Lake Avernus Has Turned Red: Bioindicator Monitoring Unveils the Secrets of “Gates of Hades”

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
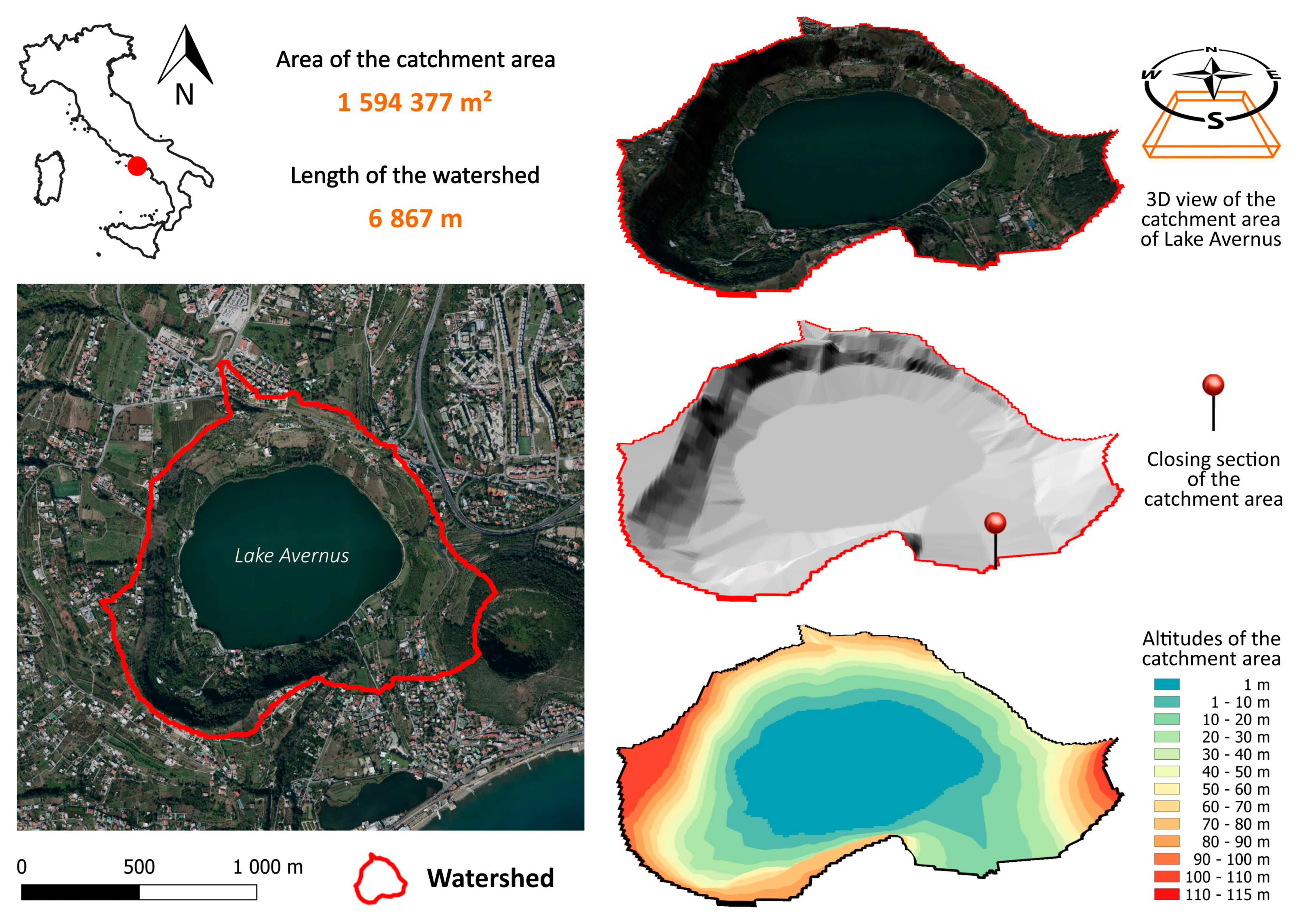
2.1. Remote/Proximal Sensing and Analysis of Multispectral Data
2.2. Taxonomic Analysis of Prevalent Cyanobacteria
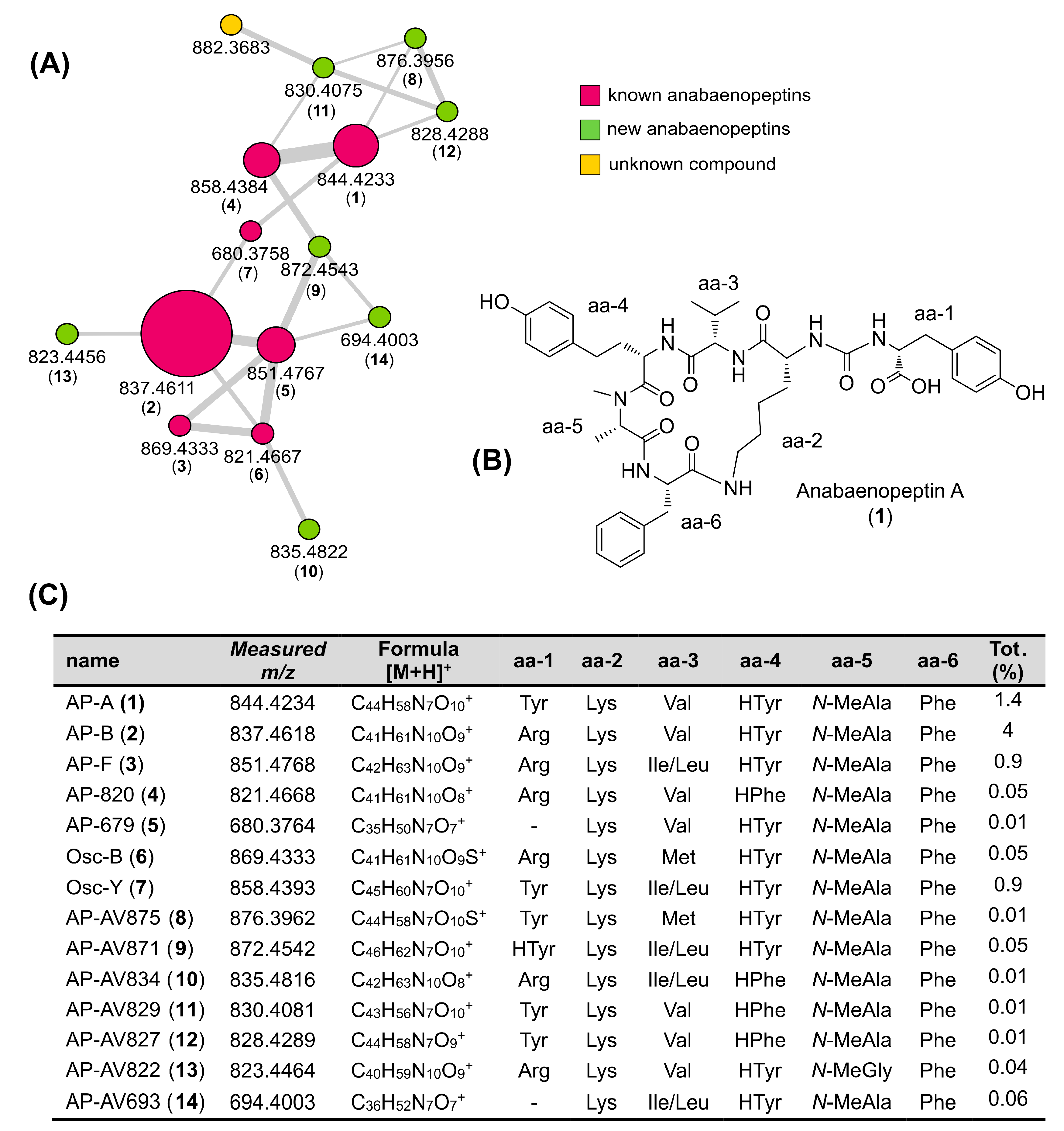
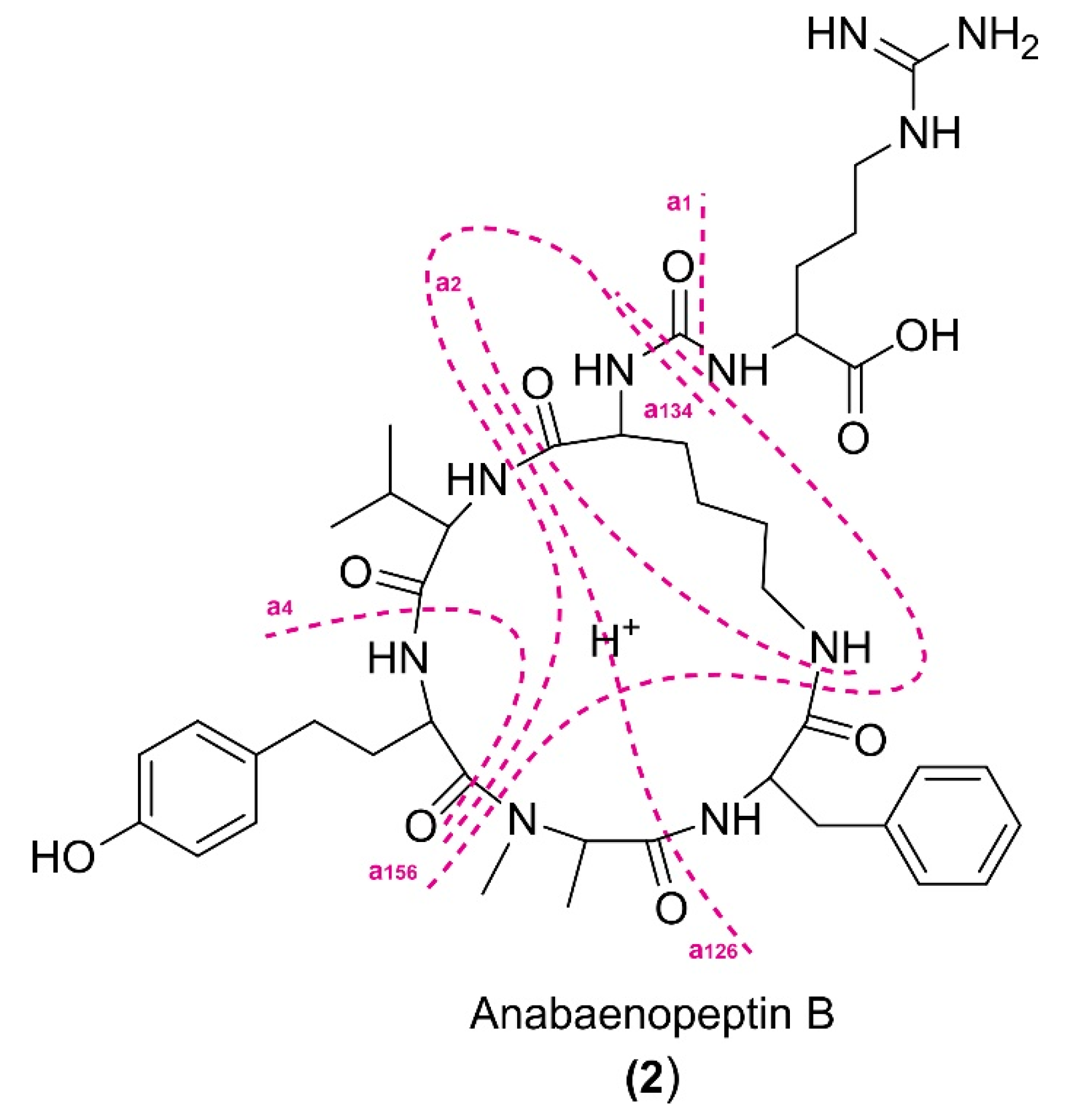
2.3. Molecular Networking and Identification of Anabaenopeptin Variants
3. Discussion and Conclusions
4. Materials and Methods
4.1. Satellite Remote Sensing and Analysis of Related Multispectral Data
4.2. Drone Proximal Sensing and Analysis of Related Multispectral Data
4.3. In-Situ Sampling and In-Lab Taxonomy
4.4. Metabolomic Profiling through HRMS/MS-Based Molecular Networking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mateo, P.; Leganés, F.; Perona, E.; Loza, V.; Fernández-Piñas, F. Cyanobacteria as Bioindicators and Bioreporters of Environmental Analysis in Aquatic Ecosystems. Biodivers. Conserv. 2015, 24, 909–948. [Google Scholar] [CrossRef]
- Teta, R.; Romano, V.; Della Sala, G.; Picchio, S.; De Sterlich, C.; Mangoni, A.; Di Tullio, G.; Costantino, V.; Lega, M. Cyanobacteria as Indicators of Water Quality in Campania Coasts, Italy: A Monitoring Strategy Combining Remote/Proximal Sensing and in Situ Data. Environ. Res. Lett. 2017, 12, 024001. [Google Scholar] [CrossRef]
- Esposito, G.; Teta, R.; Marrone, R.; De Sterlich, C.; Casazza, M.; Anastasio, A.; Lega, M.; Costantino, V. A Fast Detection Strategy for Cyanobacterial Blooms and Associated Cyanotoxins (FDSCC) Reveals the Occurrence of Lyngbyatoxin A in Campania (South Italy). Chemosphere 2019, 225, 342–351. [Google Scholar] [CrossRef]
- Caliro, S.; Chiodini, G.; Izzo, G.; Minopoli, C.; Signorini, A.; Avino, R.; Granieri, D. Geochemical and Biochemical Evidence of Lake Overturn and Fish Kill at Lake Averno, Italy. J. Volcanol. Geotherm. Res. 2008, 178, 305–316. [Google Scholar] [CrossRef]
- Ferranti, P.; Fabbrocino, S.; Cerulo, M.G.; Bruno, M.; Serpe, L.; Gallo, P. Characterisation of Biotoxins Produced by a Cyanobacteria Bloom in Lake Averno Using Two LC–MS-Based Techniques. Food Addit. Contam. Part A 2008, 25, 1530–1537. [Google Scholar] [CrossRef]
- Teta, R.; Della Sala, G.; Esposito, G.; Stornaiuolo, M.; Scarpato, S.; Casazza, M.; Anastasio, A.; Lega, M.; Costantino, V. Monitoring Cyanobacterial Blooms during the COVID-19 Pandemic in Campania, Italy: The Case of Lake Avernus. Toxins 2021, 13, 471. [Google Scholar] [CrossRef]
- Anagnostidis, K. Modern Approach to the Classification System of Cyanophytes 3—Oscillatoriales. Arch. Hydrobiol. 1985, 80, 327–472. [Google Scholar]
- Harada, K.; Fujii, K.; Shimada, T.; Suzuki, M.; Sano, H.; Adachi, K.; Carmichael, W.W. Two Cyclic Peptides, Anabaenopeptins, a Third Group of Bioactive Compounds from the cyanobacterium Anabaena Flos-Aquae NRC 525-17. Tetrahedron Lett. 1995, 36, 1511–1514. [Google Scholar] [CrossRef]
- Monteiro, P.R.; do Amaral, S.C.; Siqueira, A.S.; Xavier, L.P.; Santos, A.V. Anabaenopeptins: What We Know So Far. Toxins 2021, 13, 522. [Google Scholar] [CrossRef]
- Paganin, P.; Chiarini, L.; Bevivino, A.; Dalmastri, C.; Farcomeni, A.; Izzo, G.; Signorini, A.; Varrone, C.; Tabacchioni, S. Vertical Distribution of Bacterioplankton in Lake Averno in Relation to Water Chemistry. FEMS Microbiol. Ecol. 2013, 84, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. In NASA Special Publication; NASA: Washington, DC, USA, 1974; Volume 351, p. 309. [Google Scholar]
- Singh, R.; Saritha, V.; Pande, C.B. Monitoring of Wetland Turbidity Using Multi-Temporal Landsat-8 and Landsat-9 Satellite Imagery in the Bisalpur Wetland, Rajasthan, India. Environ. Res. 2023, 241, 117638. [Google Scholar] [CrossRef]
- Tarquini, S.; Isola, I.; Favalli, M.; Battistini, A. TINITALY, a Digital Elevation Model of Italy with a 10 Meters Cell Size ; about 3000 M Grid Cells, about 17 Gb of Disk Memory; Istituto Nazionale di Geofisica e Vulcanologia (INGV): Rome, Italy, 2007.
- Lega, M.; Endreny, T. Quantifying the Environmental Impact of Pollutant Plumes from Coastal Rivers with Remote Sensing and River Basin Modelling. Int. J. Sustain. Dev. Plan. 2016, 11, 651–662. [Google Scholar] [CrossRef]
- Komárek, J.; Johansen, J.R. Filamentous Cyanobacteria. In Freshwater Algae of North America; Elsevier: Cambridge, MA, USA, 2015; pp. 135–235. ISBN 978-0-12-385876-4. [Google Scholar]
- Available online: https://gnps.ucsd.edu/ProteoSAFe/status.jsp?task=dab305d4d24e4123bb455a0144da2ba1 (accessed on 4 November 2022).
- Available online: https://gnps.ucsd.edu/ProteoSAFe/result.jsp?task=d60eaeed14644b62b0933ce0b158448a&view=view_significant_unique (accessed on 4 November 2022).
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Microbial Metabolites through Database Search of Mass Spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef]
- Murakami, M.; Shin, H.J.; Matsuda, H.; Ishida, K.; Yamaguchi, K. A Cyclic Peptide, Anabaenopeptin B, from the Cyanobacterium Oscillatoria Agardhii. Phytochemistry 1997, 44, 449–452. [Google Scholar] [CrossRef]
- Ngoka, L.C.M.; Gross, M.L. Multistep Tandem Mass Spectrometry for Sequencing Cyclic Peptides in an Ion-Trap Mass Spectrometer. J. Am. Soc. Mass Spectrom. 1999, 10, 732–746. [Google Scholar] [CrossRef]
- Ngoka, L.C.M.; Gross, M.L. A Nomenclature System for Labeling Cyclic Peptide Fragments. J. Am. Soc. Mass Spectrom. 1999, 10, 360–363. [Google Scholar] [CrossRef]
- Zervou, S.-K.; Kaloudis, T.; Gkelis, S.; Hiskia, A.; Mazur-Marzec, H. Anabaenopeptins from Cyanobacteria in Freshwater Bodies of Greece. Toxins 2021, 14, 4. [Google Scholar] [CrossRef]
- Harms, H.; Kurita, K.L.; Pan, L.; Wahome, P.G.; He, H.; Kinghorn, A.D.; Carter, G.T.; Linington, R.G. Discovery of Anabaenopeptin 679 from Freshwater Algal Bloom Material: Insights into the Structure–Activity Relationship of Anabaenopeptin Protease Inhibitors. Bioorganic Med. Chem. Lett. 2016, 26, 4960–4965. [Google Scholar] [CrossRef]
- Sanz, M.; Andreote, A.; Fiore, M.; Dörr, F.; Pinto, E. Structural Characterization of New Peptide Variants Produced by Cyanobacteria from the Brazilian Atlantic Coastal Forest Using Liquid Chromatography Coupled to Quadrupole Time-of-Flight Tandem Mass Spectrometry. Mar. Drugs 2015, 13, 3892–3919. [Google Scholar] [CrossRef] [PubMed]
- Entfellner, E.; Frei, M.; Christiansen, G.; Deng, L.; Blom, J.; Kurmayer, R. Evolution of Anabaenopeptin Peptide Structural Variability in the Cyanobacterium Planktothrix. Front. Microbiol. 2017, 8, 219. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.M.-L. Cyanobacterial Peptides beyond Microcystins—A Review on Co-Occurrence, Toxicity, and Challenges for Risk Assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef]
- Tassi, F.; Fazi, S.; Rossetti, S.; Pratesi, P.; Ceccotti, M.; Cabassi, J.; Capecchiacci, F.; Venturi, S.; Vaselli, O. The Biogeochemical Vertical Structure Renders a Meromictic Volcanic Lake a Trap for Geogenic CO2 (Lake Averno, Italy). PLoS ONE 2018, 13, e0193914. [Google Scholar] [CrossRef]
- Lega, M.; Casazza, M.; Teta, R.; Zappa, C.J. Environmental Impact Assessment: A Multilevel, Multi-Parametric Framework for Coastal Waters. Int. J. Sustain. Dev. Plan. 2018, 13, 1041–1049. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Available online: https://sentinel.esa.int/web/sentinel/missions/sentinel-2 (accessed on 30 March 2022).
- Boutte, C.; Grubisic, S.; Balthasart, P.; Wilmotte, A. Testing of Primers for the Study of Cyanobacterial Molecular Diversity by DGGE. J. Microbiol. Methods 2006, 65, 542–550. [Google Scholar] [CrossRef]
- Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 26 July 2022).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Demchak, B.; Ideker, T. Cytoscape Tools for the Web Age: D3.js and Cytoscape.js Exporters. F1000Research 2014, 3, 143. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragment ion | AP-A (1) | AP-B (2) | Osc-B (3) | Osc-Y (4) | AP-F (5) | AP-820 (6) | AP-679 (7) | AP-AV875 (8) | AP-AV871 (9) | AP-AV834 (10) | AP-AV829 (11) | AP-AV827 (12) | AP-AV822 (13) | AP-AV693 (14) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [M + H]+ | 844.4233 C44H58N7O10+ | 837.4611 C41H61N10O9+ | 869.4333 C41H61N10O9S+ | 858.4384 C45H60N7O10+ | 851.4767 C42H63N10O9+ | 821.4668 C41H61N10O8+ | 680.3758 C35H50N7O7+ | 876.3956 C44H58N7O10S+ | 872.4543 C46H62N7O10+ | 835.4822 C42H63N10O8+ | 830.4075 C43H56N7O10+ | 828.4288 C44H58N7O9+ | 823.4456 C40H59N10O9+ | 694.4003 C36H52N7O7+ |
| [M + 2H]2+ | n.o. | 419.2344 C41H62N10O92+ | 435.2200 C41H62N10O9S2+ | n.o. | 426.2420 C42H64N10O92+ | 411.2368 C41H62N10O82+ | n.o. | n.o. | n.o. | 418.2443 C42H64N10O82+ | n.o. | n.o. | 412.2288 C40H60N10O92+ | n.o. |
| a1 | 663.3494 C35H47N6O7+ | 663.3496 C35H47N6O7+ | 695.3209 C35H47N6O7S+ | 677.3651 C36H49N6O7+ | n.o. | 647.3547 C35H47N6O6+ | 663.3520 C35H47N6O7+ | 695.3210 C35H47N6O7S+ | 677.3644 C36H49N6O7+ | n.o. | 649.3358 C34H45N6O7+ | 647.3565 C35H47N6O6+ | 649.3361 C35H47N6O7+ | 677.3678 C36H49N6O7+ |
| a1-CO | 637.3700 C34H49N6O6+ | 637.3700 C34H49N6O6+ | 669.3414 C34H49N6O6S+ | 651.3860 C35H51N6O6+ | 651.3856 C35H51N6O6+ | 621.3739 C34H49N6O5+ | 637.3727 C34H49N6O6+ | 669.3417 C34H49N6O6S+ | 651.3853 C35H51N6O6+ | 635.3898 C35H51N6O5+ | 623.3562 C33H47N6O6+ | 621.3772 C34H49N6O5+ | 623.3562 C33H47N6O6+ | 651.3883 C35H51N6O6+ |
| a2 | 509.2753 C28H37N4O5+ | 509.2753 C28H37N4O5+ | n.o. | 523.2907 C29H39N4O5+ | 523.2905 C29H39N4O5+ | 493.2794 C28H37N4O4+ | 509.2773 C28H37N4O5+ | 541.2465 C28H37N4O5S+ | 523.2903 C29H39N4O5+ | n.o. | 495.2606 C27H35N4O5+ | 493.2817 C28H37N4O4+ | 495.2608 C27H35N4O5+ | 523.2933 C29H39N4O5+ |
| a3 | 745.3551 C39H49N6O9+ | n.o. | n.o. | 745.3556 C39H49N6O9+ | n.o. | n.o. | n.o. | n.o. | 759.3680 C40H51N6O9+ | n.o. | 731.3419 C38H47N6O9+ | n.o. | 724.3792 C35H50N9O8+ | n.o. |
| a4 | 667.3443 C34H47N6O8+ | 660.3827 C31H50N9O7+ | 692.3532 C31H50N9O7S+ | 681.3602 C35H49N6O8+ | 674.3980 C32H52N9O7+ | n.o. | 503.2991 C25H39N6O5+ | 699.3160 C34H47N6O8S+ | 695.3751 C36H51N6O8+ | 674.3971 C32H52N9O7+ | n.o | 667.3468 C34H47N6O8+ | 646.3658 C30H48N9O7+ | 517.3123 C26H41N6O5+ |
| a5 | 759.3706 C40H51N6O9+ | 752.4084 C37H54N9O8+ | 784.3798 C37H54N9O8S+ | 773.3864 C41H53N6O9+ | 766.4242 C38H56N9O8+ | 736.4126 C37H54N9O7+ | n.o. | 791.3450 C40H51N6O9S+ | 787.4025 C42H55N6O9+ | 750.4283 C38H56N9O7+ | 759.3699 C40H51N6O9+ | 743.3790 C40H51N6O8+ | n.o. | n.o. |
| a6-H2O | 679.3445 C35H47N6O8+ | 672.3821 C32H50N9O7+ | n.o. | 693.3613 C36H49N6O8+ | 686.3981 C33H52N9O7+ | n.o. | n.o. | n.o. | n.o. | n.o. | n.o. | 663.3494 C35H47N6O7+ | n.o. | n.o. |
| a14-CO | 460.2912 C24H38N5O4+ | 460.2914 C24H38N5O4+ | 492.2621 C24H38N5O4S+ | 474.3068 C25H40N5O4+ | 474.3068 C25H40N5O4+ | 460.2908 C24H38N5O4+ | n.o. | n.o. | 474.3057 C25H40N5O4+ | n.o. | n.o. | n.o. | 446.2777 C23H36N5O4+ | n.o. |
| a34 | 568.2759 C29H38N5O7+ | 561.3142 C26H41N8O6+ | 561.3128 C26H41N8O6+ | 568.2761 C29H38N5O7+ | 561.3140 C26H41N8O6+ | n.o. | 404.2306 C20H30N5O4+ | 568.2762 C29H38N5O7+ | 582.2911 C30H40N5O7+ | n.o. | 554.2619 C28H36N5O7+ | 568.2776 C29H38N5O7+ | 547.2983 C25H39N8O6+ | 404.2308 C20H30N5O4+ |
| a34-H2O | 550.2654 C29H36N5O6+ | 543.3035 C26H39N8O5+ | 543.3025 C26H39N8O5+ | 550.2656 C29H36N5O6+ | 543.3033 C26H39N8O5+ | 543.3024 C26H39N8O5+ | n.o. | 550.2650 C29H36N5O6+ | n.o. | 543.3033 C26H39N8O5+ | 536.2494 C28H34N5O6+ | 550.2672 C29H36N5O6+ | 529.2898 C25H37N8O5+ | n.o. |
| a45 | 582.2916 C30H40N5O7+ | 575.3297 C27H43N8O6+ | 607.3010 C27H43N8O6S+ | 596.3074 C31H42N5O7+ | 589.3452 C28H45N8O6+ | 575.3285 C27H43N8O6+ | 418.2458 C21H32N5O4+ | n.o. | 610.3218 C32H44N5O7+ | 589.3442 C28H45N8O6+ | 582.2936 C30H40N5O7+ | 582.2925 C30H40N5O7+ | 575.3289 C27H43N8O6+ | 432.2615 C22H34N5O4+ |
| a134 + H2O | 379.2337 C19H31N4O4+ | 379.2346 C19H31N4O4+ | n.o. | 379.2338 C19H31N4O4+ | 379.2337 C19H31N4O4+ | n.o. | n.o. | 379.2341 C19H31N4O4+ | n.o. | n.o. | n.o. | 379.2348 C19H31N4O4+ | n.o. | n.o. |
| a134 + H2O + CO | 405.2129 C20H29N4O5+ | n.o. | n.o. | 405.2130 C20H29N4O5+ | n.o. | n.o. | n.o. | 405.2132 C20H29N4O5+ | n.o. | n.o. | 391.1985 C19H27N4O5+ | 405.2143 C20H29N4O5+ | n.o. | n.o. |
| a126 | 362.2070 C19H28N3O4+ | 362.2073 C19H28N3O4+ | 394.1789 C19H28N3O4S+ | 376.2229 C20H30N3O4+ | 376.2227 C20H30N3O4+ | 346.2117 C19H28N3O3+ | 362.2085 C19H28N3O4+ | n.o. | 376.2224 C20H30N3O4+ | 360.2271 C20H30N3O3+ | 348.1920 C18H26N3O4+ | n.o. | 348.1926 C18H26N3O4+ | 376.2238 C20H30N3O4+ |
| a156 | n.o. | 405.2495 C21H33N4O4+ | n.o. | n.o. | 419.2648 C22H33N4O3+ | n.o. | n.o. | n.o. | n.o. | n.o. | n.o. | n.o. | n.o. | n.o. |
| a156-H2O | 387.2387 C21H31N4O3+ | 387.2386 C21H31N4O3+ | n.o. | 401.2539 C22H35N4O4+ | 401.2541 C22H35N4O4+ | n.o. | n.o. | n.o. | 401.2542 C22H35N4O4+ | n.o. | n.o. | n.o. | 387.2394 C21H31N4O3+ | n.o. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, G.; Glukhov, E.; Gerwick, W.H.; Medio, G.; Teta, R.; Lega, M.; Costantino, V. Lake Avernus Has Turned Red: Bioindicator Monitoring Unveils the Secrets of “Gates of Hades”. Toxins 2023, 15, 698. https://doi.org/10.3390/toxins15120698
Esposito G, Glukhov E, Gerwick WH, Medio G, Teta R, Lega M, Costantino V. Lake Avernus Has Turned Red: Bioindicator Monitoring Unveils the Secrets of “Gates of Hades”. Toxins. 2023; 15(12):698. https://doi.org/10.3390/toxins15120698
Chicago/Turabian StyleEsposito, Germana, Evgenia Glukhov, William H. Gerwick, Gabriele Medio, Roberta Teta, Massimiliano Lega, and Valeria Costantino. 2023. "Lake Avernus Has Turned Red: Bioindicator Monitoring Unveils the Secrets of “Gates of Hades”" Toxins 15, no. 12: 698. https://doi.org/10.3390/toxins15120698
APA StyleEsposito, G., Glukhov, E., Gerwick, W. H., Medio, G., Teta, R., Lega, M., & Costantino, V. (2023). Lake Avernus Has Turned Red: Bioindicator Monitoring Unveils the Secrets of “Gates of Hades”. Toxins, 15(12), 698. https://doi.org/10.3390/toxins15120698