Tetrodotoxin/Saxitoxin Accumulation Profile in the Euryhaline Marine Pufferfish Chelonodontops patoca

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
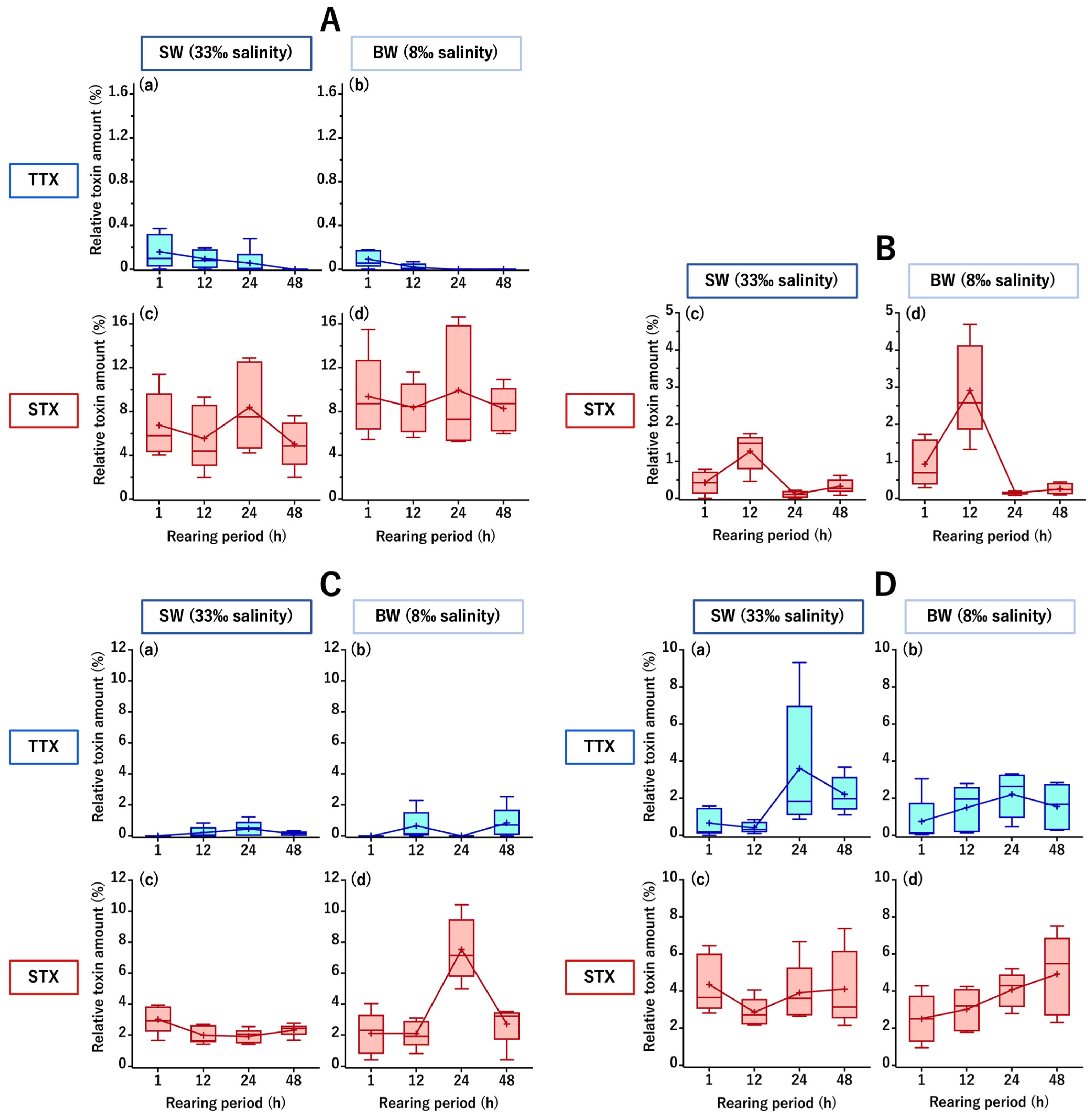
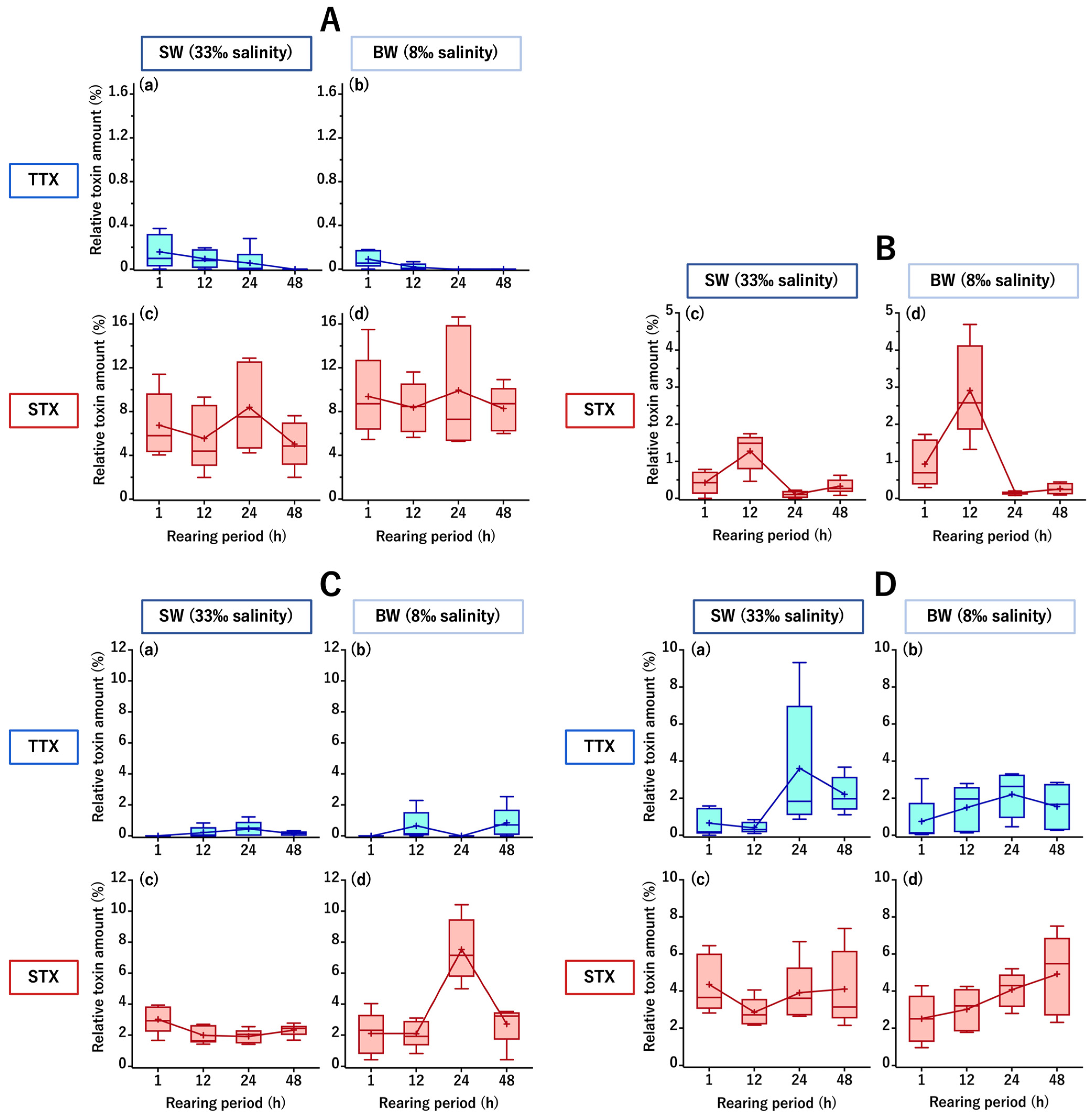
2.1. TTX/STX Accumulation Profile of C. patoca
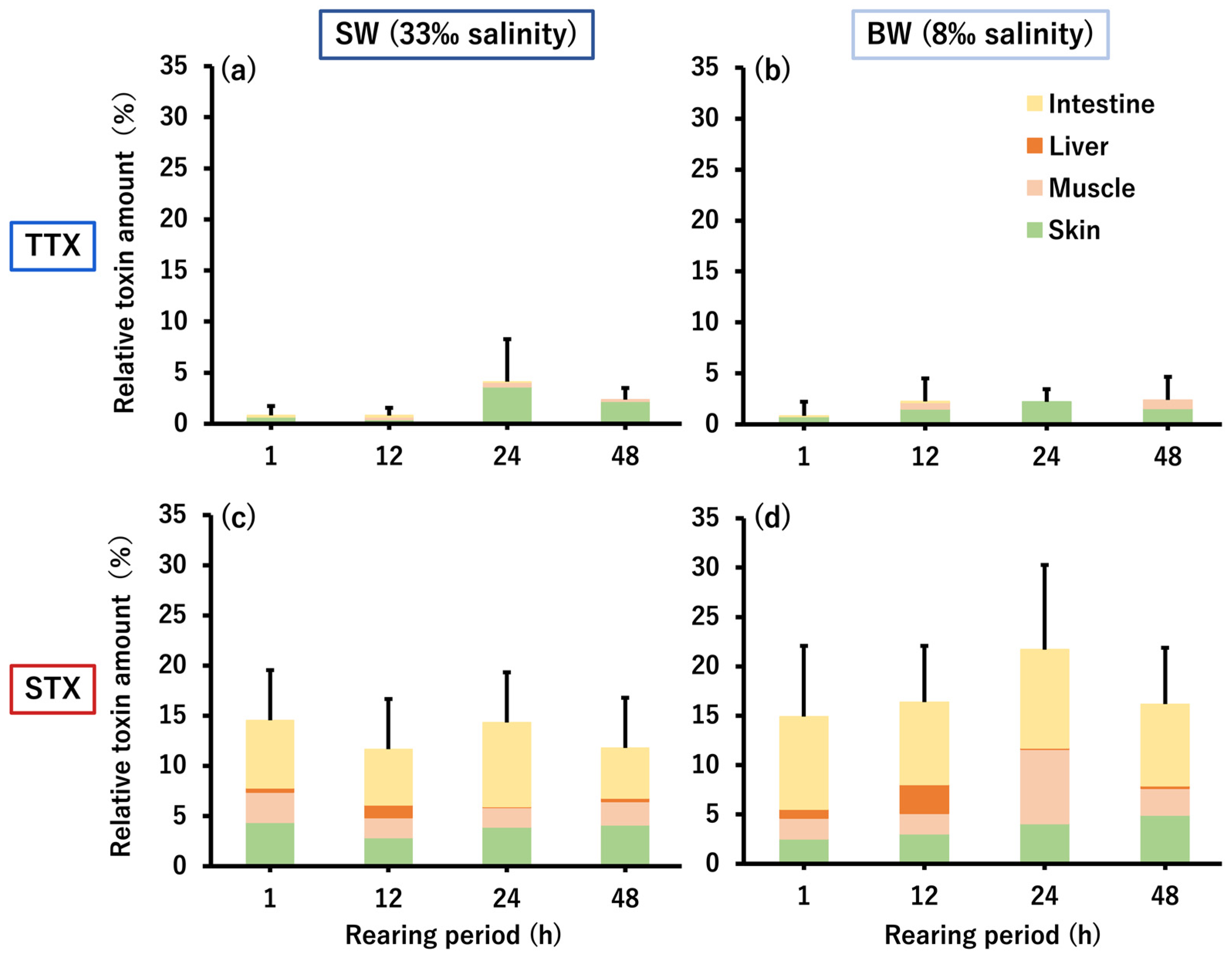
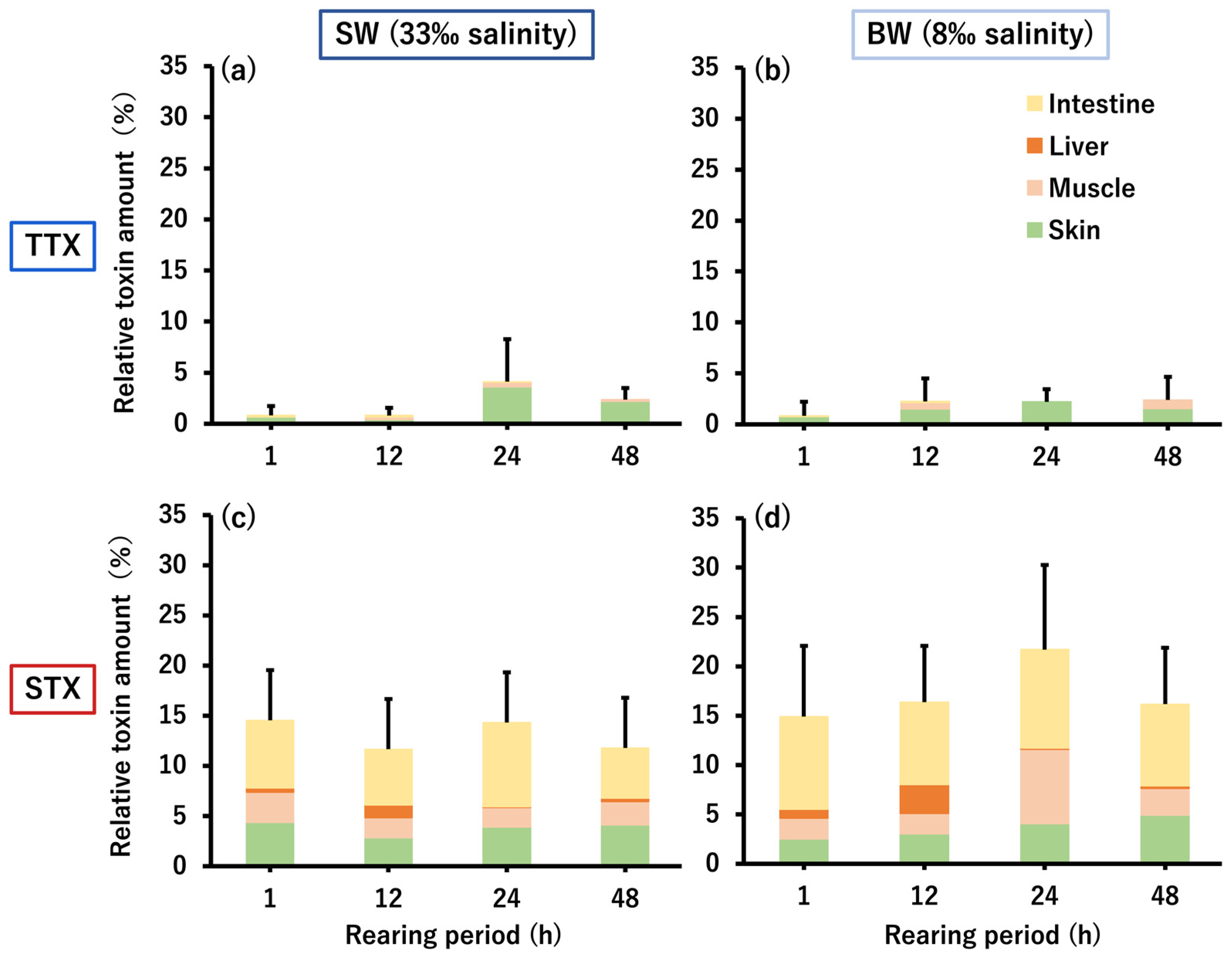
2.2. Micro-Distribution of PSTBPs and Accumulated TTX/STX in the Skin
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Pufferfish Specimens and Acclimation Procedures
5.2. Toxin Preparation
5.3. Intrarectal Toxin Administration Experiment
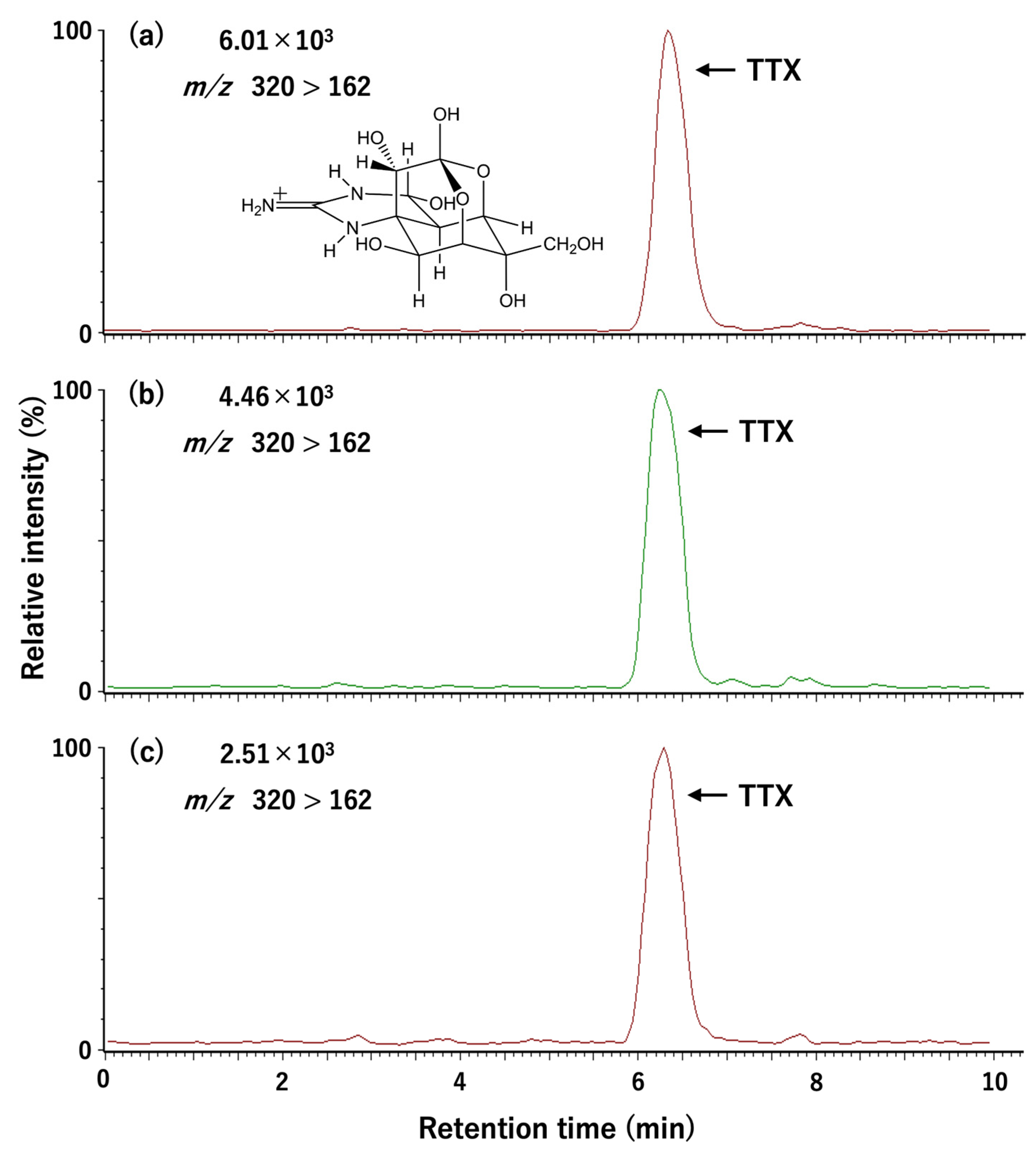
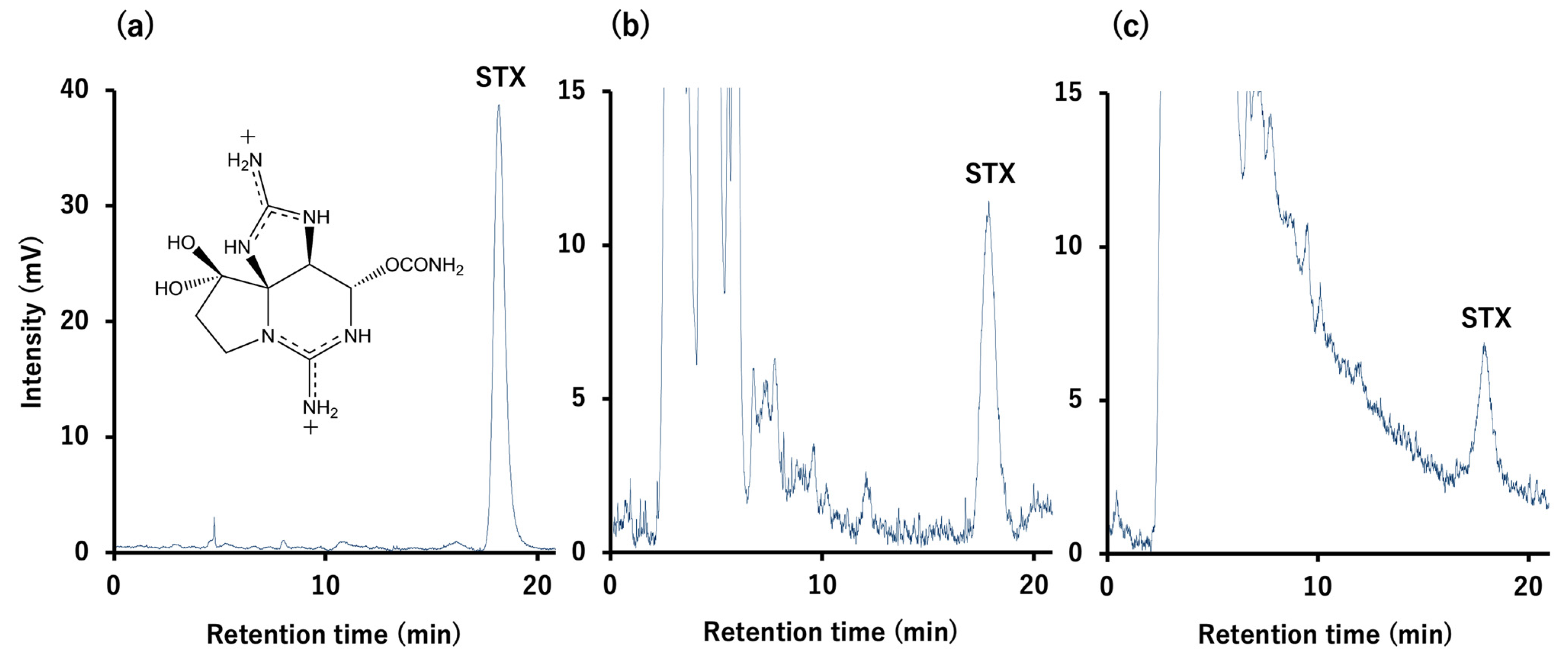
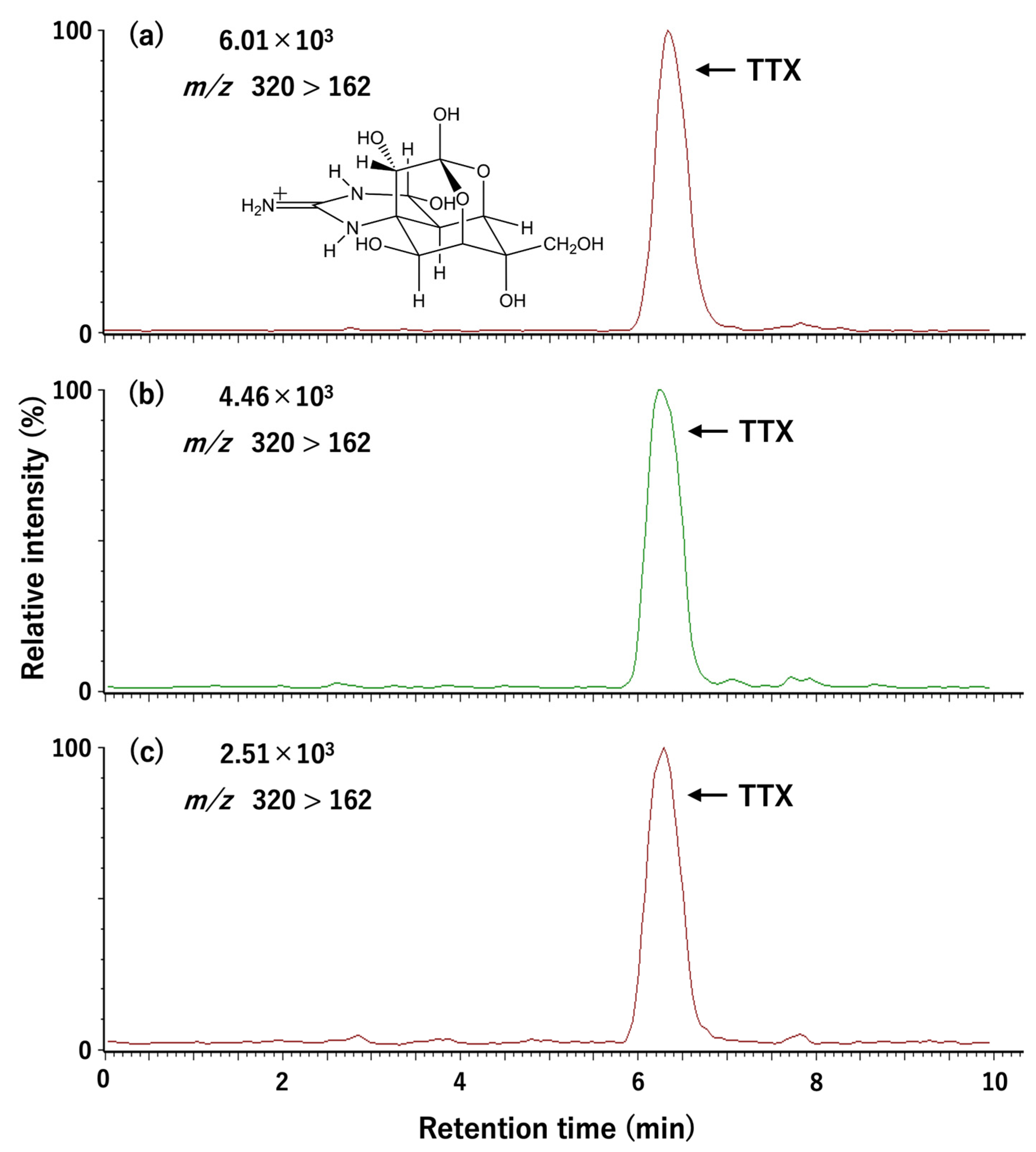
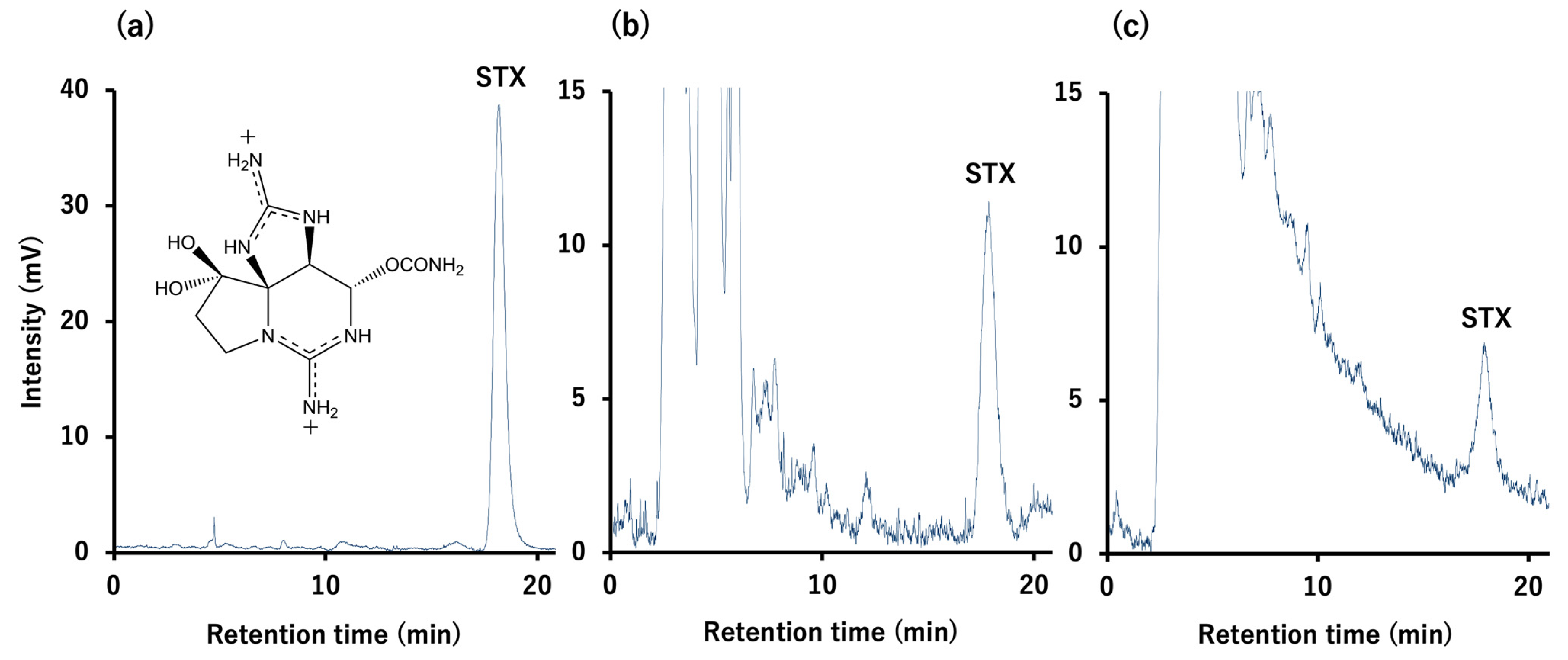
5.4. Toxin Quantification
5.5. Immunohistochemistry and Immunofluorescence Staining
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef]
- Göransson, U.; Jacobsson, E.; Strand, M.; Andersson, H.S. The toxins of nemertean worms. Toxins 2019, 11, 120. [Google Scholar] [CrossRef]
- Oyama, H.; Ito, M.; Suo, R.; Goto-Inoue, N.; Morisasa, M.; Mori, T.; Sugita, H.; Mori, T.; Nakahigashi, R.; Adachi, M.; et al. Changes in tissue distribution of tetrodotoxin and its analogues in association with maturation in the toxic flatworm, Planocera multitentaculata. Mar. Biotechnol. 2022, 24, 1158–1167. [Google Scholar] [CrossRef]
- Melnikova, D.I.; Magarlamov, T.Y. An overview of the anatomical distribution of tetrodotoxin in animals. Toxins 2022, 14, 576. [Google Scholar] [CrossRef]
- Ueyama, N.; Sugimoto, K.; Kudo, Y.; Onodera, K.I.; Cho, Y.; Konoki, K.; Nishikawa, T.; Yotsu-Yamashita, M. Spiro bicyclic guanidino compounds from pufferfish: Possible biosynthetic intermediates of tetrodotoxin in marine environments. Chem. Eur. J. 2018, 24, 7250–7258. [Google Scholar] [CrossRef]
- Itoi, S.; Kozaki, A.; Komori, K.; Tsunashima, T.; Noguchi, S.; Kawane, M.; Sugita, H. Toxic Takifugu pardalis eggs found in Takifugu niphobles gut: Implications for TTX accumulation in the pufferfish. Toxicon 2015, 108, 141–146. [Google Scholar] [CrossRef]
- Itoi, S.; Ishizuka, K.; Mitsuoka, R.; Takimoto, N.; Yokoyama, N.; Detake, A.; Takayanagi, C.; Yoshikawa, S.; Sugita, H. Seasonal changes in the tetrodotoxin content of the pufferfish Takifugu niphobles. Toxicon 2016, 114, 53–58. [Google Scholar] [CrossRef]
- Tatsuno, R.; Umeeda, M.; Miyata, Y.; Ideguchi, R.; Fukuda, T.; Furushita, M.; Ino, Y.; Yoshikawa, H.; Takahashi, H.; Nagashima, Y. Toxicity of Takifugu exascurus collected from the Sea of Kumano. Shokuhin Eiseigaku Zasshi 2021, 62, 28–32. [Google Scholar] [CrossRef]
- Jal, S.; Khora, S.S. An overview on the origin and production of tetrodotoxin, a potent neurotoxin. J. Appl. Microbiol. 2015, 119, 907–916. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins 2017, 9, 166. [Google Scholar] [CrossRef]
- Melnikova, D.I.; Vlasenko, A.E.; Magarlamov, T.Y. Stable tetrodotoxin production by Bacillus sp. Strain 1839. Mar. Drugs 2019, 17, 704. [Google Scholar] [CrossRef]
- Arakawa, O.; Takatani, T.; Taniyama, S.; Tatsuno, R. Toxins of pufferfish—Distribution, accumulation mechanism, and physiologic functions. Aqua-BioScience Monogr. 2017, 10, 41–80. [Google Scholar] [CrossRef]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef]
- Tani, T. Nihonsan Fugu no Chudokugakuteki Kenkyu (Toxicological Studies on Japanese Puffer); Teikoku Tosho: Tokyo, Japan, 1945. [Google Scholar]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: Epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–233. [Google Scholar] [CrossRef]
- Deeds, J.; Landsberg, J.; Etheridge, S.; Pitcher, G.; Longan, S. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Forty years of sodium channels: Structure, function, pharmacology, and epilepsy. Neurochem. Res. 2017, 42, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Vilariño, N.; Vale, C.; Costas, C.; Cao, A.; Raposo-García, S.; Vieytes, M.R.; Botana, L.M. Current trends and new challenges in marine phycotoxins. Mar. Drugs 2022, 20, 198. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kodama, M.; Ogata, T.; Saitanu, K.; Furuya, M.; Hirayama, K.; Kamimura, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Yamada, A.; Goto, Y.; Horn, L.; Ngy, L.; Wada, M.; Doi, H.; Lee, J.S.; Takatani, T.; Arakawa, O. Phylogeny and toxin profile of freshwater pufferfish (genus Pao) collected from 2 different regions in Cambodia. Toxins 2020, 12, 689. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Ogata, T.; Borja, V.; Gonzales, C.; Fukuyo, Y.; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef]
- Barrientos, R.G.; Hernández-Mora, G.; Alegre, F.; Field, T.; Flewelling, L.; McGrath, S.; Deeds, J.; Chacón, Y.S.; Arrieta, K.R.; Vargas, E.C.; et al. Saxitoxin poisoning in green turtles (Chelonia mydas) linked to scavenging on mass mortality of Caribbean sharpnose puffer fish (Canthigaster rostrata-Tetraodontidae). Front. Vet. Sci. 2019, 6, 466. [Google Scholar] [CrossRef]
- Li, J.; Persson, K.M. Quick detection method for paralytic shellfish toxins (PSTs) monitoring in freshwater-a review. Chemosphere 2021, 265, 128591. [Google Scholar] [CrossRef]
- Zhu, H.; Sonoyama, T.; Yamada, M.; Gao, W.; Tatsuno, R.; Takatani, T.; Arakawa, O. Co-occurrence of tetrodotoxin and saxitoxins and their intra-body distribution in the pufferfish Canthigaster valentini. Toxins 2020, 12, 436. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H.; Doi, H.; Takatani, T.; Arakawa, O. Contrasting toxin selectivity between the marine pufferfish Takifugu pardalis and the freshwater pufferfish Pao suvattii. Toxins 2019, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, Y.; Toyoda, M.; Hasobe, M.; Shimakura, K.; Shiomi, K. In vitro accumulation of tetrodotoxin in pufferfish liver tissue slices. Toxicon 2003, 41, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Nagashima, Y.; Takayama, K.; Shiomi, K. Difference between tetrodotoxin and saxitoxins in accumulation in puffer fish Takifugu rubripes liver tissue slices. Fish Physiol. Biochem. 2005, 31, 95–100. [Google Scholar] [CrossRef]
- Nagashima, Y.; Ohta, A.; Yin, X.; Ishizaki, S.; Matsumoto, T.; Doi, H.; Ishibashi, T. Difference in uptake of tetrodotoxin and saxitoxins into liver tissue slices among pufferfish, boxfish and porcupinefish. Mar. Drugs 2018, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Sakai, T.; Nagashima, Y.; Doi, H.; Takatani, T.; Arakawa, O. Tetrodotoxin/saxitoxins selectivity of the euryhaline freshwater pufferfish Dichotomyctere fluviatilis. Toxins 2021, 13, 731. [Google Scholar] [CrossRef] [PubMed]
- Talwar, P.K.; Jhingran, A.G. Inland Fishes of India and Adjacent Countries; A.A. Balkema: Rotterdam, The Netherlands, 1992; Volume 2, p. 1057. [Google Scholar]
- Mahmud, Y.; Tanu, M.B.; Takatani, T.; Asayama, E.; Arakawa, O.; Noguchi, T. Chelonodon patoca, a highly toxic marine puffer in Japan. J. Nat. Toxins 2001, 10, 69–74. [Google Scholar]
- Khora, S.S.; Isa, J.; Yasumoto, T. Toxicity of puffers from Okinawa, Japan. Nippon Suisan Gakkaishi 1991, 57, 163–167. [Google Scholar] [CrossRef]
- Nakamura, M.; Yasumoto, T. Tetrodotoxin derivatives in puffer fish. Toxicon 1985, 23, 271–276. [Google Scholar] [CrossRef]
- Shimizu, Y.; Yoshioka, M. Transformation of paralytic shellfish toxins as demonstrated in scallop homogenates. Science 1981, 212, 547–549. [Google Scholar] [CrossRef]
- Oshima, Y. Chemical and enzymatic transformation of paralytic shellfish toxins in marine organisms. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier: Paris, France, 1995; pp. 475–480. [Google Scholar]
- Santos, I.A.; Castellano, G.C.; Freire, C.A. Direct relationship between osmotic and ionic conforming behavior and tissue water regulatory capacity in echinoids. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Morera, F.J.; Baez-Nieto, D.; Lorenzo-Ceballos, Y.; Castillo, K.; Pupo, A.; Vargas-Chacoff, L.; Gonzalez, C. Role of ion channels in salt secretion by Atlantic salmon gills during acclimation to seawater. Physiol. Mini-Rev. 2015, 8, 1–11. Available online: http://sedici.unlp.edu.ar/handle/10915/127755 (accessed on 18 November 2023).
- Bœuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. Part C 2001, 130, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Sakakura, Y.; Takeshige, R.; Takatani, T.; Arakawa, O.; Kaneko, T.; Amano, M. Tetrodotoxin accumulation conflicts with low salinity tolerance in juvenile tiger puffer Takifugu rubripes. Fish. Sci. 2023, 89, 21–28. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Nagaoka, Y.; Muramoto, K.; Cho, Y.; Konoki, K. Pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) analogues in the blood plasma of the pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca. Mar. Drugs 2018, 16, 224. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Yamaki, H.; Okoshi, N.; Araki, N. Distribution of homologous proteins to puffer fish saxitoxin and tetrodotoxin binding protein in the plasma of pufferfish and among the tissues of Fugu pardalis examined by Western blot analysis. Toxicon 2010, 55, 1119–1124. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Lee, J.M.; Shiraishi, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, N.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103–1118. [Google Scholar] [CrossRef]
- Oba, Y.; Shimasaki, Y.; Oshima, Y.; Satone, H.; Kitano, T.; Nakao, M.; Kawabata, S.; Honjo, T. Purification and characterization of tributyltin-binding protein type 2 from plasma of Japanese flounder, Paralichthys olivaceus. J. Biochem. 2007, 142, 229–238. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Yoda, T.; Terano, Y.; Shibata, T. Rapid and highly sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn. J. Med. Sci. Biol. 1997, 50, 133–150. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Sugiyama, A.; Hashizume, K.; Noguchi, T. Development and application of an enzyme immunoassay based on a monoclonal antibody against gonyautoxin components of paralytic shellfish poisoning toxins. J. Food Prot. 2002, 65, 1304–1308. [Google Scholar] [CrossRef]
- Rawn, D.F.K.; Niedzwiadek, B.; Campbell, K.; Higgins, H.C.; Elliott, C.T. Evaluation of surface plasmon resonance relative to high pressure liquid chromatography for the determination of paralytic shellfish toxins. J. Agric. Food Chem. 2009, 57, 10022–10031. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Sakai, T.; Doi, H.; Yamaguchi, K.; Yamada, A.; Takatani, T.; Arakawa, O. Tetrodotoxin/Saxitoxin Accumulation Profile in the Euryhaline Marine Pufferfish Chelonodontops patoca. Toxins 2024, 16, 18. https://doi.org/10.3390/toxins16010018
Zhu H, Sakai T, Doi H, Yamaguchi K, Yamada A, Takatani T, Arakawa O. Tetrodotoxin/Saxitoxin Accumulation Profile in the Euryhaline Marine Pufferfish Chelonodontops patoca. Toxins. 2024; 16(1):18. https://doi.org/10.3390/toxins16010018
Chicago/Turabian StyleZhu, Hongchen, Takashi Sakai, Hiroyuki Doi, Kenichi Yamaguchi, Akinori Yamada, Tomohiro Takatani, and Osamu Arakawa. 2024. "Tetrodotoxin/Saxitoxin Accumulation Profile in the Euryhaline Marine Pufferfish Chelonodontops patoca" Toxins 16, no. 1: 18. https://doi.org/10.3390/toxins16010018
APA StyleZhu, H., Sakai, T., Doi, H., Yamaguchi, K., Yamada, A., Takatani, T., & Arakawa, O. (2024). Tetrodotoxin/Saxitoxin Accumulation Profile in the Euryhaline Marine Pufferfish Chelonodontops patoca. Toxins, 16(1), 18. https://doi.org/10.3390/toxins16010018