Testing the Novel Weapons Hypothesis of the Argentine Ant Venom on Amphibians

 , , and
, , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
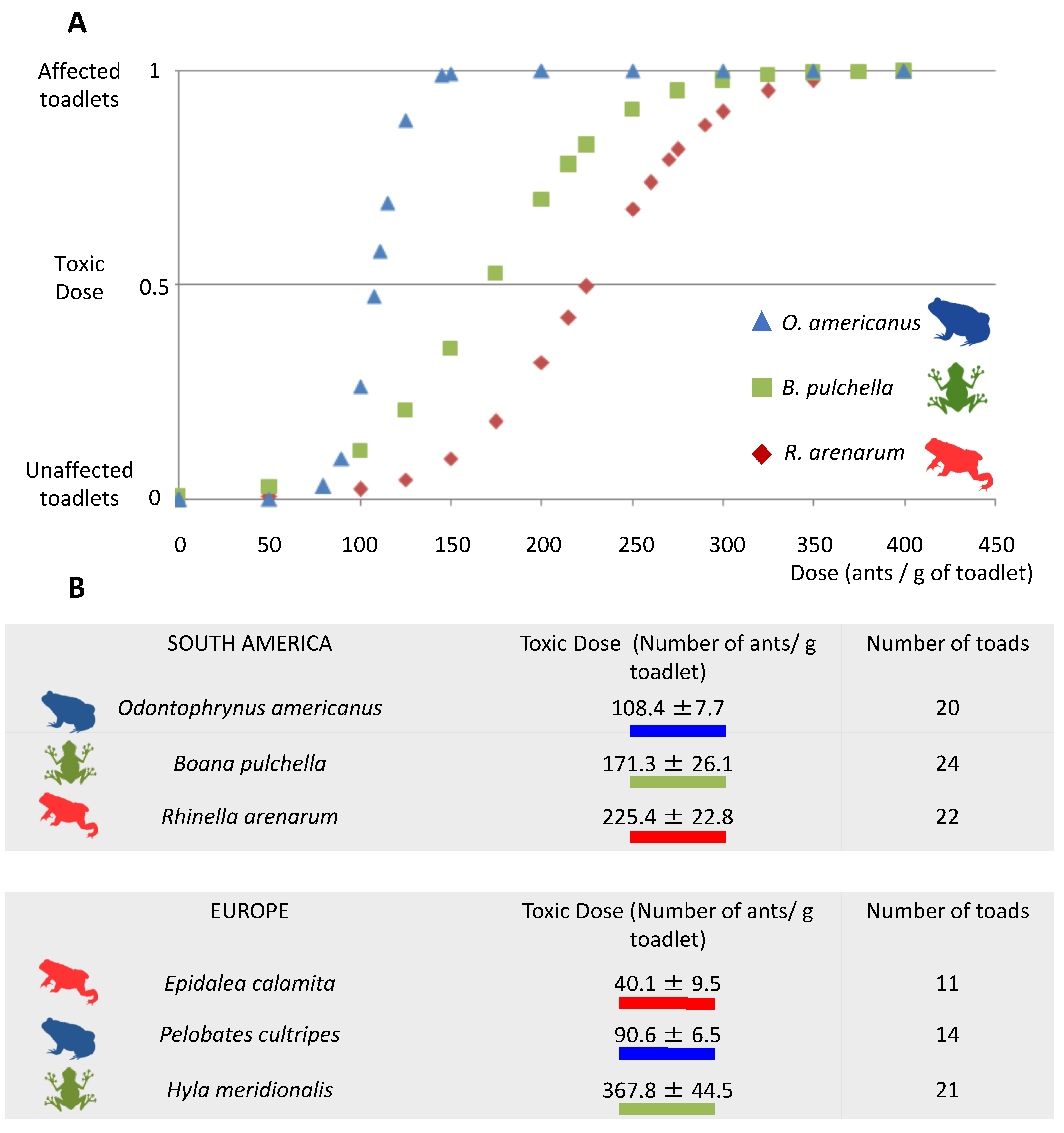
2.1. Toxic Doses to the Argentine Ant Venom
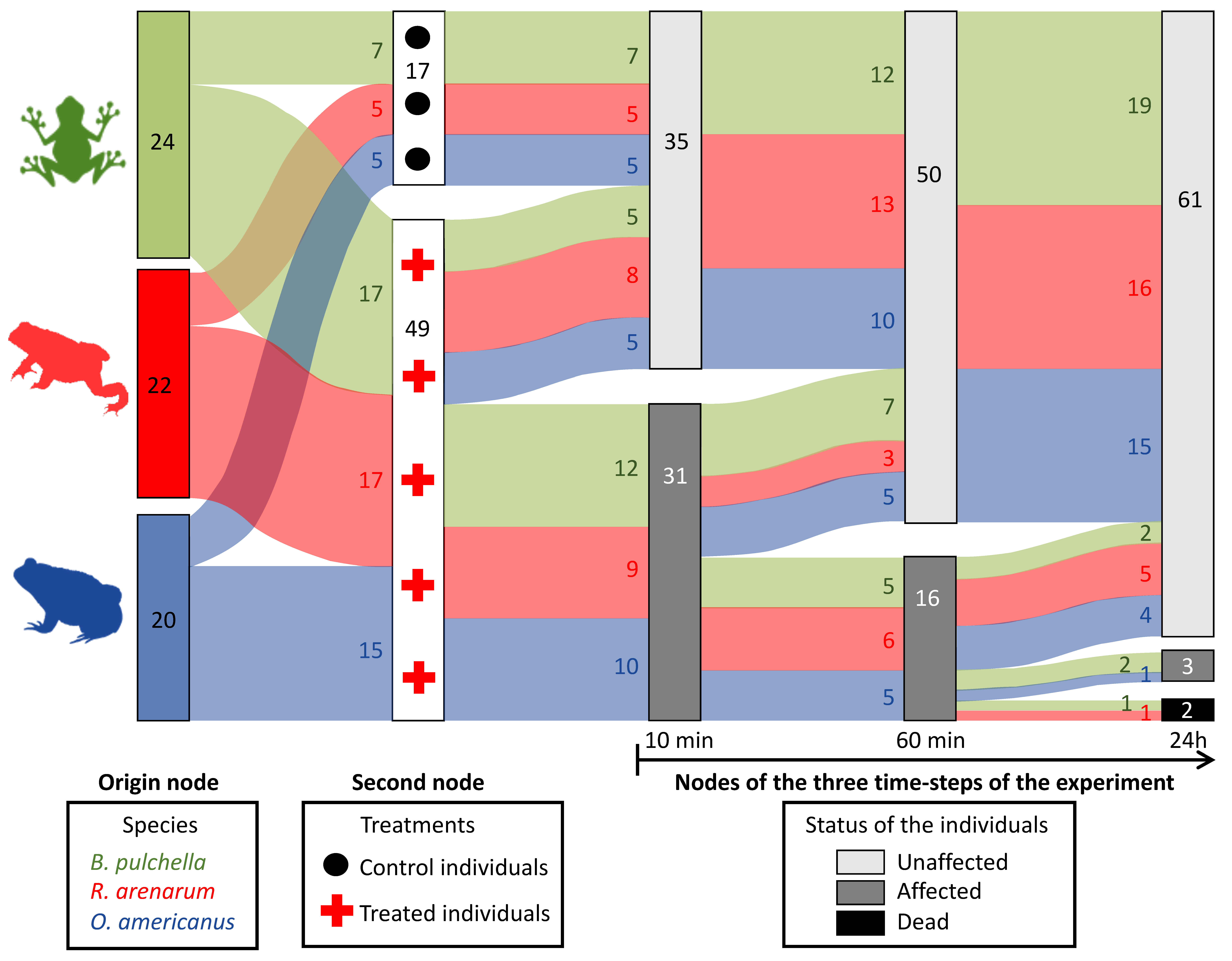
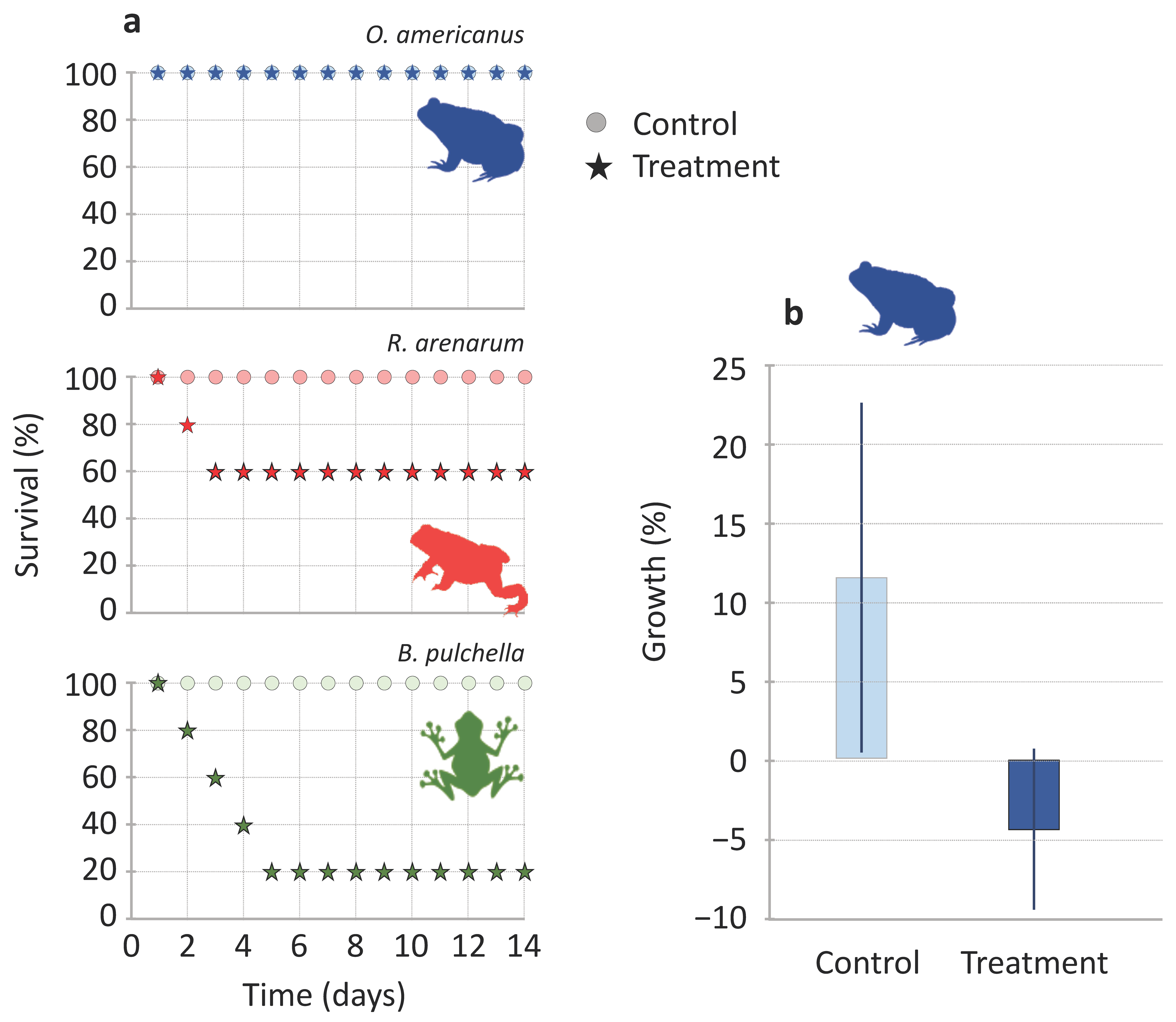
2.2. Short-Term Recovery after Venom Application
2.3. Medium-Term Effects of the Argentine Ant Venom
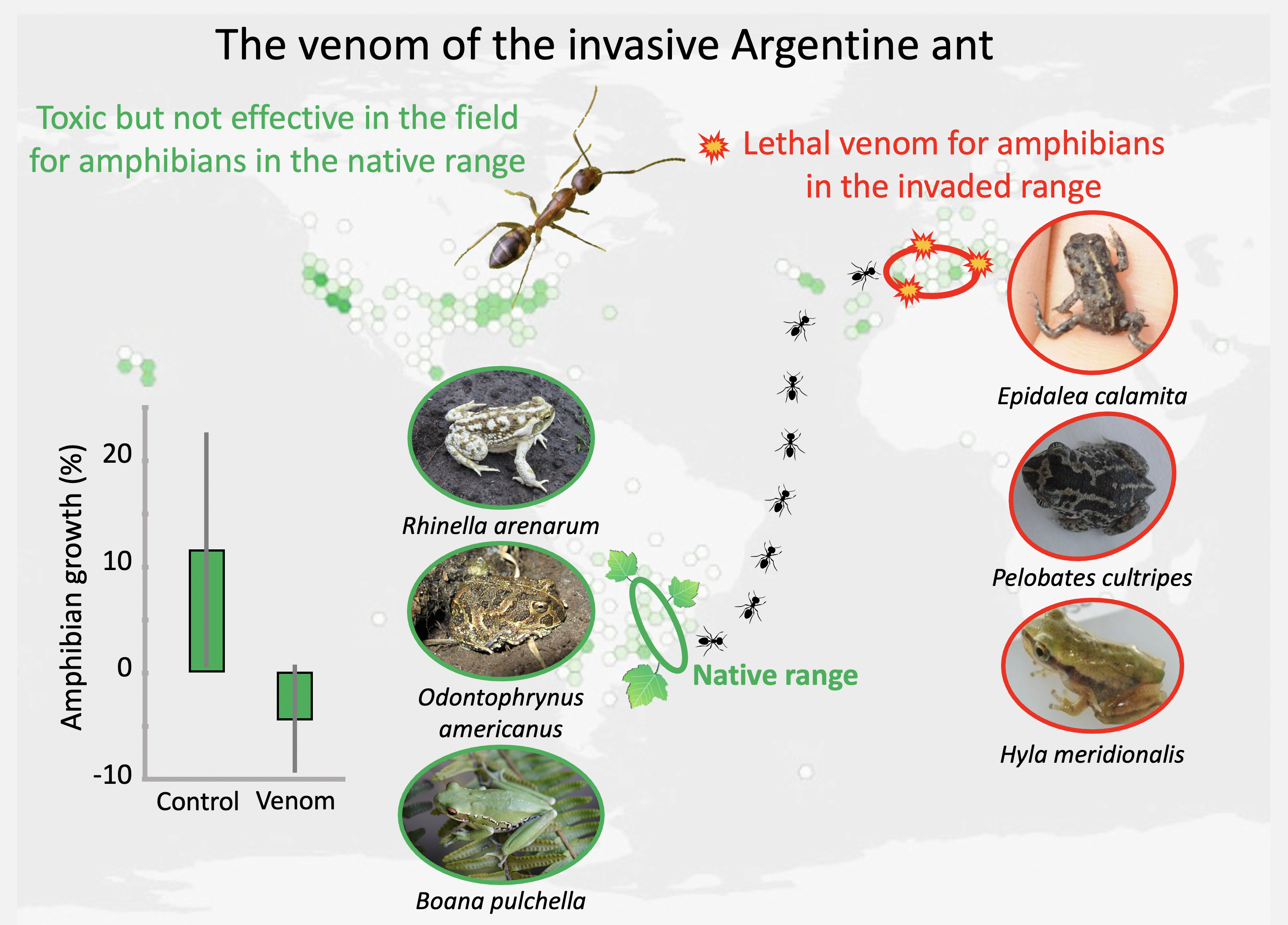
3. Discussion
3.1. Toxicity to Amphibians in the Argentine Ant’s Native Range
3.2. The Importance of Ant Community in Exposure to the Venom
4. Conclusions
5. Materials and Methods
5.1. Collection of Species and Housing
5.2. Dose-Response Experiment
5.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M.; Pergl, J.; Jarošik, V.; Sixtova, Z.; Weber, E. Geographical and taxonomic biases in invasion ecology. Trends Ecol. Evol. 2008, 23, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Kumschick, S.; Gaertner, M.; Vilà, M.; Essl, F.; Jeschke, J.M.; Pyšek, P.; Ricciardi, A.; Bacher, S.; Blackburn, T.M.; Dick, J.T.; et al. Ecological impacts of alien species: Quantification, scope, caveats, and recommendations. Bioscience 2015, 65, 55–63. [Google Scholar] [CrossRef]
- Bellard, C.; Jeschke, J.M. A spatial mismatch between invader impacts and research publications. Conserv. Biol. 2016, 30, 230–232. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M.; et al. Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; Garcia-Berthou, E.; Pascal, M. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG): Auckland, New Zealand, 2000. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Suarez, A.V.; Holway, D.A.; Tsutsui, N.D. Genetics and behavior of a colonizing species: The invasive Argentine ant. Am. Nat. 2008, 172, S72–S84. [Google Scholar] [CrossRef] [PubMed]
- Passera, L. Characteristics of tramp species. In Exotic Ants: Biology, Impact and Control of Introduced Species; Williams, D.F., Ed.; Westview Press: Boulder, CO, USA, 1997; pp. 23–43. [Google Scholar]
- Holway, D.A.; Lach, L.; Suarez, A.V.; Tsutsui, N.D.; Case, T.J. The causes and consequences of ant invasions. Annu. Rev. Ecol. Syst. 2002, 33, 181–233. [Google Scholar] [CrossRef]
- Welzel, K.F.; Lee, S.H.; Dossey, A.T.; Chauhan, K.R.; Choe, D.-H. Verification of Argentine ant defensive compounds and their behavioral effects on heterospecific competitors and conspecific nestmates. Sci. Rep. 2018, 8, 1477. [Google Scholar] [CrossRef]
- Alvarez-Blanco, P.; Cerdá, X.; Hefetz, A.; Boulay, R.; Bertó-Moran, A.; Díaz-Paniagua, C.; Lenoir, A.; Billen, J.; Liedtke, C.; Chauhan, K.R.; et al. Effects of the Argentine ant venom on terrestrial amphibians. Conserv. Biol. 2021, 35, 216–226. [Google Scholar] [CrossRef]
- Suarez, A.V.; Case, T.J. Bottom-up effects on persistence of a specialist predator: Ant invasions and horned lizards. Ecol. Appl. 2002, 12, 291–298. [Google Scholar] [CrossRef]
- Touyama, Y.; Ilhara, Y.; Ito, F. Argentine ant infestation affects the abundance of the native myrmecophagic jumping spider Siler cupreus Simon in Japan. Insectes Sociaux 2008, 55, 144–146. [Google Scholar] [CrossRef]
- Alvarez-Blanco, P.; Caut, S.; Cerdá, X.; Angulo, E. Native predators living in invaded areas: Responses of terrestrial amphibian species to an Argentine ant invasion. Oecologia 2017, 185, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Blanco, P.; Broggi, J.; Cerdá, X.; González-Jarri, O.; Angulo, E. Breeding consequences for a songbird nesting in Argentine ant’ invaded land. Biol. Invasions 2020, 22, 2883–2898. [Google Scholar] [CrossRef]
- Pianka, E.R.; Parker, W.S. Ecology of horned lizard: A review with special reference to Phrynosoma platyrhinos. Copeia 1975, 1975, 141–162. [Google Scholar] [CrossRef]
- Gotelli, N.J. Ant community structure: Effects of predatory ant lions. Ecology 1996, 77, 630–638. [Google Scholar] [CrossRef]
- Naples, V.L. Morphology, evolution and function offeeding in the giant anteater (Myrmecophaga tridactyla). J. Zool. 1999, 249, 19–41. [Google Scholar] [CrossRef]
- Berazategui, M.; Camargo, A.; Maneyro, R. Enviromental and seasonal variation in the diet of Elachistocleis bicolor (Guérin-Méneville1838) (Anura: Mycrohylidae) from northern Uruguay. Zool. Sci. 2007, 24, 225–231. [Google Scholar] [CrossRef]
- Wiebe, K.L.; Gow, E.A. Choice of foraging habitat by northern flickers reflects changes in availability of their ant prey linked to ambient temperature. Ecoscience 2013, 20, 122–130. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2010. [Google Scholar]
- Caldwell, J.P. The evolution of myrmecophagy and its correlates in poison frogs (Family Dendrobatidae). J. Zool. 1996, 240, 75–101. [Google Scholar] [CrossRef]
- Mebs, D.; Jansen, M.; Köhler, G.; Pogoda, W.; Kauert, G. Myrmecophagy and alkaloid sequestration in amphibians: A study on Ameerega picta (Dendrobatidae) and Elachistocleis sp. (Microhylidae) Frogs. Salamandra 2010, 46, 11–15. [Google Scholar]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 7217–7233. [Google Scholar]
- Sih, A.; Bolnick, D.I.; Luttbeg, B.; Orrock, J.L.; Peacor, S.D.; Pintor, L.M.; Preisser, E.; Rehage, J.S.; Vonesh, J.R. Predator-prey naïveté, antipredator behavior, and the ecology of predator invasions. Oikos 2010, 119, 610–621. [Google Scholar]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 2009, 15, 22–40. [Google Scholar] [CrossRef]
- Quiroga, L.B.; Sanabria, E.A.; Acosta, J.C. Size-and sex-dependent variation in diet of Rhinella arenarum (Anura: Bufonidae) in a wetland of San Juan, Argentina. J. Herpetol. 2009, 42, 311–317. [Google Scholar] [CrossRef]
- Moreira, L. Ecologíatrófica de Anfibios Juveniles (Amphibia: Anura) y Análisis de la Incidencia de las Hormigas (Hymenoptera: Formicidae) Como Presa Potencial. Undergraduate Thesis, Universidad de la República, Montevideo, Uruguay, 2021. [Google Scholar]
- Isacch, J.P.; Barg, M. Are bufonid toads specialized ant-feeders? A case test from the Argentinian flooding pampa. J. Nat. Hist. 2002, 36, 2005–2012. [Google Scholar] [CrossRef]
- Achaval Coppes, F. Análisis Cualitativo y Cuantitativo de la Dieta de Odontophrynus Americanus (Duméril & Bibron, 1841)(Anura: Cycloramphidae) en una Población del Centro-sur del Departamento de Tacuarembó (Uruguay). Undergraduate Thesis, Facultad de Ciencias. Universidad de la República, Montevideo, Uruguay, 2011. [Google Scholar]
- Maneyro, R.; da Rosa, I. Temporal and spatial changes in the diet of Hyla pulchella (Anura, Hylidae) in southern Uruguay. Phyllomedusa 2004, 3, 101–113. [Google Scholar] [CrossRef]
- Da Rosa, I.; Canavero, A.; Maneyro, R.; Camargo, A. Trophic niche variation and individual specialization in Hypsiboas pulchellus (Duméril and Bibron, 1841) (Anura, Hylidae) from Uruguay. South Am. J. Herpetol. 2011, 6, 98–106. [Google Scholar] [CrossRef]
- Allaire, J.J.; Gandrud, C.; Russell, K.; Yetman, C.J. Networkd3: D3 JavaScript Network Graphsfrom R. R Package Version 0.4. 2017. Available online: https://CRAN.R-project.org/package=networkD3 (accessed on 7 March 2022).
- Bortolini, S.V.; Maneyro, R.; Achaval-Coppes, F.; Zanella, N. Diet of Melanophryniscus devincenzii (Anura: Bufonidae) from Parque Municipal de Sertão, Rio Grande doSul, Brazil. Herpetol. J. 2013, 23, 115–119. [Google Scholar]
- Pollo, F.E.; Grenat, P.R.; Otero, M.A.; Babini, S.; Salas, N.E.; Martino, A.L. Evaluation in situ of genotoxic and cytotoxic response in the diploid/polyploid complex Odontophrynus (Anura: Odontophrynidae) inhabiting agroecosystems. Chemosphere 2019, 216, 306–312. [Google Scholar] [PubMed]
- Calcaterra, L.; Cabrera, S.; Briano, J. Local co-occurrence of several highly invasive ants in their native range: Are they all ecologically dominant species? Insectes Sociaux 2016, 63, 407–419. [Google Scholar]
- Clark, N.J.; Olsson-Pons, S.; Ishtiaq, F.; Clegg, S.M. Specialist enemies, generalist weapons and the potential spread of exotic pathogens: Malaria parasites in a highly invasive bird. Int. J. Parasitol. 2015, 45, 891–899. [Google Scholar] [PubMed]
- Lopes, R.J.; Correia, J.; Batalha, H.; Cardoso, G.C. Haemosporidian parasites missed the boat during the introduction of common waxbills (Estrildaastrild) in Iberia. Parasitology 2018, 145, 1493–1498. [Google Scholar] [PubMed]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar]
- Suarez, A.V.; Tsutsui, N.D.; Holway, D.A.; Case, T.J. Behavioral and genetic differentiation between native and introduced populations of the Argentine ant. Biol. Invasions 1999, 1, 43–53. [Google Scholar] [CrossRef]
- Calcaterra, L.A.; Livore, J.; Delgado, A.; Briano, J.A. Ecological dominance of the red imported fire ant, Solenopsis invicta, in its native range. Oecologia 2008, 156, 411–421. [Google Scholar]
- Blight, O.; Josens, R.; Bertelsmeier, C.; Abril, S.; Boulay, R.; Cerdá, X. Differences in behavioural traits among native and introduced colonies of an invasive ant. Biol. Invasions 2017, 19, 1389–1398. [Google Scholar]
- Holway, D.A.; Suarez, A.V. Homogenization of ant communities in Mediterranean California: The effects of urbanization and invasion. Biol. Conserv. 2006, 127, 319–326. [Google Scholar]
- Angulo, E.; Caut, S.; Cerdá, X. Scavenging in Mediterranean ecosystems: Effect of the invasive Argentine ant. Biol. Invasions 2011, 13, 1183–1194. [Google Scholar]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.M.; Brockerhoff, E.G.; Ward, D.; Keller, L. Recurrent bridgehead effects accelerate global alien ant spread. Proc. Natl. Acad. Sci. USA 2018, 115, 5486–5491. [Google Scholar] [PubMed]
- McConnell, J.F.; McL Mathieson, A.; Schoenborn, B.P. The crystal structure of the monoterpene iridomyrmecin at −150 °C. Acta Crystallogr. 1964, 17, 472–477. [Google Scholar] [CrossRef]
- Salado-Ortega, I.; Alvarez-Blanco, P.; Boulay, R.; Abril, S.; Cerdá, X.; Angulo, E. Defensive compounds and the success of the Argentine ant invasion. Chemoecology, in press. [CrossRef]
- Pavan, M. Iridomyrmyecin as insecticide. IX IntCongrEmtomol Amst. 1952, 19511, 321–327. [Google Scholar]
- Kahn, C. The Merck Veterinary Manual, 9th ed.; Merck Hardcover: Philadelphia, PA, USA, 2005. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Core Team, R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 7 March 2022).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llopart, J.P.; Alvarez-Blanco, P.; Moreira-Demarco, L.; Bang, A.; Angulo, E.; Maneyro, R. Testing the Novel Weapons Hypothesis of the Argentine Ant Venom on Amphibians. Toxins 2023, 15, 235. https://doi.org/10.3390/toxins15040235
Llopart JP, Alvarez-Blanco P, Moreira-Demarco L, Bang A, Angulo E, Maneyro R. Testing the Novel Weapons Hypothesis of the Argentine Ant Venom on Amphibians. Toxins. 2023; 15(4):235. https://doi.org/10.3390/toxins15040235
Chicago/Turabian StyleLlopart, Juan Pablo, Paloma Alvarez-Blanco, Lucía Moreira-Demarco, Alok Bang, Elena Angulo, and Raúl Maneyro. 2023. "Testing the Novel Weapons Hypothesis of the Argentine Ant Venom on Amphibians" Toxins 15, no. 4: 235. https://doi.org/10.3390/toxins15040235
APA StyleLlopart, J. P., Alvarez-Blanco, P., Moreira-Demarco, L., Bang, A., Angulo, E., & Maneyro, R. (2023). Testing the Novel Weapons Hypothesis of the Argentine Ant Venom on Amphibians. Toxins, 15(4), 235. https://doi.org/10.3390/toxins15040235