Assessment of Agricultural Practices for Controlling Fusarium and Mycotoxins Contamination on Maize Grains: Exploratory Study in Maize Farms

 ,
,  , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
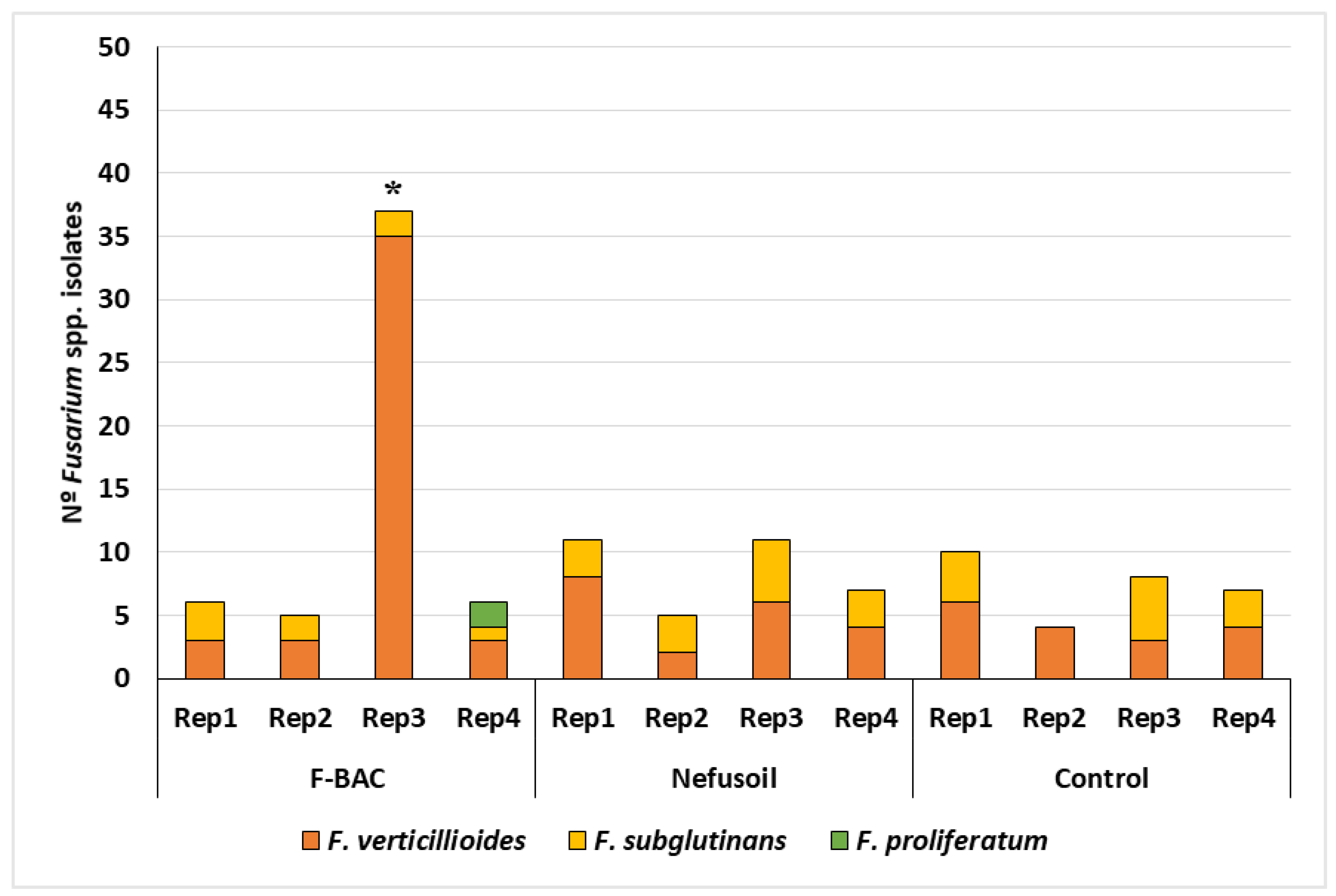
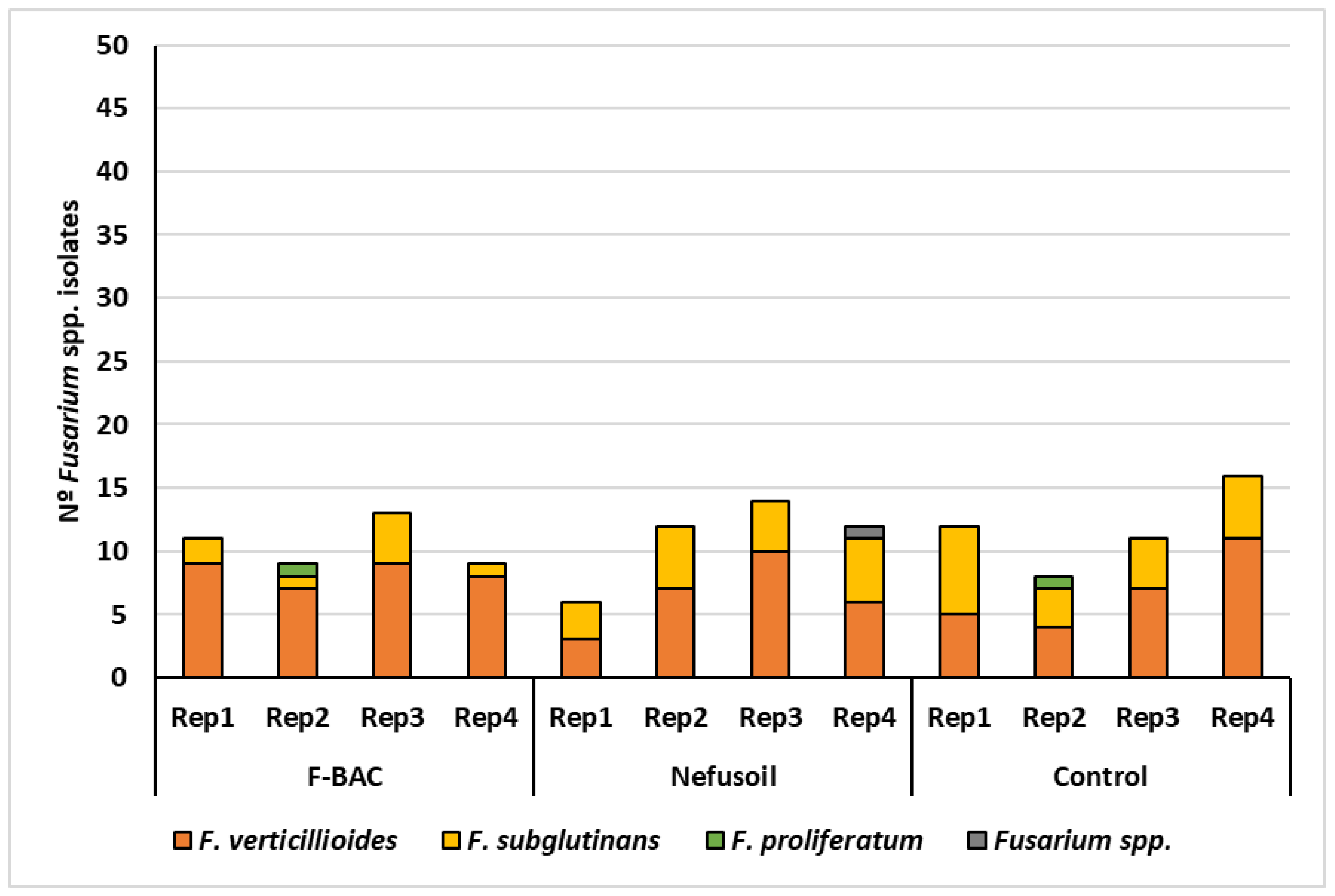
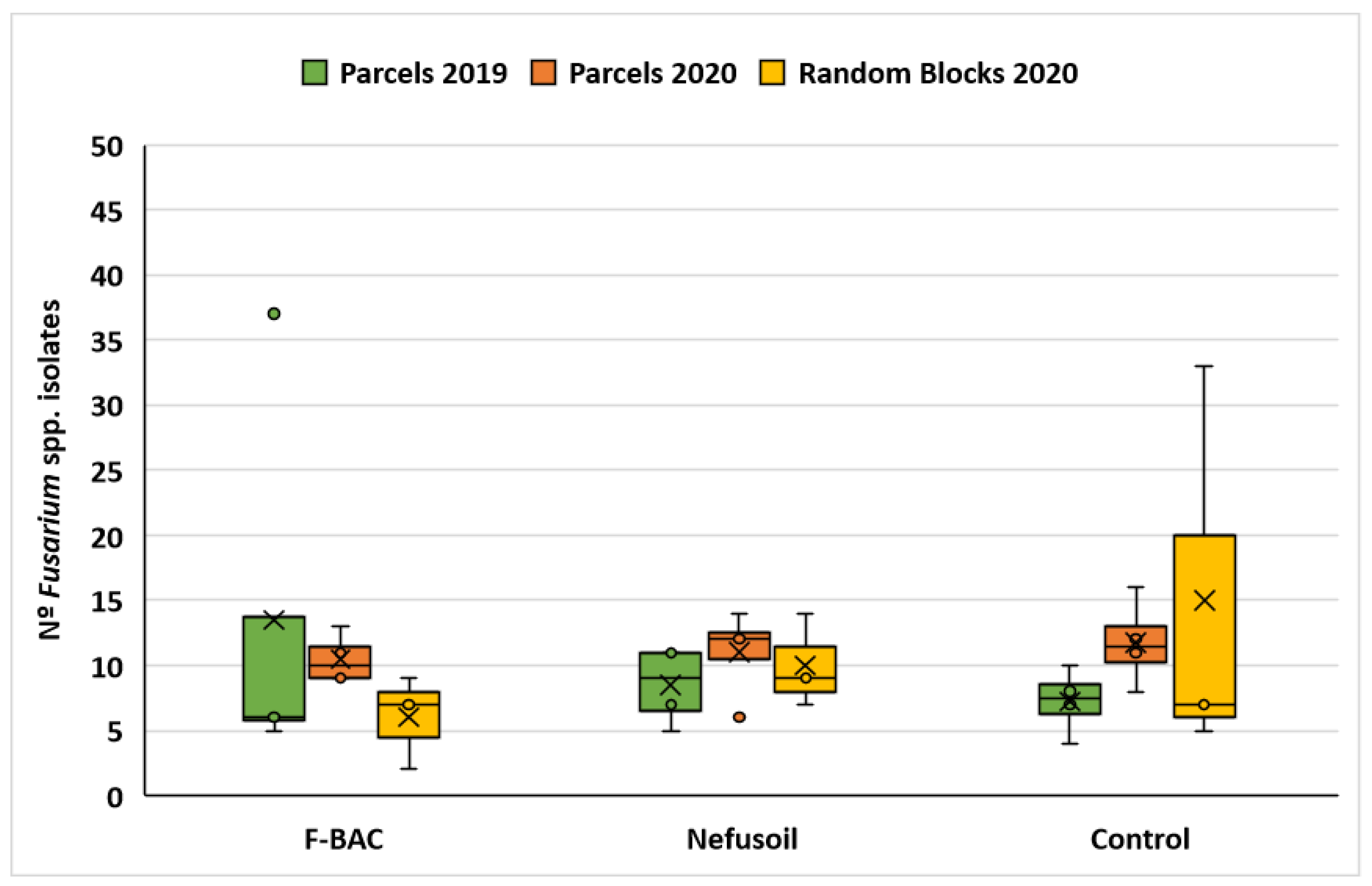
2.1. Fungal Contamination of Maize
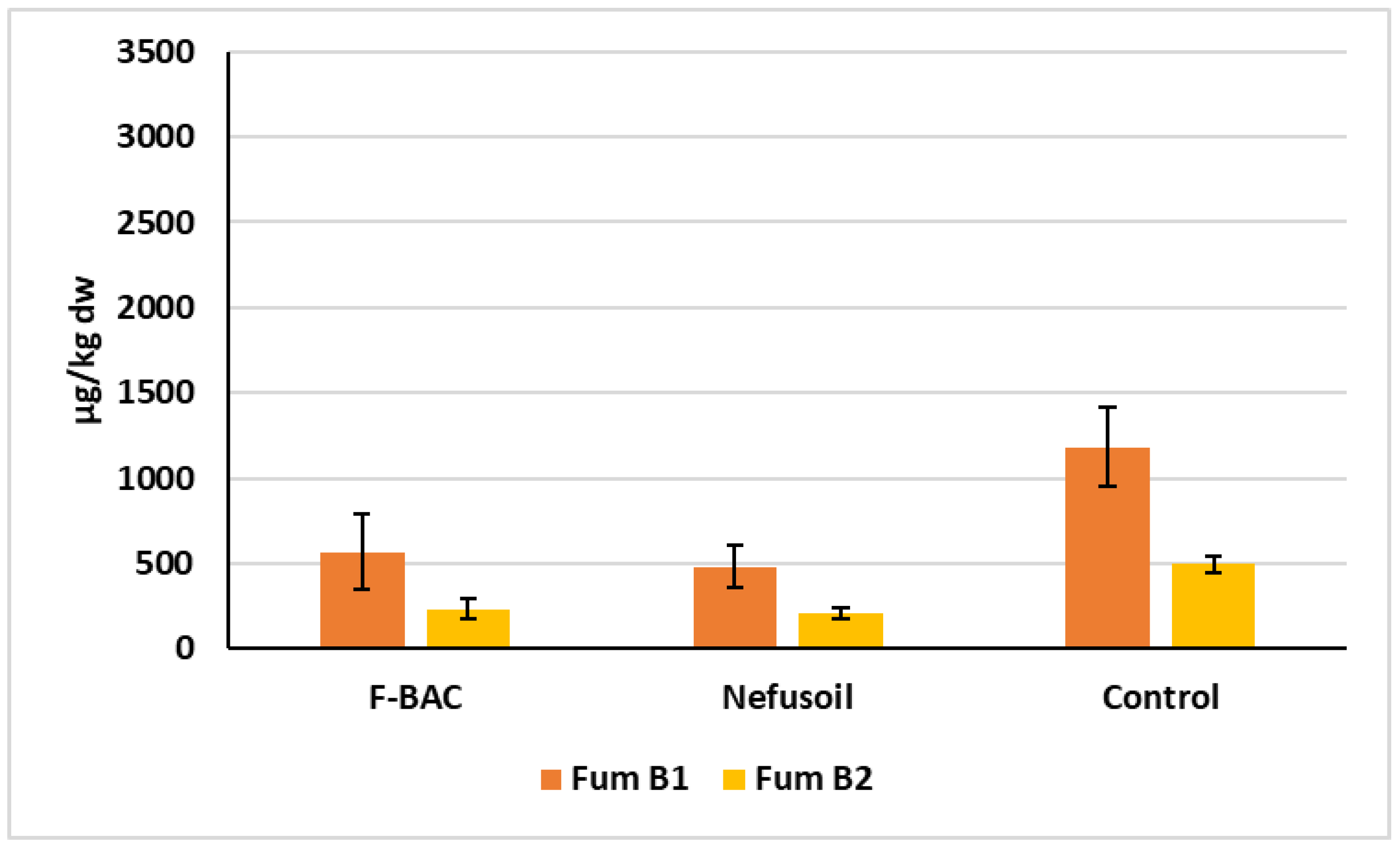
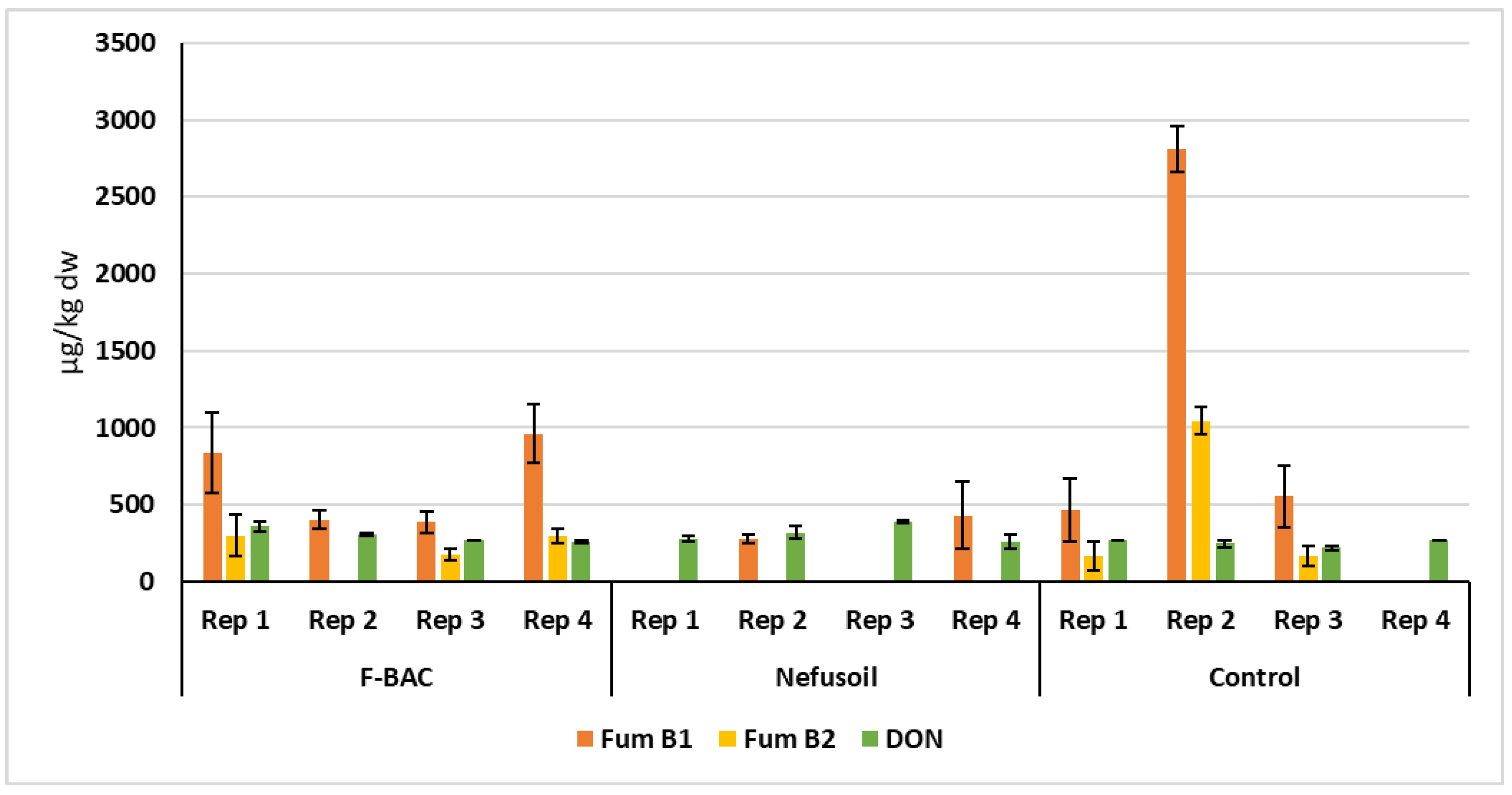
2.2. Analyses of Mycotoxins
3. Discussion
4. Conclusions
5. Materials and Methods
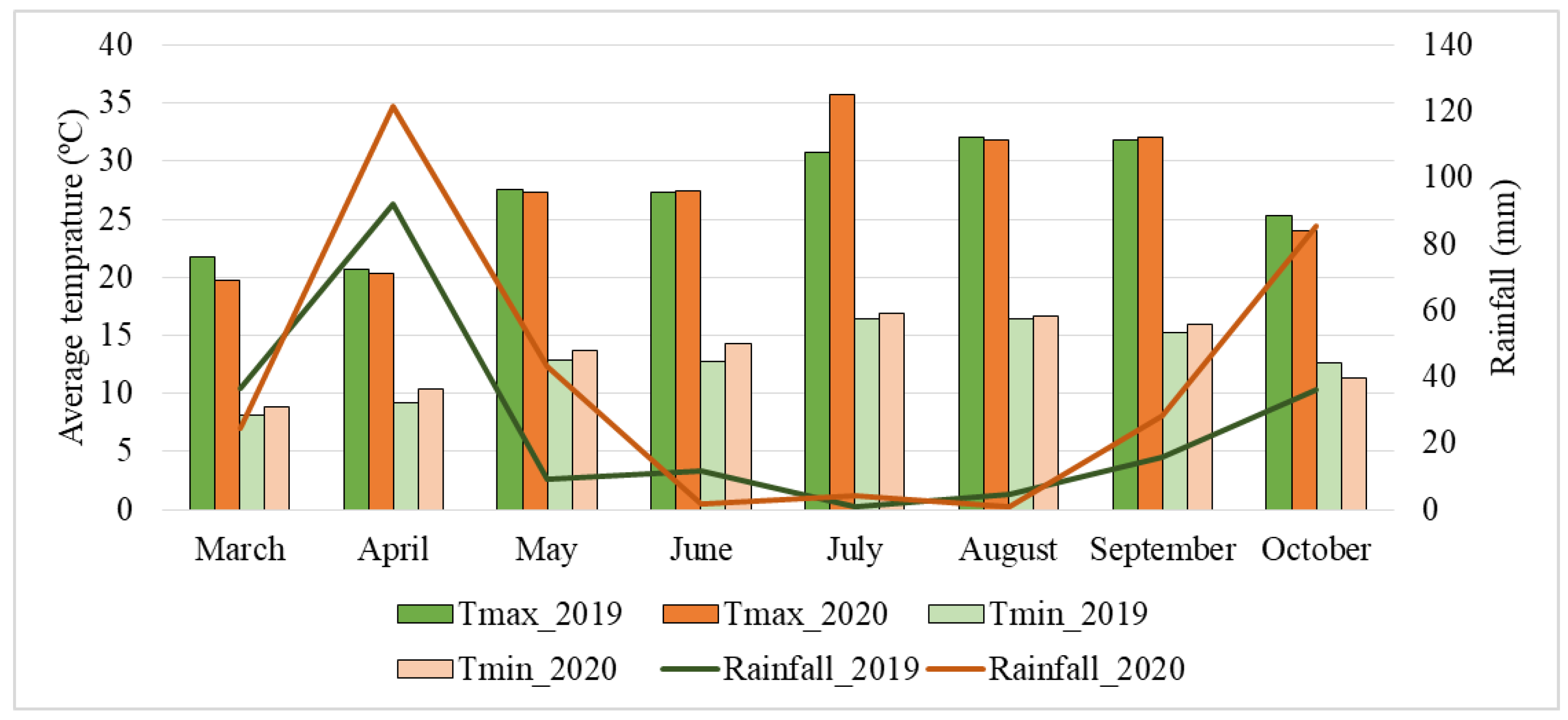
5.1. Maize Cultivation Conditions
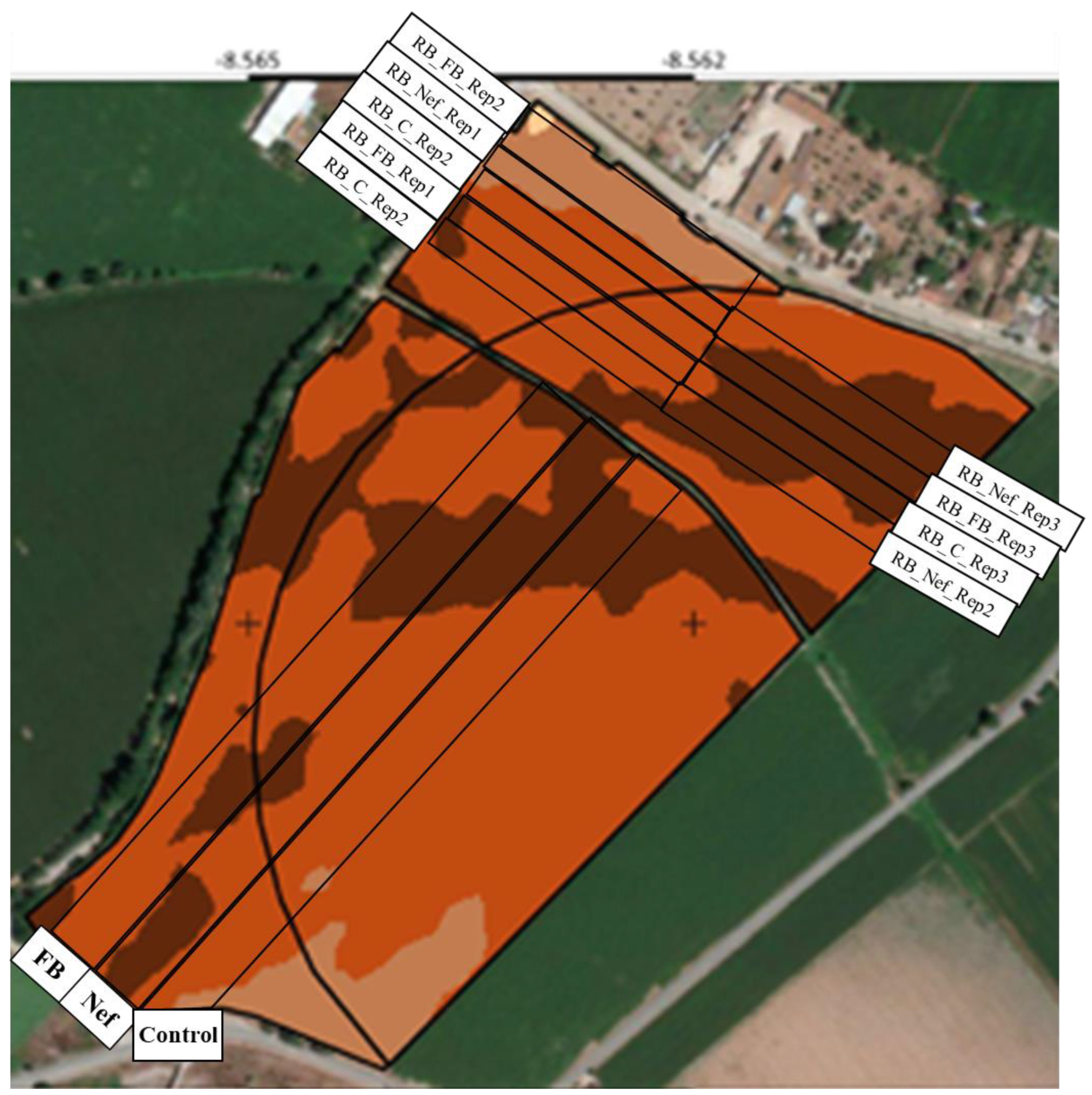
5.2. Crop Harvest and Sampling
5.3. Fungal Detection, Identification, and Quantification
5.4. DNA Extraction and Molecular Identification
5.5. Mycotoxins’ Extraction
5.6. Mycotoxin UHPLC-ToF-MS Analyses
5.7. Deoxynivalenol (DON) Analyses
5.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, J.H.; Lee, S.; Nah, J.Y.; Kim, H.K.; Paek, J.S.; Lee, S.; Ham, H.; Hong, S.K.; Yun, S.H.; Lee, T. Species composition of and fumonisin production by the Fusarium fujikuroi species complex isolated from Korean cereals. Int. J. Food Microbiol. 2018, 267, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dorn, B.; Forrer, H.R.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Vogelgsang, S. Fusarium species complex and mycotoxins in grain maize from maize hybrid trials and from grower’s fields. J. Appl. Microbiol. 2011, 111, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Estatísticas Agrícolas—2020; Instituto Nacional de Estatística I.P.: Lisboa, Portugal, 2021; ISBN 978-989-25-0572-5.
- Aguín, O.; Cao, A.; Pintos, C.; Santiago, R.; Mansilla, P.; Butrón, A. Occurrence of Fusarium species in maize kernels grown in northwestern Spain. Plant Pathol. 2014, 63, 946–951. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2006; ISBN 9780813819198. [Google Scholar]
- Carbas, B.; Simões, D.; Soares, A.; Freitas, A.; Ferreira, B.; Carvalho, A.R.F.; Silva, A.S.; Pinto, T.; Diogo, E.; Andrade, E.; et al. Occurrence of fusarium spp. In maize grain harvested in portugal and accumulation of related mycotoxins during storage. Foods 2021, 10, 375. [Google Scholar] [CrossRef]
- Simões, D.; Diogo, E.; de Andrade, E. First Report of Fusarium andiyazi Presence in Portuguese Maize Kernels. Agriculture 2022, 12, 336. [Google Scholar] [CrossRef]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; De Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic potentials of Fusarium species in various culture matrices revealed by mycotoxin profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef]
- Afsah-Hejri, L.; Jinap, S.; Hajeb, P.; Radu, S.; Shakibazadeh, S. A review on mycotoxins in food and feed: Malaysia case study. Compr. Rev. Food Sci. Food Saf. 2013, 12, 629–651. [Google Scholar] [CrossRef]
- Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Reduction of mycotoxins and toxigenic fungi in the mediterranean basin maize chain. Phytopathol. Mediterr. 2012, 51, 93–118. [Google Scholar] [CrossRef]
- Guerre, P. Worldwide mycotoxins exposure in pig and poultry feed formulations. Toxins 2016, 8, 350. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Carbas, B.; Simões, D.; Soares, A.; Freitas, A.; Carvalho, A.; Silva, A.S.; Pinto, T.; Varela, M.; Semedo, J.; Covão, S.; et al. Estratégias de Mitigação da Incidência de Micotoxinas durante o Armazenamento do Milho. Vida Rural 2021, 1869, 56–63. [Google Scholar]
- Carbas, B.; Soares, A.; Barros, S.; Carqueijo, A.; Freitas, A.; Silva, A.S.; Simões, D.; Pinto, T.; de Andrade, E.; Brites, C. Evaluation of Biobased Solutions for Mycotoxin Mitigation on stored maize. Chem. Proc. 2022, 10, 22. [Google Scholar] [CrossRef]
- Deepthi, B.V.; Deepa, N.; Vanitha, P.R.; Sreenivasa, M.Y. Stress responses on the growth and mycotoxin biosynthesis of Fusarium proliferatum associated with stored poultry feeds. Appl. Food Res. 2022, 2, 100091. [Google Scholar] [CrossRef]
- Alsamir, M.; Al-Samir, E.; Kareem, T.A.; Abass, M.; Trethowan, R. The application of zinc fertilizer reduces Fusarium infection and development in wheat. Aust. J. Crop Sci. 2020, 14, 1088–1094. [Google Scholar] [CrossRef]
- Nowembabaz, A.; Taulya, G.; Tinzaara, W.; Karamura, E. Effect of integrated potassium nutrition on Fusarium wilt tolerance in apple bananas. Afr. J. Plant Sci. 2021, 15, 257–265. [Google Scholar] [CrossRef]
- Gilardi, G.; Vasileiadou, A.; Garibaldi, A.; Gullino, M.L. The effects of biological control agents, potassium phosphite and calcium oxide on race 1 of Fusarium oxysporum f. sp. lactucae of lettuce in closed soilless cultivation systems. J. Phytopathol. 2022, 170, 626–634. [Google Scholar] [CrossRef]
- EIBOL Nefusoil. Available online: https://www.eibol.com/pt-pt/productos/nefusoil-regenerador-suelo-mantiene-equilibrio-biologico/ (accessed on 26 April 2022).
- EIBOL F-BAC. Available online: https://www.eibol.com/pt-pt/productos/f-bac-inductor-autodefensa-para-todo-tipo-cultivos/ (accessed on 26 April 2022).
- Brera, C.; Caputi, R.; Miraglia, M.; Iavicoli, I.; Salerno, A.; Carelli, G. Exposure assessment to mycotoxins in workplaces: Aflatoxins and ochratoxin A occurrence in airborne dusts and human sera. Microchem. J. 2002, 73, 167–173. [Google Scholar] [CrossRef]
- Mayer, S.; Engelhart, S.; Kolk, A.; Blome, H. The significance of mycotoxins in the framework of assessing workplace related risks. Mycotoxin Res. 2008, 24, 151–164. [Google Scholar] [CrossRef]
- Alvito, P.; Pereira-Da-silva, L. Mycotoxin Exposure during the First 1000 Days of Life and Its Impact on Children’s Health: A Clinical Overview. Toxins 2022, 14, 189. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef]
- Fandohan, P.; Hell, K.; Marasas, W.F.O.; Wingfield, M.J. Infection of maize by Fusarium species and contamination with fumonisin in Africa. Afr. J. Biotechnol. 2003, 2, 646–665. [Google Scholar]
- Bocianowski, J.; Szulc, P.; Wáskiewicz, A.; Cyplik, A. The effect of agrotechnical factors on Fusarium mycotoxins level in maize. Agriculture 2020, 10, 528. [Google Scholar] [CrossRef]
- Stepień, L.; Koczyk, G.; Waśkiewicz, A. FUM cluster divergence in fumonisins-producing Fusarium species. Fungal Biol. 2011, 115, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Ogoshi, C.; Scheuermann, K.K.; Silva-Lobo, V.L.; Schurt, D.A.; Ritieni, A.; Moretti, A.; Pfenning, L.H.; et al. Nationwide survey reveals high diversity of Fusarium species and related mycotoxins in Brazilian rice: 2014 and 2015 harvests. Food Control 2020, 113, 107171. [Google Scholar] [CrossRef]
- Picot, A.; Hourcade-Marcolla, D.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Richard-Forget, F.; Lannou, C. Interactions between Fusarium verticillioides and Fusarium graminearum in maize ears and consequences for fungal development and mycotoxin accumulation. Plant Pathol. 2012, 61, 140–151. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium Head Blight From a Microbiome Perspective. Front. Microbiol. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- European Union. Commission of the European Communities Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L364/5, 5–24. [Google Scholar]
- Carbas, B.; Soares, A.; Freitas, A.; Silva, A.S.; Pinto, T.; Andrade, E.; Brites, C. Mycotoxin Incidence in Pre-Harvest Maize Grains. Proceedings 2020, 70, 24. [Google Scholar] [CrossRef]
- Krysińska-Traczyk, E.; Kiecana, I.; Perkowski, J.; Dutkiewicz, J. Levels of fungi and mycotoxins in samples of grain and grain dust collected on farms in eastern Poland. Ann. Agric. Environ. Med. 2001, 8, 269–274. [Google Scholar]
- Carvajal-Moreno, M. Mycotoxin challenges in maize production and possible control methods in the 21st century. J. Cereal Sci. 2022, 103, 103293. [Google Scholar] [CrossRef]
- Cunha, S.C.; Fernandes, J.O. Development and validation of a method based on a QuEChERS procedure and heart-cutting GC-MS for determination of five mycotoxins in cereal products. J. Sep. Sci. 2010, 33, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Cegielska-Radziejewska, R.; Stuper, K.; Szablewski, T. Microflora and mycotoxin contamination in poultry feed mixtures from western Poland. Ann. Agric. Environ. Med. 2013, 20, 30–35. [Google Scholar] [PubMed]
- Castellá, G.; Bragulat, M.R.; Rubiales, M.V.; Cabañes, F.J. Malachite green agar, a new selective medium for Fusarium spp. Mycopathologia 1997, 137, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.S.; Aveling, T.A.S.; Blanco Prieto, R. A new semi-selective medium for Fusarium graminearum, F. proliferatum, F. subglutinans and F. verticillioides in maize seed. S. Afr. J. Bot. 2013, 84, 94–101. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Biotechniques 30th anniversary gem Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 2013, 54, 506–513. [Google Scholar] [CrossRef]
- Brewer, M.T.; Milgroom, M.G. Phylogeography and population structure of the grape powdery mildew fungus, Erysiphe necator, from diverse Vitis species. BMC Evol. Biol. 2010, 10, 268. [Google Scholar] [CrossRef]
- Silva, A.S.; Brites, C.; Pouca, A.V.; Barbosa, J.; Freitas, A. UHPLC-ToF-MS method for determination of multi-mycotoxins in maize: Development and validation. Curr. Res. Food Sci. 2019, 1, 1–7. [Google Scholar] [CrossRef]
- Freitas, A.; Barros, S.; Brites, C.; Barbosa, J.; Silva, A.S. Validation of a Biochip Chemiluminescent Immunoassay for Multi-Mycotoxins Screening in Maize (Zea mays L.). Food Anal. Methods 2019, 12, 2675–2684. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Sample (Year) | Fum B1 Mean (µg/Kg dw) | Variation between Years * (%) | Fum B2 Mean (µg/Kg dw) | Variation between Years * (%) | Deoxynivalenol Mean (µg/Kg dw) |
|---|---|---|---|---|---|---|
| F-BAC | ||||||
| Long parcel 2019 | 568.8 | 12.0 | 231.5 | 16.6 | n.d. | |
| Long parcel 2020 | 646.1 | 193.1 | 298.4 | |||
| Random blocks | 509.6 | 125.2 | 382.5 | |||
| Nefusoil | ||||||
| Long parcel 2019 | 480.8 | 63.1 | 207.6 | 100.0 | n.d | |
| Long parcel 2020 | 177.2 | n.d | 309.8 | |||
| Random blocks | 85.3 | n.d | 335.3 | |||
| Control | ||||||
| Long parcel 2019 | 1182.4 | 19.1 | 495.7 | 30.6 | n.d | |
| Long parcel 2020 | 956.4 | 343.9 | 251.6 | |||
| Random blocks | 560.5 | 124.0 | 343.8 | |||
| Years before | ||||||
| Survey 2018 [6] | 1666.75 | 922.7 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, D.; Carbas, B.; Soares, A.; Freitas, A.; Silva, A.S.; Brites, C.; Andrade, E.d. Assessment of Agricultural Practices for Controlling Fusarium and Mycotoxins Contamination on Maize Grains: Exploratory Study in Maize Farms. Toxins 2023, 15, 136. https://doi.org/10.3390/toxins15020136
Simões D, Carbas B, Soares A, Freitas A, Silva AS, Brites C, Andrade Ed. Assessment of Agricultural Practices for Controlling Fusarium and Mycotoxins Contamination on Maize Grains: Exploratory Study in Maize Farms. Toxins. 2023; 15(2):136. https://doi.org/10.3390/toxins15020136
Chicago/Turabian StyleSimões, Daniela, Bruna Carbas, Andreia Soares, Andreia Freitas, Ana Sanches Silva, Carla Brites, and Eugénia de Andrade. 2023. "Assessment of Agricultural Practices for Controlling Fusarium and Mycotoxins Contamination on Maize Grains: Exploratory Study in Maize Farms" Toxins 15, no. 2: 136. https://doi.org/10.3390/toxins15020136
APA StyleSimões, D., Carbas, B., Soares, A., Freitas, A., Silva, A. S., Brites, C., & Andrade, E. d. (2023). Assessment of Agricultural Practices for Controlling Fusarium and Mycotoxins Contamination on Maize Grains: Exploratory Study in Maize Farms. Toxins, 15(2), 136. https://doi.org/10.3390/toxins15020136