Functional Comparison of Three Chitinases from Symbiotic Bacteria of Entomopathogenic Nematodes


Abstract
1. Introduction
2. Results
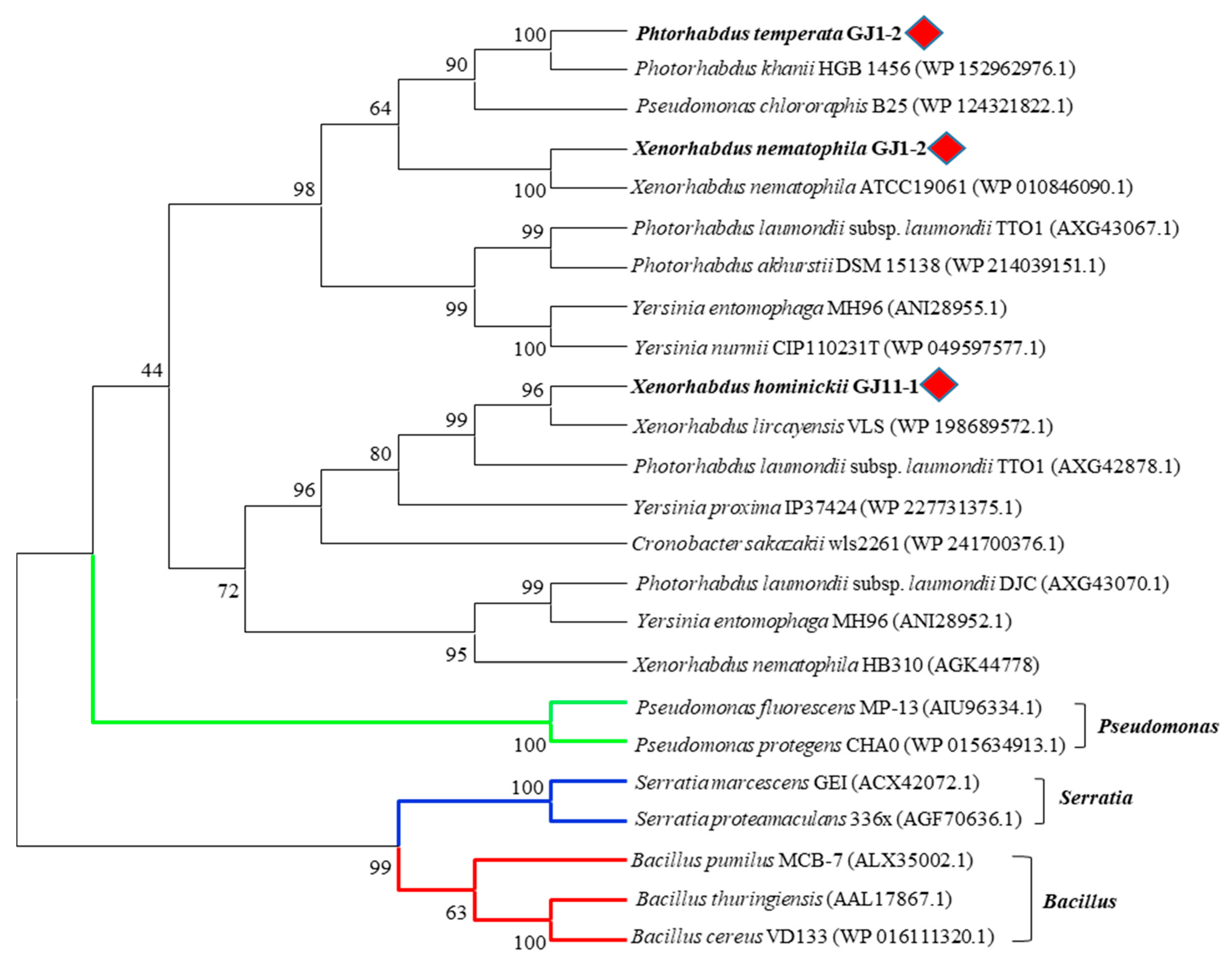
2.1. Sequencing and Phylogenetic Analysis of Chitinases of Xenorhabdus and Photorhabdus Strains Isolated from Korean Indigenous EPNs
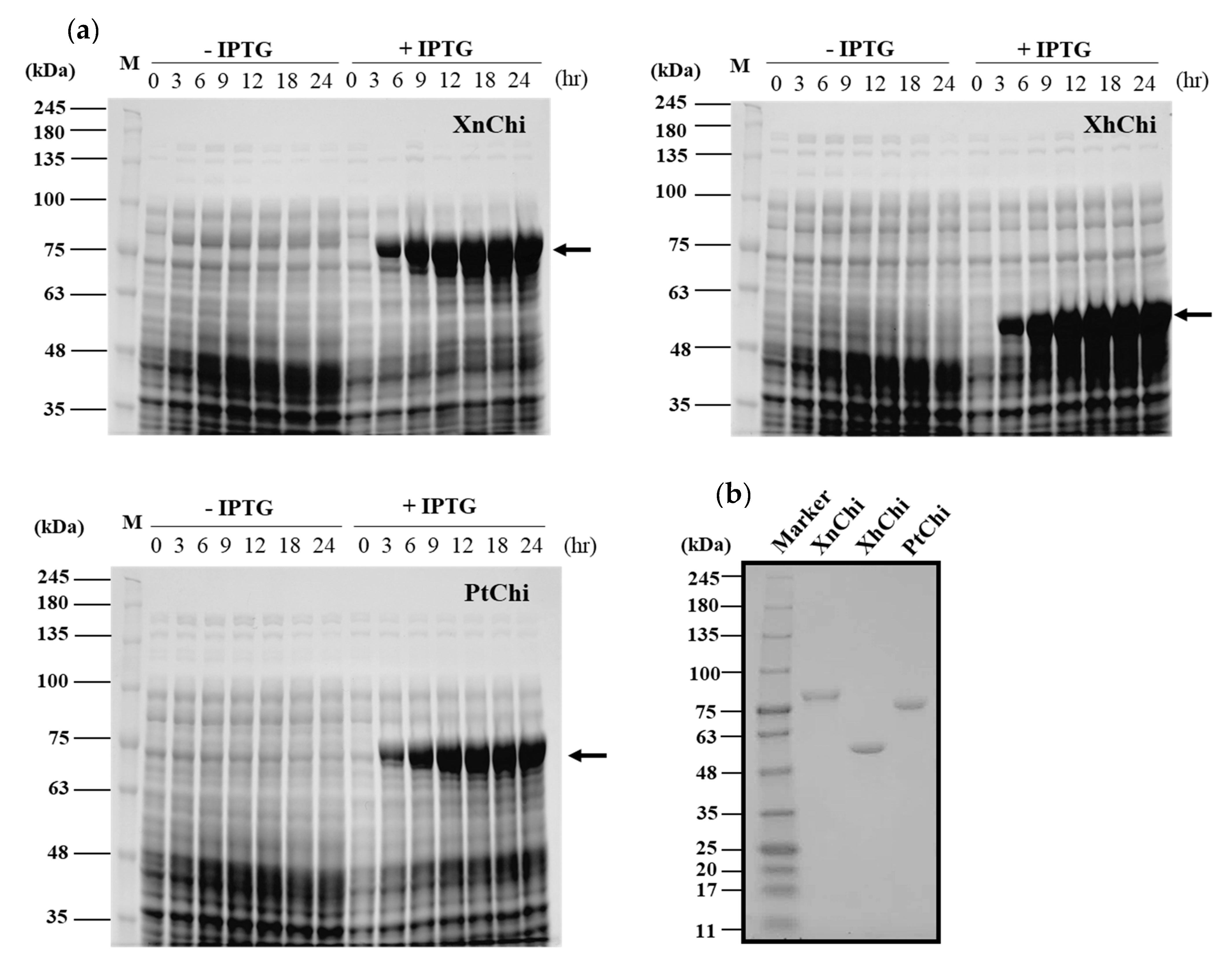
2.2. Cloning, Expression and Purification of Recombinant Chitnases
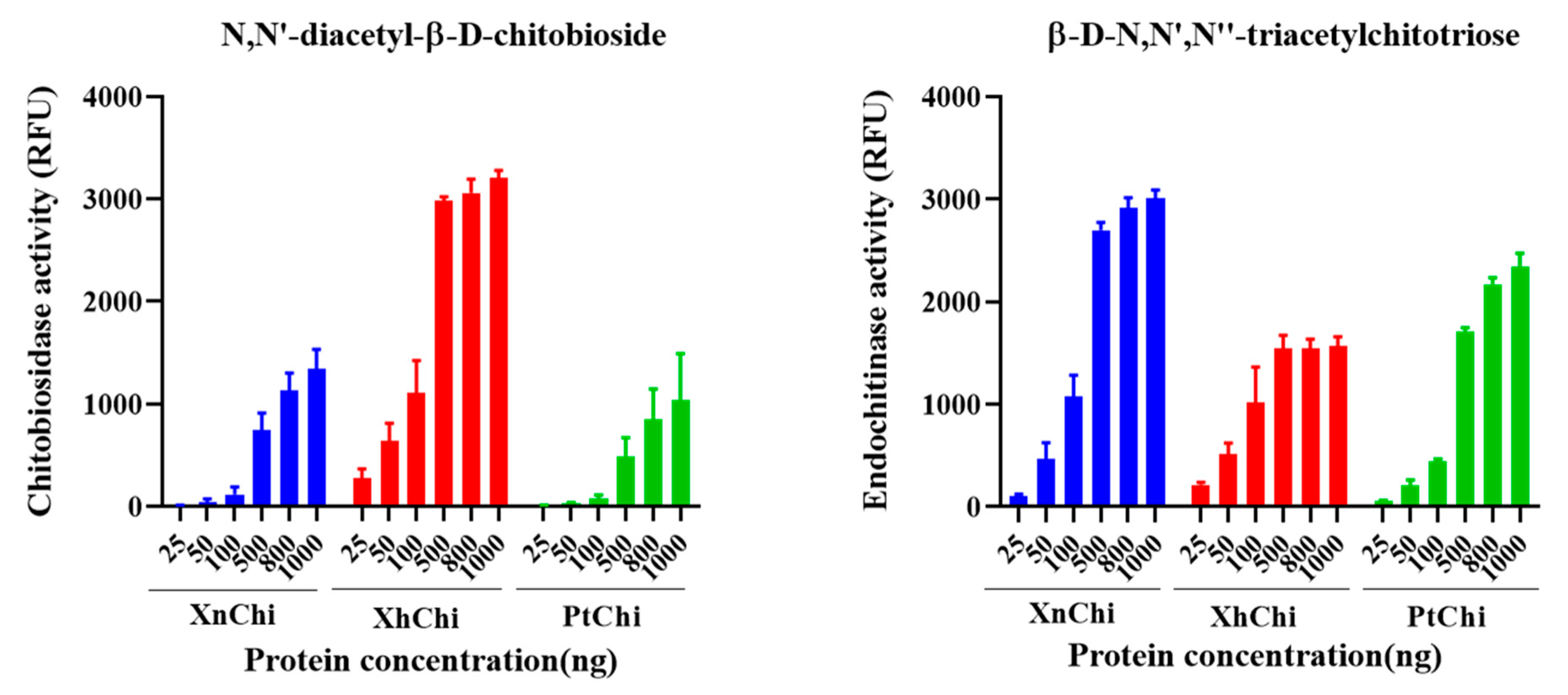
2.3. Chitinolytic Activity Assay with Recombinant Chitinases
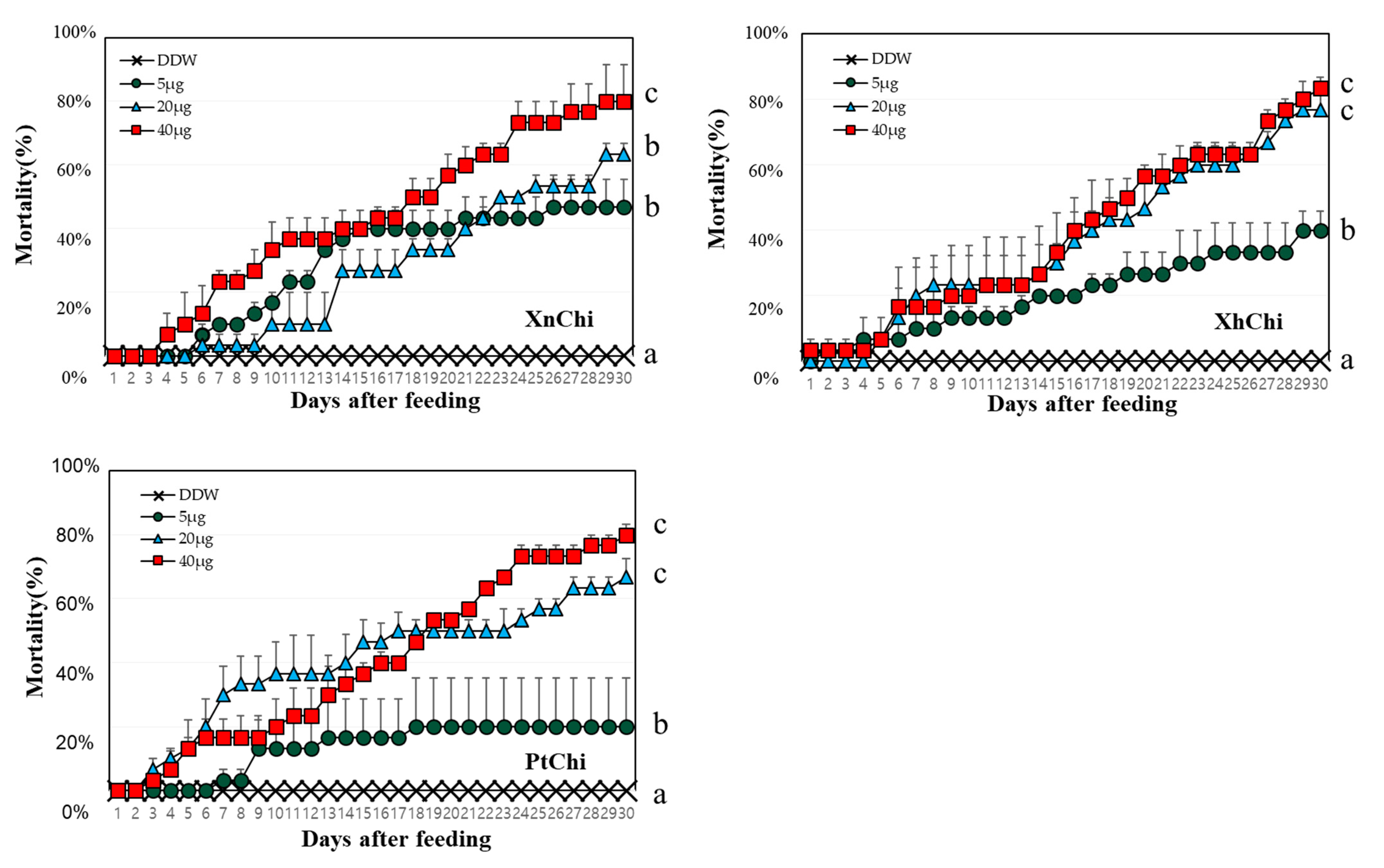
2.4. Evaluation of Insecticidal Activity of Chitinase against Galleria mellonella
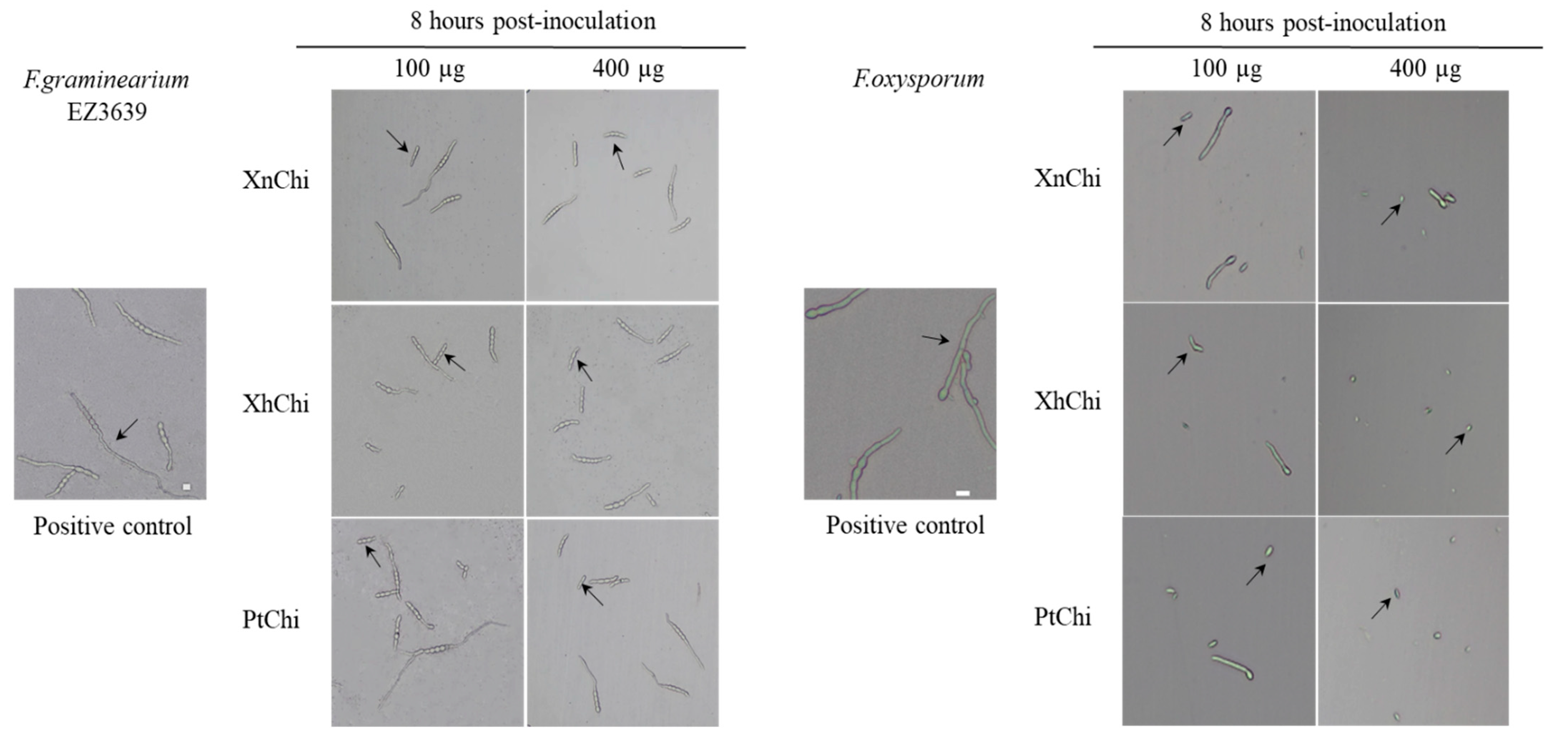
2.5. Assessment of Antifungal Activity of Chitinase Proteins against Fusarium
3. Discussion
4. Materials and Methods
4.1. Selection of Type Strain to Identify Chitinase Gene of EPN Symbiotic Bacteria
4.2. Chitinase Sequence Multi-Alignment and Phylogenetic Analysis
4.3. Construction of Recombinant Chitinase-Encoding Genes
4.4. Expression and Purification of Recombinant Chitinase Protein
4.5. Chitinolytic Activity Assay with Recombinant Chitinase Protein
4.6. Insecticidal Activity Bioassay of Recombinant Chitinase Protein against Galleria mellonella
4.7. Antifungal Activity Assay of Recombinant Chitinase Protein against Plant Pathogenic Fungi
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boemare, N. Interactions between the Partners of the Entomopathogenic Bacterium Nematode Complexes, Steinernema-Xenorhabdus and Heterorhabditis-Photorhabdus. Nematology 2002, 4, 601–603. [Google Scholar] [CrossRef]
- Choo, H.Y.; Lee, S.M.; Lee, D.W.; Kim, H.H. Ecology of entomopathogenic nematodes. In Microbial Insecticides: Principles and Applications; Borgio, J.F., Sahayaraj, K., Susurluk, I.A., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2011; ISBN 978-1-61209-223-2. [Google Scholar]
- Park, Y.; Kim, Y. Eicosanoids Rescue Spodoptera exigua Infected with Xenorhabdus nematophilus, the Symbiotic Bacteria to the Entomopathogenic Nematode Steinernema carpocapsae. J. Insect Physiol. 2000, 46, 1469–1476. [Google Scholar] [CrossRef]
- Labaude, S.; Griffin, C. Transmission Success of Entomopathogenic Nematodes Used in Pest Control. Insects 2018, 9, 72. [Google Scholar] [CrossRef]
- Kumari, P.; Mahapatro, G.K.; Banerjee, N.; Sarin, N.B. Ectopic Expression of GroEL from Xenorhabdus nematophila in Tomato Enhances Resistance against Helicoverpa armigera and Salt and Thermal Stress. Transgenic Res. 2015, 24, 859–873. [Google Scholar] [CrossRef]
- Stock, S.P.; Kusakabe, A.; Orozco, R.A. Secondary Metabolites Produced by Heterorhabditis Symbionts and Their Application in Agriculture: What We Know and What to Do Next. J. Nematol. 2017, 49, 373–383. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, J.; Liu, F.; Zeng, F. Expression of a Nematode Symbiotic Bacterium-Derived Protease Inhibitor Protein in Tobacco Enhanced Tolerance against Myzus persicae. Plant Cell Rep. 2012, 31, 1981–1989. [Google Scholar] [CrossRef]
- Sergeant, M.; Jarrett, P.; Ousley, M.; Morgan, J.A.W. Interactions of Insecticidal Toxin Gene Products from Xenorhabdus nematophilus PMFI296. Appl. Environ. Microbiol. 2003, 69, 3344–3349. [Google Scholar] [CrossRef]
- Brown, S.E.; Cao, A.T.; Dobson, P.; Hines, E.R.; Akhurst, R.J.; East, P.D. Txp40, a Ubiquitous Insecticidal Toxin Protein from Xenorhabdus and Photorhabdus Bacteria. Appl. Environ. Microbiol. 2006, 72, 1653–1662. [Google Scholar] [CrossRef]
- Vigneux, F.; Zumbihl, R.; Jubelin, G.; Ribeiro, C.; Poncet, J.; Baghdiguian, S.; Givaudan, A.; Brehélin, M. The xaxAB Genes Encoding a New Apoptotic Toxin from the Insect Pathogen Xenorhabdus nematophila Are Present in Plant and Human Pathogens. J. Biol. Chem. 2007, 282, 9571–9580. [Google Scholar] [CrossRef]
- Kumari, P.; Kant, S.; Zaman, S.; Mahapatro, G.K.; Banerjee, N.; Sarin, N.B. A Novel Insecticidal GroEL Protein from Xenorhabdus nematophila Confers Insect Resistance in Tobacco. Transgenic Res. 2014, 23, 99–107. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, J.; Li, T.; Liu, S.; Song, P.; Nangong, Z.; Wang, Q. PirAB Protein from Xenorhabdus nematophila HB310 Exhibits a Binary Toxin with Insecticidal Activity and Cytotoxicity in Galleria mellonella. J. Invertebr. Pathol. 2017, 148, 43–50. [Google Scholar] [CrossRef]
- Santhoshkumar, K.; Mathur, C.; Mandal, A.; Dutta, T.K. A Toxin Complex Protein from Photorhabdus akhurstii Conferred Oral Insecticidal Activity against Galleria mellonella by Targeting the Midgut Epithelium. Microbiol. Res. 2021, 242, 126642. [Google Scholar] [CrossRef]
- Jang, E.-K.; Kwon Jung, B.; Park, G.-S.; Rahim Khan, A.; Hong, S.-J.; Park, Y.-J.; Kim, W.-C.; Shin, J.-H.; Al-Ghamdi, K.M.S.; Oudh Al-Johny, B.; et al. Cloning and Expression of the Insecticidal Toxin Gene “tccB” from Photorhabdus temperata M1021 in Escherichia coli Expression System. J. Asia-Pac. Entomol. 2020, 23, 172–176. [Google Scholar] [CrossRef]
- Mathur, C.; Phani, V.; Kushwah, J.; Somvanshi, V.S.; Dutta, T.K. TcaB, an Insecticidal Protein from Photorhabdus akhurstii Causes Cytotoxicity in the Greater Wax Moth, Galleria mellonella. Pestic. Biochem. Physiol. 2019, 157, 219–229. [Google Scholar] [CrossRef]
- Mathur, C.; Kushwah, J.; Somvanshi, V.S.; Dutta, T.K. A 37 kDa Txp40 Protein Characterized from Photorhabdus luminescens Sub Sp. Akhurstii Conferred Injectable and Oral Toxicity to Greater Wax Moth, Galleria mellonella. Toxicon 2018, 154, 69–73. [Google Scholar] [CrossRef]
- Ullah, I.; Jang, E.-K.; Kim, M.-S.; Shin, J.-H.; Park, G.-S.; Khan, A.; Hong, S.-J.; Jung, B.-K.; Choi, J.; Park, Y.; et al. Identification and Characterization of the Insecticidal Toxin “Makes Caterpillars Floppy” in Photorhabdus temperata M1021 Using a Cosmid Library. Toxins 2014, 6, 2024–2040. [Google Scholar] [CrossRef]
- Forst, S.; Nealson, K. Molecular Biology of the Symbiotic-Pathogenic Bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol. Rev. 1996, 60, 21–43. [Google Scholar] [CrossRef]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and Chitosan: Production and Application of Versatile Biomedical Nanomaterials. Int. J. Adv. Res. 2017, 4, 411. [Google Scholar]
- Moussian, B. Chitin: Structure, Chemistry and Biology. Adv. Exp. Med. Biol. 2019, 1142, 5–18. [Google Scholar]
- Kramer, K.J.; Muthukrishnan, S. Insect Chitinases: Molecular Biology and Potential Use as Biopesticides. Insect Biochem. Mol. Biol. 1997, 27, 887–900. [Google Scholar] [CrossRef]
- Merzendorfer, H. The Cellular Basis of Chitin Synthesis in Fungi and Insects: Common Principles and Differences. Eur. J. Cell Biol. 2011, 90, 759–769. [Google Scholar] [CrossRef]
- Liu, J.; Bai, H.; Song, P.; Nangong, Z.; Dong, Z.; Li, Z.; Wang, Q. Insecticidal Activity of Chitinases from Xenorhabdus nematophila HB310 and Its Relationship with the Toxin Complex. Toxins 2022, 14, 646. [Google Scholar] [CrossRef]
- Martínez-Zavala, S.A.; Barboza-Pérez, U.E.; Hernández-Guzmán, G.; Bideshi, D.K.; Barboza-Corona, J.E. Chitinases of Bacillus Thuringiensis: Phylogeny, Modular Structure, and Applied Potentials. Front. Microbiol. 2020, 10, 3032. [Google Scholar] [CrossRef]
- He, B.; Yang, L.; Yang, D.; Jiang, M.; Ling, C.; Chen, H.; Ji, F.; Pan, L. Biochemical Purification and Characterization of a Truncated Acidic, Thermostable Chitinase from Marine Fungus for N-Acetylglucosamine Production. Front. Bioeng. Biotechnol. 2022, 10, 1013313. [Google Scholar] [CrossRef]
- Rathore, A.S.; Gupta, R.D. Chitinases from Bacteria to Human: Properties, Applications, and Future Perspectives. Enzym. Res. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Henrissat, B.; Bairoch, A. New Families in the Classification of Glycosyl Hydrolases Based on Amino Acid Sequence Similarities. Biochem. J. 1993, 293, 781–788. [Google Scholar] [CrossRef]
- Chen, W.; Jiang, X.; Yang, Q. Glycoside Hydrolase Family 18 Chitinases: The Known and the Unknown. Biotechnol. Adv. 2020, 43, 107553. [Google Scholar] [CrossRef]
- Bahar, A.A.; Sezen, K.; Demirbağ, Z.; Nalçacioğlu, R. The Relationship between Insecticidal Effects and Chitinase Activities of Coleopteran-Originated Entomopathogens and Their Chitinolytic Profile. Ann. Microbiol. 2012, 62, 647–653. [Google Scholar] [CrossRef]
- Regev, A.; Keller, M.; Strizhov, N.; Sneh, B.; Prudovsky, E.; Chet, I.; Ginzberg, I.; Koncz-Kalman, Z.; Koncz, C.; Schell, J.; et al. Synergistic Activity of a Bacillus thuringiensis δ-Endotoxin and a Bacterial Endochitinase against Spodoptera littoralis Larvae. Appl. Environ. Microbiol. 1996, 62. [Google Scholar] [CrossRef]
- Busby, J.N.; Landsberg, M.J.; Simpson, R.M.; Jones, S.A.; Hankamer, B.; Hurst, M.R.H.; Lott, J.S. Structural Analysis of Chi1 Chitinase from Yen-Tc: The Multisubunit Insecticidal ABC Toxin Complex of Yersinia entomophaga. J. Mol. Biol. 2012, 415, 359–371. [Google Scholar] [CrossRef]
- Liu, J.; NanGong, Z.; Zhang, J.; Song, P.; Tang, Y.; Gao, Y.; Wang, Q. Expression and Characterization of Two Chitinases with Synergistic Effect and Antifungal Activity from Xenorhabdus nematophila. World J. Microbiol. Biotechnol. 2019, 35, 106. [Google Scholar] [CrossRef]
- Dominelli, N.; Platz, F.; Heermann, R. The Insect Pathogen Photorhabdus luminescens Protects Plants from Phytopathogenic Fusarium Graminearum via Chitin Degradation. Appl. Environ. Microbiol. 2022, 88, e00645-22. [Google Scholar] [CrossRef]
- Sajnaga, E.; Kazimierczak, W. Evolution and Taxonomy of Nematode-Associated Entomopathogenic Bacteria of the Genera Xenorhabdus and Photorhabdus: An Overview. Symbiosis 2020, 80, 1–13. [Google Scholar] [CrossRef]
- Boemare, N.; Akhurst, R. The Genera Photorhabdus and Xenorhabdus. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 451–494. ISBN 978-0-387-25496-8. [Google Scholar]
- Hinchliffe, S.J. Insecticidal Toxins from the Photorhabdus and Xenorhabdus Bacteria. Open Toxinol. J. 2013, 3, 101–118. [Google Scholar] [CrossRef]
- Fukruksa, C.; Yimthin, T.; Suwannaroj, M.; Muangpat, P.; Tandhavanant, S.; Thanwisai, A.; Vitta, A. Isolation and Identification of Xenorhabdus and Photorhabdus Bacteria Associated with Entomopathogenic Nematodes and Their Larvicidal Activity against Aedes aegypti. Parasit. Vectors 2017, 10, 440. [Google Scholar] [CrossRef]
- Mollah, M.M.I.; Roy, M.C.; Choi, D.-Y.; Hasan, M.A.; Al Baki, M.A.; Yeom, H.-S.; Kim, Y. Variations of Indole Metabolites and NRPS-PKS Loci in Two Different Virulent Strains of Xenorhabdus hominickii. Front. Microbiol. 2020, 11, 583594. [Google Scholar] [CrossRef]
- Rivera-Ramírez, A.; Salgado-Morales, R.; Jiménez-Pérez, A.; Pérez-Martínez, R.; García-Gómez, B.I.; Dantán-González, E. Comparative Genomics and Pathogenicity Analysis of Two Bacterial Symbionts of Entomopathogenic Nematodes: The Role of The GroEL Protein in Virulence. Microorganisms 2022, 10, 486. [Google Scholar] [CrossRef]
- Mahmood, S.; Kumar, M.; Kumari, P.; Mahapatro, G.K.; Banerjee, N.; Sarin, N.B. Novel Insecticidal Chitinase from the Insect Pathogen Xenorhabdus nematophila. Int. J. Biol. Macromol. 2020, 159, 394–401. [Google Scholar] [CrossRef]
- Palmer, I.; Wingfield, P.T. Preparation and Extraction of Insoluble (Inclusion-Body) Proteins from Escherichia coli. Curr. Protoc. Protein Sci. 2012, 70, 6.3.1–6.3.20. [Google Scholar] [CrossRef]
- Singh, G.; Arya, S.K. Antifungal and Insecticidal Potential of Chitinases: A Credible Choice for the Eco-Friendly Farming. Biocatal. Agric. Biotechnol. 2019, 20, 101289. [Google Scholar] [CrossRef]
- Prasanna, L.; Eijsink, V.G.H.; Meadow, R.; Gåseidnes, S. A Novel Strain of Brevibacillus laterosporus Produces Chitinases That Contribute to Its Biocontrol Potential. Appl. Microbiol. Biotechnol. 2013, 97, 1601–1611. [Google Scholar] [CrossRef]
- Suganthi, M.; Senthilkumar, P.; Arvinth, S.; Chandrashekara, K.N. Chitinase from Pseudomonas Fluorescens and Its Insecticidal Activity against Helopeltis theivora. J. Gen. Appl. Microbiol. 2017, 63, 222–227. [Google Scholar] [CrossRef]
- Stutz, E.W.; Défago, G.; Kern, H. Naturally Occurring Fluorescent Pseudomonads Involved in Suppression. Phytopathology 1986, 76, 181–185. [Google Scholar] [CrossRef]
- Tu, S.; Qiu, X.; Cao, L.; Han, R.; Zhang, Y.; Liu, X. Expression and Characterization of the Chitinases from Serratia Marcescens GEI Strain for the Control of Varroa Destructor, a Honey Bee Parasite. J. Invertebr. Pathol. 2010, 104, 75–82. [Google Scholar] [CrossRef]
- Wang, M.; Xing, Y.; Wang, J.; Xu, Y.; Wang, G. The Role of the Chi1 Gene from the Endophytic Bacteria Serratia proteamaculans 336x in the Biological Control of Wheat Take-All. Can. J. Microbiol. 2014, 60, 533–540. [Google Scholar] [CrossRef]
- Rishad, K.S.; Varghese, S.; Jisha, M.S. Sequence Analysis and Docking Performance of Extracellular Chitinase from Bacillus Pumilus MCB-7, a Novel Mangrove Isolate. Enzym. Microb. Technol. 2020, 140, 109624. [Google Scholar] [CrossRef]
- Barboza-Corona, J.E.; Nieto-Mazzocco, E.; Velázquez-Robledo, R.; Salcedo-Hernandez, R.; Bautista, M.; Jiménez, B.; Ibarra, J.E. Cloning, Sequencing, and Expression of the Chitinase Gene chiA74 from Bacillus thuringiensis. Appl. Environ. Microbiol. 2003, 69, 1023–1029. [Google Scholar] [CrossRef]
- Russell, J.; Kim, S.-K.; Duma, J.; Nothaft, H.; Himmel, M.E.; Bomble, Y.J.; Szymanski, C.M.; Westpheling, J. Deletion of a Single Glycosyltransferase in Caldicellulosiruptor bescii Eliminates Protein Glycosylation and Growth on Crystalline Cellulose. Biotechnol. Biofuels 2018, 11, 259. [Google Scholar] [CrossRef]
- Boysen, A.; Palmisano, G.; Krogh, T.J.; Duggin, I.G.; Larsen, M.R.; Møller-Jensen, J. A Novel Mass Spectrometric Strategy “BEMAP” Reveals Extensive O-Linked Protein Glycosylation in Enterotoxigenic Escherichia coli. Sci. Rep. 2016, 6, 32016. [Google Scholar] [CrossRef]
- Beier, S.; Bertilsson, S. Bacterial Chitin Degradation—Mechanisms and Ecophysiological Strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef]
- Oyeleye, A.; Normi, Y.M. Chitinase: Diversity, Limitations, and Trends in Engineering for Suitable Applications. Biosci. Rep. 2018, 38, BSR2018032300. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.; Li, J.; Dunphy, G.B.; Punja, Z.K.; Webster, J.M. Chitinase Activity of Xenorhabdus and Photorhabdus Species, Bacterial Associates of Entomopathogenic Nematodes. J. Invertebr. Pathol. 1996, 68, 101–108. [Google Scholar] [CrossRef]
- Berini, F.; Casartelli, M.; Montali, A.; Reguzzoni, M.; Tettamanti, G.; Marinelli, F. Metagenome-Sourced Microbial Chitinases as Potential Insecticide Proteins. Front. Microbiol. 2019, 10, 1358. [Google Scholar] [CrossRef]
- Churklam, W.; Aunpad, R. Enzymatic Characterization and Structure-Function Relationship of Two Chitinases, LmChiA and LmChiB, from Listeria monocytogenes. Heliyon 2020, 6, e04252. [Google Scholar] [CrossRef]
- Roberts, W.K.; Selitrennikoff, C.P. Plant and Bacterial Chitinases Differ in Antifungal Activity. Microbiology 1988, 134, 169–176. [Google Scholar] [CrossRef]
- Lorito, M.; Harman, G.E.; Hayes, C.K.; Broadway, R.M.; Tronsmo, A.; Woo, S.L.; Di Pietro, A. Chitinolytic Enzymes Produced by Trichoderma harzianum: Antifungal Activity of Purified Endochitinase and Chitobiosidase. Phytopathology 1993, 83, 302–307. [Google Scholar] [CrossRef]
- Hjort, K.; Presti, I.; Elväng, A.; Marinelli, F.; Sjöling, S. Bacterial Chitinase with Phytopathogen Control Capacity from Suppressive Soil Revealed by Functional Metagenomics. Appl. Microbiol. Biotechnol. 2014, 98, 2819–2828. [Google Scholar] [CrossRef]
- Jiménez-Ortega, E.; Kidibule, P.E.; Fernández-Lobato, M.; Sanz-Aparicio, J. Structure–Function Insights into the Fungal Endo-Chitinase Chit33 Depict Its Mechanism on Chitinous Material. Int. J. Mol. Sci. 2022, 23, 7599. [Google Scholar] [CrossRef]
- Koga, D.; Funakoshi, T.; Mizuki, K.; Ide, A.; Kramer, K.J.; Zen, K.-C.; Choi, H.; Muthukrishnan, S. Immunoblot Analysis of Chitinolytic Enzymes in Integument and Molting Fluid of the Silkworm, Bombyx Mori, and the Tobacco Hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 1992, 22, 305–311. [Google Scholar] [CrossRef]
- Koga, D.; Jilka, J.; Kramer, K.J. Insect Endochitinases: Glycoproteins from Moulting Fluid, Integument and Pupal Haemolymph of Manduca sexta L. Insect Biochem. 1983, 13, 295–305. [Google Scholar] [CrossRef]
- Navarro-González, S.S.; Ramírez-Trujillo, J.A.; Peña-Chora, G.; Gaytán, P.; Roldán-Salgado, A.; Corzo, G.; Lina-García, L.P.; Hernández-Velázquez, V.M.; Suárez-Rodríguez, R. Enhanced Tolerance against a Fungal Pathogen and Insect Resistance in Transgenic Tobacco Plants Overexpressing an Endochitinase Gene from Serratia marcescens. Int. J. Mol. Sci. 2019, 20, 3482. [Google Scholar] [CrossRef]
- Zarei, M.; Aminzadeh, S.; Zolgharnein, H.; Safahieh, A.; Daliri, M.; Noghabi, K.A.; Ghoroghi, A.; Motallebi, A. Characterization of a Chitinase with Antifungal Activity from a Native Serratia marcescens B4A. Braz. J. Microbiol. 2011, 42, 1017–1029. [Google Scholar] [CrossRef]
- Schoffelmeer, E.A.M.; Klis, F.M.; Sietsma, J.H.; Cornelissen, B.J.C. The Cell Wall of Fusarium oxysporum. Fungal Genet. Biol. 1999, 27, 275–282. [Google Scholar] [CrossRef]
- Barbosa, I.P.; Kemmelmeier, C. Chemical Composition of the Hyphal Wall from Fusarium graminearum. Exp. Mycol. 1993, 17, 274–283. [Google Scholar] [CrossRef]
- Liu, M.; Gong, Y.; Sun, H.; Zhang, J.; Zhang, L.; Sun, J.; Han, Y.; Huang, J.; Wu, Q.; Zhang, C.; et al. Characterization of a Novel Chitinase from Sweet Potato and Its Fungicidal Effect against Ceratocystis fimbriata. J. Agric. Food Chem. 2020, 68, 7591–7600. [Google Scholar] [CrossRef]
- Kotb, E.; Alabdalall, A.H.; Alghamdi, A.I.; Ababutain, I.M.; Aldakeel, S.A.; Al-Zuwaid, S.K.; Algarudi, B.M.; Algarudi, S.M.; Ahmed, A.A.; Albarrag, A.M. Screening for Chitin Degrading Bacteria in the Environment of Saudi Arabia and Characterization of the Most Potent Chitinase from Streptomyces Variabilis Am1. Sci. Rep. 2023, 13, 11723. [Google Scholar] [CrossRef]
- Rodou, A.; Ankrah, D.O.; Stathopoulos, C. Toxins and Secretion Systems of Photorhabdus luminescens. Toxins 2010, 2, 1250–1264. [Google Scholar] [CrossRef]
- Sheets, J.J.; Hey, T.D.; Fencil, K.J.; Burton, S.L.; Ni, W.; Lang, A.E.; Benz, R.; Aktories, K. Insecticidal Toxin Complex Proteins from Xenorhabdus nematophilus. J. Biol. Chem. 2011, 286, 22742–22749. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Dowling, A.; Waterfield, N.R. Insecticidal Toxins from Photorhabdus Bacteria and Their Potential Use in Agriculture. Toxicon 2007, 49, 436–451. [Google Scholar] [CrossRef]
- Mahmood, S.; Kumari, P.; Kisku, A.V.; Mahapatro, G.K.; Banerjee, N.; Sarin, N.B. Ectopic Expression of Xenorhabdus nematophila Chitinase in Tobacco Confers Resistance against Helicoverpa armigera. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 151, 593–604. [Google Scholar] [CrossRef]
- Cappellini, R.A.; Peterson, J.L. Macroconidium Formation in Submerged Cultures by a Nonsporulating Strain of Gibberella zeae. Mycologia 1965, 57, 962–966. [Google Scholar] [CrossRef]
- Li, T.; Jung, B.; Park, S.-Y.; Lee, J. Survival Factor Gene FgSvf1 Is Required for Normal Growth and Stress Resistance in Fusarium graminearum. Plant Pathol. J. 2019, 35, 393–405. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Time to Death and/or Death Response (40 μg/Diet-Fed Larva) | ||
|---|---|---|---|
| LT50 (days) | LT70 (days) | R2 Value | |
| XnChi | 16.155 (14.066–18.661) | 25.289 (21.543–31.838) | 0.964 |
| XhChi | 18.337 (15.879–21.666) | 29.433 (24.460–38.900) | 0.880 |
| PtChi | 17.797 (15.384–21.307) | 27.841 (22.944–37.917) | 0.932 |
| Protein | Toxicity in mg/mL to F. graminearum EZ3639 | ||
|---|---|---|---|
| 8 h | |||
| IC50 | IC70 | R2 Value | |
| XnChi | 0.055 (0.015–0.089) | 0.325 (0.201–1.431) | 0.987 |
| XhChi | 0.040 (0.010–0.068) | 0.200 (0.135–0.392) | 0.956 |
| PtChi | 0.133 (0.080–0.210) | 0.742 (0.382–5.226) | 0.949 |
| Protein | Toxicity in mg/mL to F. oxysporum | ||
| 8 h | |||
| IC50 | IC70 | R2 Value | |
| XnChi | 0.072 (0.042–0.098) | 0.239 (0.175–0.397) | 0.938 |
| XhChi | 0.031 (0.004–0.060) | 0.191 (0.122–0.423) | 0.971 |
| PtChi | 0.046 (0.024–0.066) | 0.609 (0.378–1.562) | 0.931 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, D.-J.; Kim, G.-G.; Choo, H.-Y.; Chung, N.-J.; Choo, Y.-M. Functional Comparison of Three Chitinases from Symbiotic Bacteria of Entomopathogenic Nematodes. Toxins 2024, 16, 26. https://doi.org/10.3390/toxins16010026
Son D-J, Kim G-G, Choo H-Y, Chung N-J, Choo Y-M. Functional Comparison of Three Chitinases from Symbiotic Bacteria of Entomopathogenic Nematodes. Toxins. 2024; 16(1):26. https://doi.org/10.3390/toxins16010026
Chicago/Turabian StyleSon, Da-Jeong, Geun-Gon Kim, Ho-Yul Choo, Nam-Jun Chung, and Young-Moo Choo. 2024. "Functional Comparison of Three Chitinases from Symbiotic Bacteria of Entomopathogenic Nematodes" Toxins 16, no. 1: 26. https://doi.org/10.3390/toxins16010026
APA StyleSon, D.-J., Kim, G.-G., Choo, H.-Y., Chung, N.-J., & Choo, Y.-M. (2024). Functional Comparison of Three Chitinases from Symbiotic Bacteria of Entomopathogenic Nematodes. Toxins, 16(1), 26. https://doi.org/10.3390/toxins16010026