A.; Prjibelski, A.D. rnaSPAdes: A de novo transcriptome assembler and its application to RNA-Seq data. Gigascience 2019, 8, giz100. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Farias, I.B.; Morais-Zani, K.; Serino-Silva, C.; Sant’Anna, S.S.; Rocha, M.M.T.D.; Grego, K.F.; Andrade-Silva, D.; Serrano, S.M.T.; Tanaka-Azevedo, A.M. Functional and proteomic comparison of Bothrops jararaca venom from captive specimens and the Brazilian Bothropic Reference Venom. J. Proteom. 2018, 174, 36–46. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Mann, M. Combination of FASP and StageTip-based fractionation allows in-depth analysis of the hippocampal membrane proteome. J. Proteome Res. 2009, 8, 5674–5678. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Dávila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Junier, T.; Zdobnov, E.M. The Newick utilities: High-throughput phylogenetic tree processing in the UNIX shell. Bioinformatics 2010, 26, 1669–1670. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Class * | Bothrocophias hioprora | Bothrops atrox | Bothrops bilineatus smaragdinus | Bothrops brazili | Bothrops taeniatus |
|---|---|---|---|---|---|
| SVMP | 14 | 6 | 17 | 14 | 12 |
| CTL | 4 | 16 | 22 | 9 | 15 |
| SVSP | 12 | 6 | 12 | 13 | 7 |
| PLA2 | 2 | 2 | 5 | 3 | 4 |
| VEGF | 3 | 2 | 3 | 3 | 2 |
| LAO | 0 | 2 | 1 | 1 | 0 |
| HYAL | 1 | 1 | 1 | 1 | 0 |
| CRISP | 1 | 1 | 1 | 1 | 1 |
| NGF | 1 | 1 | 1 | 1 | 0 |
| NUC | 1 | 0 | 1 | 1 | 2 |
| PDE | 2 | 1 | 2 | 2 | 1 |
| PLB | 1 | 1 | 1 | 1 | 1 |
| BPP | 1 | 0 | 1 | 1 | 0 |
| DIPEP | 1 | 0 | 1 | 1 | 0 |
| CYS | 1 | 0 | 1 | 2 | 0 |
| iPLA | 0 | 0 | 1 | 1 | 0 |
| LIPA | 1 | 0 | 1 | 1 | 1 |
| WAP | 1 | 0 | 0 | 2 | 1 |
| KUN | 1 | 0 | 0 | 0 | 0 |
| Total | 48 | 39 | 72 | 58 | 47 |
| Code | Genus | Sex | Age | SVL (mm) | City | Locality |
|---|---|---|---|---|---|---|
| SB0779 | Bothrocophias hyoprora | F | adult | 370 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB0780 | Bothrocophias hyoprpra | M | juvenile | 252 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB0597 | Bothrops atrox | M | adult | 1020 | Sena Madureira—Acre | RESEX Cazumbá-Iracema |
| SB0600 | Bothrops atrox | F | adult | 1090 | Sena Madureira—Acre | RESEX Cazumbá-Iracema |
| SB0796 | Bothrops atrox | M | adult | 1035 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB0582 | Bothrops b. smaragdinus | M | adult | 532 | Sena Madureira—Acre | RESEX Cazumbá-Iracema |
| SB0782 | Bothrops b. smaragdinus | F | adult | 675 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB1942 | Bothrops b. smaragdinus | M | adult | 534 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB1455 | Bothrops brazili | M | adult | 795 | Mâncio Lima—Acre | Serra do Divisor |
| SB1948 | Bothrops brazili | F | adult | 685 | Mâncio Lima—Acre | PARNA Serra do Divisor, Formosa trail |
| SB1953 | Bothrops brazili | F | juvenile | 341 | Guajará—Amazonas | ND |
| SB1580 | Bothrops taeniatus | F | adult | 517 | Mâncio Lima—Acre | PARNA Serra do Divisor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas-de-Sousa, L.A.; Colombini, M.; Souza, V.C.; Silva, J.P.C.; Mota-da-Silva, A.; Almeida, M.R.N.; Machado, R.A.; Fonseca, W.L.; Sartim, M.A.; Sachett, J.; et al. Venom Composition of Neglected Bothropoid Snakes from the Amazon Rainforest: Ecological and Toxinological Implications. Toxins 2024, 16, 83. https://doi.org/10.3390/toxins16020083
Freitas-de-Sousa LA, Colombini M, Souza VC, Silva JPC, Mota-da-Silva A, Almeida MRN, Machado RA, Fonseca WL, Sartim MA, Sachett J, et al. Venom Composition of Neglected Bothropoid Snakes from the Amazon Rainforest: Ecological and Toxinological Implications. Toxins. 2024; 16(2):83. https://doi.org/10.3390/toxins16020083
Chicago/Turabian StyleFreitas-de-Sousa, Luciana A., Mônica Colombini, Vinicius C. Souza, Joanderson P. C. Silva, Ageane Mota-da-Silva, Marllus R. N. Almeida, Reginaldo A. Machado, Wirven L. Fonseca, Marco A. Sartim, Jacqueline Sachett, and et al. 2024. "Venom Composition of Neglected Bothropoid Snakes from the Amazon Rainforest: Ecological and Toxinological Implications" Toxins 16, no. 2: 83. https://doi.org/10.3390/toxins16020083
APA StyleFreitas-de-Sousa, L. A., Colombini, M., Souza, V. C., Silva, J. P. C., Mota-da-Silva, A., Almeida, M. R. N., Machado, R. A., Fonseca, W. L., Sartim, M. A., Sachett, J., Serrano, S. M. T., Junqueira-de-Azevedo, I. L. M., Grazziotin, F. G., Monteiro, W. M., Bernarde, P. S., & Moura-da-Silva, A. M. (2024). Venom Composition of Neglected Bothropoid Snakes from the Amazon Rainforest: Ecological and Toxinological Implications. Toxins, 16(2), 83. https://doi.org/10.3390/toxins16020083