Sodium Propionate Relieves LPS-Induced Inflammation by Suppressing the NF-ĸB and MAPK Signaling Pathways in Rumen Epithelial Cells of Holstein Cows

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Culture and Identification of Rumen Epithelial Cells
2.2. Effects of LPS, SP and PDTC on Cell Viability
2.3. Establishment of LPS-Induced Inflammatory Rumen Epithelial Cells Models In Vitro
2.4. Changes of Cytokines in Rumen Epithelial Cell Inflammation Models
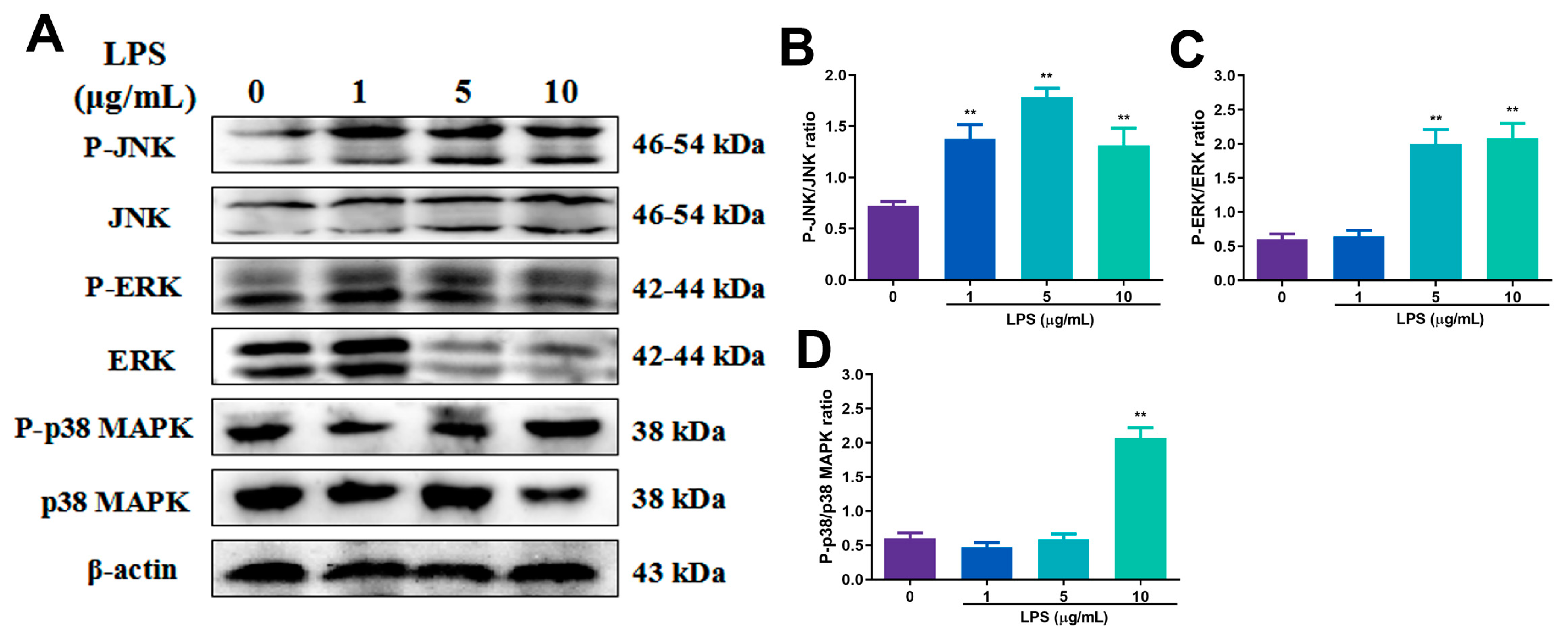
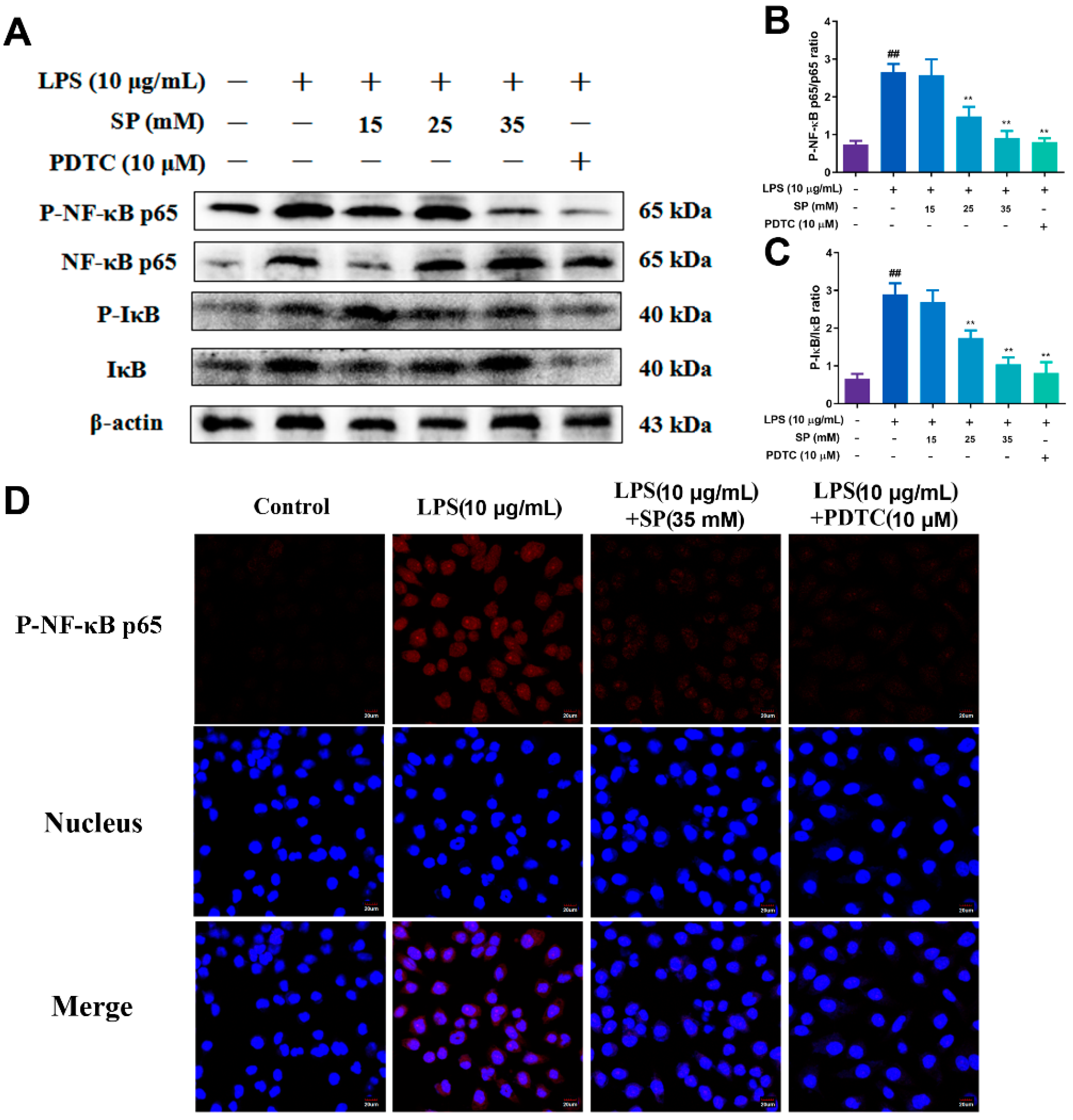
2.5. Effects of Different Concentrations of SP on LPS-Induced NF-κB and MAPK Signaling Pathway Proteins
2.6. Effects of SP on LPS-induced Cytokine Release and mRNA Expression
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Tissue Collection
5.2. Isolation, Culture and Identification of Primary Rumen Epithelial Cells (RECs)
5.3. Cell Viability Assay
5.4. Cell Treatments
5.5. Protein Extraction and Western Blotting (WB) Assay
5.6. Immunofluorescence
5.7. Enzyme-Linked Immunosorbent Assay (ELISA)
5.8. Quantitative Real-Time PCR
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abaker, J.A.; Xu, T.L.; Jin, D.; Chang, G.J.; Zhang, K.; Shen, X.Z. Lipopolysaccharide derived from the digestive tract provokes oxidative stress in the liver of dairy cows fed a high-grain diet. J. Dairy Sci. 2017, 100, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Su, X.; Li, J.; Yang, Y.; Wang, P.; Yan, F.; Yao, J.; Wu, S. Real-time monitoring of ruminal microbiota reveals their roles in dairy goats during subacute ruminal acidosis. NPJ Biofilms Microbi. 2021, 7, 45. [Google Scholar] [CrossRef]
- Castillo-Lopez, E.; Rivera-Chacon, R.; Ricci, S.; Petri, R.M.; Reisinger, N.; Zebeli, Q. Short-term screening of multiple phytogenic compounds for their potential to modulate chewing behavior, ruminal fermentation profile, and pH in cattle fed grain-rich diets. J. Dairy Sci. 2021, 104, 4271–4289. [Google Scholar] [CrossRef]
- Enemark, J.M. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. Vet. J. 2008, 176, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Mulligan, F.J.; Neville, E.W.; Guan, L.L.; Steele, M.A.; Penner, G.B. Invited review: Effect of subacute ruminal acidosis on gut health of dairy cows. J. Dairy Sci. 2022, 105, 7141–7160. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.G.; Lin, X.Y.; Yan, Z.G.; Hu, Z.Y.; Liu, G.M.; Sun, Y.D.; Liu, X.W.; Wang, Z.H. Effect of dietary roughage level on chewing activity, ruminal pH, and saliva secretion in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2660–2671. [Google Scholar] [CrossRef]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Li, X.; Guan, Y.; Wang, Y.; Yuan, X.; Sun, G.; Wang, Z.; Li, X. Inflammatory mechanism of Rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef]
- Petri, R.M.; Aditya, S.; Humer, E.; Zebeli, Q. Effect of an intramammary lipopolysaccharide challenge on the hindgut microbial composition and fermentation of dairy cattle experiencing intermittent subacute ruminal acidosis. J. Dairy Sci. 2021, 104, 5417–5431. [Google Scholar] [CrossRef]
- Nyati, K.K.; Masuda, K.; Zaman, M.M.; Dubey, P.K.; Millrine, D.; Chalise, J.P.; Higa, M.; Li, S.; Standley, D.M.; Saito, K.; et al. TLR4-induced NF-kappaB and MAPK signaling regulate the IL-6 mRNA stabilizing protein Arid5a. Nucleic Acids Res. 2017, 45, 2687–2703. [Google Scholar] [CrossRef]
- Hoogland, I.C.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic inflammation and microglial activation: Systematic review of animal experiments. J. Neuroinflamm. 2015, 12, 114. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Zebeli, Q.; Patra, A.K.; Greco, G.; Amasheh, S.; Penner, G.B. Symposium review: The importance of the ruminal epithelial barrier for a healthy and productive cow. J. Dairy Sci. 2019, 102, 1866–1882. [Google Scholar] [CrossRef]
- Zebeli, Q.; Ghareeb, K.; Humer, E.; Metzler-Zebeli, B.U.; Besenfelder, U. Nutrition, rumen health and inflammation in the transition period and their role on overall health and fertility in dairy cows. Res. Vet. Sci. 2015, 103, 126–136. [Google Scholar] [CrossRef]
- Zebeli, Q.; Metzler-Zebeli, B.U. Interplay between rumen digestive disorders and diet-induced inflammation in dairy cattle. Res. Vet. Sci. 2012, 93, 1099–1108. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Al-Lahham, S.; Rezaee, F. Propionic acid counteracts the inflammation of human subcutaneous adipose tissue: A new avenue for drug development. Daru 2019, 27, 645–652. [Google Scholar] [CrossRef]
- Wang, K.; Nan, X.; Tong, J.; Zhao, G.; Jiang, L.; Xiong, B. Steam Explosion Pretreatment Changes Ruminal Fermentation in vitro of Corn Stover by Shifting Archaeal and Bacterial Community Structure. Front. Microbiol. 2020, 11, 2027. [Google Scholar] [CrossRef]
- Otaru, S.M.; Adamu, A.M.; Ehoche, O.W. Influence of levels of supplementary concentrate mixture on lactation performance of Red Sokoto does and the pre-weaning growth rate of their kids. Vet. Anim. Sci. 2020, 10, 100137. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Loh, T.C.; Samsudin, A.A.; Foo, H.L.; Humam, A.M.; Shazali, N. Effects of postbiotic supplementation on growth performance, ruminal fermentation and microbial profile, blood metabolite and GHR, IGF-1 and MCT-1 gene expression in post-weaning lambs. BMC Vet. Res. 2019, 15, 315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Nan, X.; Wang, H.; Guo, Y.; Xiong, B. Research on the Applications of Calcium Propionate in Dairy Cows: A Review. Animals 2020, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Tamate, H.; McGilliard, A.D.; Jacobson, N.L.; Getty, R. Effect of Various Dietaries on the Anatomical Development of the Stomach in the Calf1. J. Dairy Sci. 1962, 45, 408–420. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 15876. [Google Scholar] [CrossRef] [PubMed]
- Chalermwatanachai, T.; Vilchez-Vargas, R.; Holtappels, G.; Lacoere, T.; Jáuregui, R.; Kerckhof, F.M.; Pieper, D.H.; Van de Wiele, T.; Vaneechoutte, M.; Van Zele, T.; et al. Chronic rhinosinusitis with nasal polyps is characterized by dysbacteriosis of the nasal microbiota. Sci. Rep-UK 2018, 8, 7926. [Google Scholar] [CrossRef]
- Wang, J.; Wei, Z.; Zhang, X.; Wang, Y.; Yang, Z.; Fu, Y. Propionate Protects against Lipopolysaccharide-Induced Mastitis in Mice by Restoring Blood-Milk Barrier Disruption and Suppressing Inflammatory Response. Front. Immunol. 2017, 8, 1108. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, Y.; Wang, P.; Huang, Y.; Wang, F. Short-Chain Fatty Acids Manifest Stimulative and Protective Effects on Intestinal Barrier Function Through the Inhibition of NLRP3 Inflammasome and Autophagy. Cell Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-chain fatty acids suppress lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines through inhibition of NF-κB pathway in RAW264.7 cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef]
- Garrett, E.F.; Nordlund, K.V.; Goodger, W.J.; Oetzel, G.R. cross-sectional field study investigating the effect of periparturient dietary management on ruminal pH in early lactation dairy cows. J. Dairy Sci. 1997, 80, 169. [Google Scholar]
- Oetzel, G.R.; Nordlund, K.V.; Garrett, E.F. Effect of ruminal pH and stage of lactation on ruminal lactate concentrations in dairy cows. J. Dairy Sci. 1999, 82, 38. [Google Scholar]
- Wang, M.Y.; Li, Y.; Gao, M.; Song, L.W.; Xu, M.; Zhao, X.L.; Jia, Y.; Zhao, M.; Sun, Y.Y.; Hu, H.L. Effects of subacute ruminal acidosis on colon epithelial morphological structure, permeability, and expression of key tight junction proteins in dairy goats. J. Dairy Sci. 2021, 104, 4260–4270. [Google Scholar] [CrossRef]
- Humer, E.; Petri, R.M.; Aschenbach, J.R.; Bradford, B.J.; Penner, G.B.; Tafaj, M.; Sudekum, K.H.; Zebeli, Q. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 2018, 101, 872–888. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Garrett, E.F.; Pereira, M.N.; Nordlund, K.V.; Armentano, L.E.; Goodger, W.J.; Oetzel, G.R. Diagnostic methods for the detection of subacute ruminal acidosis in dairy cows. J. Dairy Sci. 1999, 82, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, K.V.; Garrett, E.F.; Oetzel, G.R. Herd-based rumenocentesis: A clinical approach to the diagnosis of subacute rumen acidosis. Compend. Contin. Educ. Pract. Vet. 1995, 17, 48–56. [Google Scholar]
- Humer, E.; Aschenbach, J.R.; Neubauer, V.; Kroger, I.; Khiaosa-Ard, R.; Baumgartner, W.; Zebeli, Q. Signals for identifying cows at risk of subacute ruminal acidosis in dairy veterinary practice. J. Anim. Physiol. An. N. 2018, 102, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Tech. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Abdela, N. Sub-acute Ruminal Acidosis (SARA) and its Consequence in Dairy Cattle: A Review of Past and Recent Research at Global Prospective. Achiev. Life Sci. 2016, 10, 187–196. [Google Scholar] [CrossRef]
- Ma, J.; Shah, A.M.; Wang, Z.; Fan, X. Potential protective effects of thiamine supplementation on the ruminal epithelium damage during subacute ruminal acidosis. Anim. Sci. J. 2021, 92, e13579. [Google Scholar] [CrossRef]
- Gozho, G.N.; Krause, D.O.; Plaizier, J.C. Rumen lipopolysaccharide and inflammation during grain adaptation and subacute ruminal acidosis in steers. J. Dairy Sci. 2006, 89, 4404–4413. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Khafipour, E.; Li, S.; Gozho, G.N.; Krause, D.O. Subacute ruminal acidosis (SARA), endotoxins and health consequences. Anim. Feed Sci. Tech. 2012, 172, 9–21. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Danesh Mesgaran, M.; Derakhshani, H.; Golder, H.; Khafipour, E.; Kleen, J.L.; Lean, I.; Loor, J.; Penner, G.; Zebeli, Q. Review: Enhancing gastrointestinal health in dairy cows. Animal 2018, 12, s399–s418. [Google Scholar] [CrossRef] [PubMed]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. Int. J. Mol. Sci. 2020, 21, 3026. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lei, Q.; Ma, H.; Jiang, M.; Yang, T.; Ma, Q.; Datsomor, O.; Zhan, K.; Zhao, G. Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury. Toxins 2022, 14, 337. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Teshima, S.; Kuwano, Y.; Oka, A.; Kishi, K.; Rokutan, K. Helicobacter pylori lipopolysaccharide induces apoptosis of cultured guinea pig gastric mucosal cells. Am. J. Physiol. Gastr. L. 2001, 281, G726–G734. [Google Scholar] [CrossRef]
- Xie, M.Y.; Hou, L.J.; Sun, J.J.; Zeng, B.; Xi, Q.Y.; Luo, J.Y.; Chen, T.; Zhang, Y.L. Porcine Milk Exosome MiRNAs Attenuate LPS-Induced Apoptosis through Inhibiting TLR4/NF-kappaB and p53 Pathways in Intestinal Epithelial Cells. J. Agric. Food Chem. 2019, 67, 9477–9491. [Google Scholar] [CrossRef]
- Hu, Z.; Lin, M.; Ma, X.; Zhao, G.; Zhan, K. Effect of Tea Tree Oil on the Expression of Genes Involved in the Innate Immune System in Goat Rumen Epithelial Cells. Animals 2021, 11, 2460. [Google Scholar] [CrossRef]
- Kang, Y.; Su, G.; Sun, J.; Zhang, Y. Activation of the TLR4/MyD88 signaling pathway contributes to the development of human hepatocellular carcinoma via upregulation of IL-23 and IL-17A. Oncol. Lett. 2018, 15, 9647–9654. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Bartley, E.E.; Fina, L.R.; Anthony, H.D. Relationship of rumen gram-negative bacteria and free endotoxin to lactic acidosis in cattle. J. Anim. Sci. 1978, 47, 1329–1337. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, M.; Chen, C.; Liu, L.; Wei, X.; Zeng, S. Toll-Like Receptor 4 (TLR4)/Opioid Receptor Pathway Crosstalk and Impact on Opioid Analgesia, Immune Function, and Gastrointestinal Motility. Front. Immunol. 2020, 11, 1455. [Google Scholar] [CrossRef]
- Kellum, J.A.; Kong, L.; Fink, M.P.; Weissfeld, L.A.; Yealy, D.M.; Pinsky, M.R.; Fine, J.; Krichevsky, A.; Delude, R.L.; Angus, D.C. Understanding the inflammatory cytokine response in pneumonia and sepsis: Results of the Genetic and Inflammatory Markers of Sepsis (GenIMS) Study. Arch. Intern. Med. 2007, 167, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhu, W.; Mao, S. High-concentrate feeding upregulates the expression of inflammation-related genes in the ruminal epithelium of dairy cattle. J. Anim. Sci. Biotechnol. 2016, 7, 42. [Google Scholar] [CrossRef]
- Harding, A.; Cortez-Toledo, E.; Magner, N.L.; Beegle, J.R.; Coleal-Bergum, D.P.; Hao, D.; Wang, A.; Nolta, J.A.; Zhou, P. Highly Efficient Differentiation of Endothelial Cells from Pluripotent Stem Cells Requires the MAPK and the PI3K Pathways. Stem Cells (Dayton, Ohio) 2017, 35, 909–919. [Google Scholar] [CrossRef]
- El-Gamal, M.I.; Anbar, H.S.; Tarazi, H.; Oh, C.H. Discovery of a potent p38α/MAPK14 kinase inhibitor: Synthesis, in vitro/in vivo biological evaluation, and docking studies. Eur. J. Med. Chem. 2019, 183, 111684. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Elmhadi, M.; Wang, C.; Li, Z.; Zhang, H.; He, B.; Zhao, X.; Zhang, Z.; Wang, H. Thiamine Supplementation Alleviates Lipopolysaccharide-Triggered Adaptive Inflammatory Response and Modulates Energy State via Suppression of NFkappaB/p38 MAPK/AMPK Signaling in Rumen Epithelial Cells of Goats. Antioxidants 2022, 11, 2048. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xiong, B.; Zhao, X. Could propionate formation be used to reduce enteric methane emission in ruminants? Sci. Total. Environ. 2023, 855, 158867. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Gao, W.; Fang, Z.; Lei, L.; Ju, L.; Jin, B.; Loor, J.J.; Liang, Y.; Shi, Z.; Shen, T.; Yu, H.; et al. Propionate alleviates palmitic acid-induced endoplasmic reticulum stress by enhancing autophagy in calf hepatic cells. J. Dairy Sci. 2021, 104, 9316–9326. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Tong, L.C.; Wang, Y.; Wang, Z.B.; Liu, W.Y.; Sun, S.; Li, L.; Su, D.F.; Zhang, L.C. Propionate Ameliorates Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation and Oxidative Stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef]
- Kara, Ç.; Orman, A.; Udum, D.; Yavuz, H.M.; Kovanlıkaya, A. Effects of calcium propionate by different numbers of applications in first week postpartum of dairy cows on hypocalcemia, milk production and reproductive disorders. Ital. J. Anim. Sci 2016, 8, 259–270. [Google Scholar] [CrossRef]
- Huang, W.; Man, Y.; Gao, C.; Zhou, L.; Gu, J.; Xu, H.; Wan, Q.; Long, Y.; Chai, L.; Xu, Y.; et al. Short-Chain Fatty Acids Ameliorate Diabetic Nephropathy via GPR43-Mediated Inhibition of Oxidative Stress and NF-κB Signaling. Oxid. Med. Cell. Longevity 2020, 2020, 4074832. [Google Scholar] [CrossRef]
- Moylan, H.E.C.; Nguyen-Ngo, C.; Lim, R.; Lappas, M. The short-chain fatty acids butyrate and propionate protect against inflammation-induced activation of mediators involved in active labor: Implications for preterm birth. Mol. Hum. Reprod. 2020, 26, 452–468. [Google Scholar] [CrossRef]
- Park, H.S.; Han, J.H.; Park, J.W.; Lee, D.H.; Jang, K.W.; Lee, M.; Heo, K.S.; Myung, C.S. Sodium propionate exerts anticancer effect in mice bearing breast cancer cell xenograft by regulating JAK2/STAT3/ROS/p38 MAPK signaling. Acta Pharmacol. Sin. 2021, 42, 1311–1323. [Google Scholar] [CrossRef]
- Shi, M.; Deng, Y.; Yu, H.; Xu, L.; Shi, C.; Chen, J.; Li, G.; Du, Y.; Wang, Y.G. Protective Effects of Oridonin on Acute Liver Injury via Impeding Posttranslational Modifications of Interleukin-1 Receptor-Associated Kinase 4 (IRAK4) in the Toll-Like Receptor 4 (TLR4) Signaling Pathway. Mediat. Inflamm. 2019, 2019, 7634761. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Yang, M.; Wang, Y.; Yang, C.; Shafiq, M.; Wang, G.; Li, L. Sodium propionate protect the blood-milk barrier integrity, relieve lipopolysaccharide-induced inflammatory injury and cells apoptosis. Life Sci. 2021, 270, 119138. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.; Henricks, P.A.J.; Garssen, J.; Folkerts, G. Time and Concentration Dependent Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor α-Induced Endothelial Activation. Front. Pharmacol. 2018, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90 (Suppl. S1), E17–E38. [Google Scholar] [CrossRef]
- Zhao, C.; Bobe, G.; Wang, Y.; Zhang, X.; Zhao, Z.; Zhang, S.; Sun, G.; Yuan, X.; Li, X.; Liu, G. Potential Role of SLC5A8 Expression in the Etiology of Subacute Ruminal Acidosis. Front. Vet. Sci. 2020, 7, 394. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, Y.; Yuan, X.; Sun, G.; Shen, B.; Xu, F.; Fan, G.; Jin, M.; Li, X.; Liu, G. Berberine inhibits lipopolysaccharide-induced expression of inflammatory cytokines by suppressing TLR4-mediated NF-kB and MAPK signaling pathways in rumen epithelial cells of Holstein calves. J. Dairy Res. 2019, 86, 171–176. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Tm (°C) | Length |
|---|---|---|---|
| TNF-α | For CTGCCGGACTACCTGGACTAT Rev CCTCACTTCCCTACATCCCTAA | 60.75 58.35 | 234 bp |
| IL-6 | For AACGAGTGGGTAAAGAACGC Rev CTGACCAGAGGAGGGAATGC | 58.49 59.82 | 140 bp |
| IL-1β | For CTGAACCCATCAACGAAA Rev ATGACCGACACCACCTGC | 52.48 59.65 | 190 bp |
| β-actin | For GCCCTGAGGCTCTCTTCCA Rev GCGGATGTCGACGTCACA | 60.99 60.13 | 101 bp |
| GAPDH | For CCTGCCAAGTATGATGAGAT Rev AGTGTCGCTGTTGAAGTC | 58.53 59.75 | 117 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Yi, F.; Wei, B.; Tan, P.; Huang, Y.; Zeng, F.; Wang, Y.; Xu, C.; Wang, J. Sodium Propionate Relieves LPS-Induced Inflammation by Suppressing the NF-ĸB and MAPK Signaling Pathways in Rumen Epithelial Cells of Holstein Cows. Toxins 2023, 15, 438. https://doi.org/10.3390/toxins15070438
Zhao C, Yi F, Wei B, Tan P, Huang Y, Zeng F, Wang Y, Xu C, Wang J. Sodium Propionate Relieves LPS-Induced Inflammation by Suppressing the NF-ĸB and MAPK Signaling Pathways in Rumen Epithelial Cells of Holstein Cows. Toxins. 2023; 15(7):438. https://doi.org/10.3390/toxins15070438
Chicago/Turabian StyleZhao, Chenxu, Fanxuan Yi, Bo Wei, Panpan Tan, Yan Huang, Fangyuan Zeng, Yazhou Wang, Chuang Xu, and Jianguo Wang. 2023. "Sodium Propionate Relieves LPS-Induced Inflammation by Suppressing the NF-ĸB and MAPK Signaling Pathways in Rumen Epithelial Cells of Holstein Cows" Toxins 15, no. 7: 438. https://doi.org/10.3390/toxins15070438
APA StyleZhao, C., Yi, F., Wei, B., Tan, P., Huang, Y., Zeng, F., Wang, Y., Xu, C., & Wang, J. (2023). Sodium Propionate Relieves LPS-Induced Inflammation by Suppressing the NF-ĸB and MAPK Signaling Pathways in Rumen Epithelial Cells of Holstein Cows. Toxins, 15(7), 438. https://doi.org/10.3390/toxins15070438