A Direct Analysis of β-N-methylamino-l-alanine Enantiomers and Isomers and Its Application to Cyanobacteria and Marine Mollusks

 ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Source of Biological and Chemical Materials
4.2. Extraction and Purification of Neurotoxic Amino Acids
4.3. Analysis of AQC-Derivatized BMAA via UPLC-MS/MS
4.4. Analysis and Partial Validation of (S)-NIFE Derivatized Standards and Cyanobacterial Samples
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Co-occurrence of cyanobacteria and cyanotoxins with other environmental health hazards: Impacts and implications. Toxins 2020, 12, 629. [Google Scholar] [CrossRef] [PubMed]
- Altermann, W.; Kazmierczak, J.; Oren, A.; Wright, D.T. Cyanobacterial calcification and its rock-building potential during 3.5 billion years of Earth history. Geobiology 2006, 3, 147–166. [Google Scholar] [CrossRef]
- Sergeev, V.N.; Gerasimenko, L.M.; Zavarzin, G.A. The proterozoic history and present state of cyanobacteria. Microbiology 2002, 71, 623–637. [Google Scholar] [CrossRef]
- Schirrmeister, B.E.; Sanchez-Baracaldo, P.; Wacey, D. Cyanobacterial evolution during the Precambrian. Int. J. Astrobiol. 2016, 3, 187–204. [Google Scholar] [CrossRef]
- Schopf, J.W. The fossil record: Tracing the roots of the cyanobacterial lineage. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Springer BV: Dordrecht, The Netherlands, 2002; pp. 13–35. [Google Scholar]
- Schopf, J.W.; Packer, B.M. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 1987, 237, 70–73. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and biogeochemical cycles through Earth history. Trends Microbiol. 2022, 30, 143–157. [Google Scholar] [CrossRef]
- Knoll, A.H. Cyanobacteria and earth history. In The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Caister Academic Press: Poole, UK, 2008; 484p. [Google Scholar]
- Sánchez-Baracaldo, P.; Cardona, T. On the origin of oxygenic photosynthesis and Cyanobacteria. New Phytol. 2020, 4, 1440–1446. [Google Scholar] [CrossRef]
- Shih, P.M. Cyanobacterial evolution: Fresh insight into ancient questions. Curr. Biol. 2015, 5, R192–R193. [Google Scholar] [CrossRef]
- Nugumanova, G.; Ponomarev, E.D.; Askarova, S.; Fasler-Kan, E.; Barteneva, N.S. Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases. Toxins 2023, 15, 233. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 1, 31. [Google Scholar] [CrossRef]
- Svirčev, Z.; Chen, L.; Sántha, K.; Drobac Backović, D.; Šušak, S.; Vulin, A.; Palanački Malešević, T.; Codd, G.A.; Meriluoto, J.A. review and assessment of cyanobacterial toxins as cardiovascular health hazards. Arch. Toxicol. 2022, 11, 2829–2863. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 12, 714. [Google Scholar] [CrossRef]
- Shi, L.; Du, X.; Liu, H.; Chen, X.; Ma, Y.; Wang, R.; Tian, Z.; Zhang, S.; Guo, H.; Zhang, H. Update on the adverse effects of microcystins on the liver. Environ. Res. 2021, 195, 110890. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, H.; Du, X.; Chen, X.; Petlulu, P.; Tian, Z.; Shi, L.; Zhang, B.; Yuan, S.; Guo, X.; et al. A new identity of microcystins: Environmental endocrine disruptors? An evidence-based review. Sci. Total Environ. 2022, 851, 158262. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yi, X.; Liu, W.; Zhang, C.; Massey, I.Y.; Yang, F.; Tian, L. A review of nephrotoxicity of microcystins. Toxins 2020, 12, 693. [Google Scholar] [CrossRef] [PubMed]
- Schreidah, C.M.; Ratnayake, K.; Senarath, K.; Karunarathne, A. Microcystins: Biogenesis, toxicity, analysis, and control. Chem. Res. Toxicol. 2020, 9, 2225–2246. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, G.; Chen, Y.; Jia, N.; Li, R. Four decades of progress in cylindrospermopsin research: The ins and outs of a potent cyanotoxin. J. Hazard. Mater. 2021, 406, 124653. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef]
- Zhang, Y.; Duy, S.V.; Munoz, G.; Sauvé, S. Phytotoxic effects of microcystins, anatoxin-a and cylindrospermopsin to aquatic plants: A meta-analysis. Sci. Total Environ. 2022, 810, 152104. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Benítez-González, M.D.; Puerto, M.; Jos, A.; Cameán, A.M. Immunotoxic effects induced by microcystins and cylindrospermopsin: A review. Toxins 2021, 13, 711. [Google Scholar] [CrossRef]
- Scarlett, K.R.; Kim, S.; Lovin, L.M.; Chatterjee, S.; Scott, J.T.; Brooks, B.W. Global scanning of cylindrospermopsin: Critical review and analysis of aquatic occurrence, bioaccumulation, toxicity and health hazards. Sci. Total Environ. 2020, 738, 139807. [Google Scholar] [CrossRef]
- Casas-Rodriguez, A.; Cameán, A.M.; Jos, A. Potential endocrine disruption of cyanobacterial toxins, microcystins and cylindrospermopsin: A review. Toxins 2022, 14, 882. [Google Scholar] [CrossRef]
- Colas, S.; Marie, B.; Lance, E.; Quiblier, C.; Tricoire-Leignel, H.; Mattei, C. Anatoxin-a: Overview on a harmful cyanobacterial neurotoxin from the environmental scale to the molecular target. Environ. Res. 2021, 193, 110590. [Google Scholar] [CrossRef] [PubMed]
- Plata-Calzado, C.; Prieto, A.I.; Cameán, A.M.; Jos, A. Toxic Effects Produced by Anatoxin-a under Laboratory Conditions: A Review. Toxins 2022, 14, 861. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.; Botelho, M.J.; Churro, C.; Asselman, J.; Pereira, P.; Pereira, J.L. A review on aquatic toxins-Do we really know it all regarding the environmental risk posed by phytoplankton neurotoxins? J. Environ. Manag. 2023, 345, 118769. [Google Scholar] [CrossRef]
- Cirés, S.; Casero, M.C.; Quesada, A. Toxicity at the edge of life: A review on cyanobacterial toxins from extreme environments. Mar. Drugs 2017, 15, 233. [Google Scholar] [CrossRef] [PubMed]
- Florczyk, M.; Łakomiak, A.; Woźny, M.; Brzuzan, P. Neurotoxicity of cyanobacterial toxins. Environ. Biotechnol. 2014, 10, 26–43. [Google Scholar] [CrossRef]
- Falfushynska, H.; Kasianchuk, N.; Siemens, E.; Henao, E.; Rzymski, P. A review of common cyanotoxins and their effects on fish. Toxics 2023, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; Van Hassel, W.H.; Andjelkovic, M.; Wilmotte, A.; Rajkovic, A. Cyanotoxins and food contamination in developing countries: Review of their types, toxicity, analysis, occurrence and mitigation strategies. Toxins 2021, 13, 786. [Google Scholar] [CrossRef]
- Manganelli, M.; Testai, E.; Tazart, Z.; Scardala, S.; Codd, G.A. Co-Occurrence of Taste and Odor Compounds and Cyanotoxins in Cyanobacterial Blooms: Emerging Risks to Human Health? Microorganisms 2023, 11, 872. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.T.; Fallon, T.R.; Cordoza, J.L.; Chekan, J.R.; Delbaje, E.; Hopiavuori, A.R.; Alvarenga, D.O.; Wood, S.M.; Luhavaya, H.; Baumgartner, J.T.; et al. Biosynthesis of guanitoxin enables global environmental detection in freshwater cyanobacteria. J. Am. Chem. Soc. 2022, 144, 9372–9379. [Google Scholar] [CrossRef]
- Fiore, M.F.; de Lima, S.T.; Carmichael, W.W.; McKinnie, S.M.; Chekan, J.R.; Moore, B.S. Guanitoxin, re-naming a cyanobacterial organophosphate toxin. Harmful Algae 2020, 92, 101737. [Google Scholar] [CrossRef]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. Int. Ed. 2014, 53, 5760–5784. [Google Scholar] [CrossRef]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS ONE 2009, 4, e5758. [Google Scholar] [CrossRef]
- Hackett, J.D.; Wisecaver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of saxitoxin synthesis in cyanobacteria and dinoflagellates. Mol. Biol. Evol. 2013, 30, 70–78. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health 2006, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Durai, P.; Batool, M.; Choi, S. Structure and effects of cyanobacterial lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef] [PubMed]
- Gemma, S.; Molteni, M.; Rossetti, C. Lipopolysaccharides in cyanobacteria: A brief overview. Adv. Microbiol. 2016, 6, 391–397. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Simeunović, J.; Baltić, V.; Stanić, D.; Svirčev, Z. Human exposure to cyanotoxins and their effects on health. Arh. Za Hig. Rada I Toksikol. 2013, 64, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Funari, E.; Testai, E. Human health risk assessment related to cyanotoxins exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef]
- Skafi, M.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Simon, D.F.; Juneau, P.; Sauvé, S. Occurrence of microcystins, anabaenopeptins and other cyanotoxins in fish from a freshwater wildlife reserve impacted by harmful cyanobacterial blooms. Toxicon 2021, 194, 44–52. [Google Scholar] [CrossRef]
- Hardy, F.J.; Johnson, A.; Hamel, K.; Preece, E. Cyanotoxin bioaccumulation in freshwater fish, Washington State, USA. Environ. Monit. Assess. 2015, 187, 667. [Google Scholar] [CrossRef]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Labohá, P.; Sychrová, E.; Brózman, O.; Sovadinová, I.; Bláhová, L.; Prokeš, R.; Ondráček, J.; Babica, P. Cyanobacteria, cyanotoxins and lipopolysaccharides in aerosols from inland freshwater bodies and their effects on human bronchial cells. Environ. Toxicol. Pharmacol. 2023, 98, 104073. [Google Scholar] [CrossRef]
- Wood, S.A.; Dietrich, D.R. Quantitative assessment of aerosolized cyanobacterial toxins at two New Zealand lakes. J. Environ. Monit. 2011, 13, 1617–1624. [Google Scholar] [PubMed]
- Facciponte, D.N.; Bough, M.W.; Seidler, D.; Carroll, J.L.; Ashare, A.; Andrew, A.S.; Tsongalis, G.J.; Vaickus, L.J.; Henegan, P.L.; Butt, T.H.; et al. Identifying aerosolized cyanobacteria in the human respiratory tract: A proposed mechanism for cyanotoxin-associated diseases. Sci. Total Environ. 2018, 645, 1003–1013. [Google Scholar] [PubMed]
- Gaget, V.; Lau, M.; Sendall, B.; Froscio, S.; Humpage, A.R. Cyanotoxins: Which detection technique for an optimum risk assessment? Water Res. 2017, 118, 227–238. [Google Scholar]
- Yuan, J.; Yoon, K.-J. Overview of PCR Methods Applied for the Identification of Freshwater Toxigenic Cyanobacteria. Cyanobacteria-Recent Adv. Taxon. Appl. 2022, 1–24. [Google Scholar] [CrossRef]
- Pacheco, A.B.; Guedes, I.A.; Azevedo, S.M. Is qPCR a reliable indicator of cyanotoxin risk in freshwater? Toxins 2016, 8, 172. [Google Scholar]
- Ouellette, A.J.; Wilhelm, S.W. Toxic cyanobacteria: The evolving molecular toolbox. Front. Ecol. Environ. 2003, 1, 359–366. [Google Scholar]
- Jaramillo, M.; O’Shea, K.E. Analytical methods for assessment of cyanotoxin contamination in drinking water sources. Curr. Opin. Environ. Sci. Health 2019, 7, 45–51. [Google Scholar]
- Van Apeldoorn, M.E.; Van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar]
- Ramya, M.; Umamaheswari, A.; Elumalai, S. Global health concern of cyanotoxins in surface water and its various detection methods. Curr. Bot. 2020, 11, 65–74. [Google Scholar] [CrossRef]
- Ahari, H.; Nowruzi, B.; Anvar, A.A.; Porzani, S.J. The toxicity testing of cyanobacterial toxins in vivo and in vitro by mouse bioassay: A review. Mini Rev. Med. Chem. 2022, 22, 1131–1151. [Google Scholar]
- Kaushik, R.; Balasubramanian, R. Methods and approaches used for detection of cyanotoxins in environmental samples: A review. Crit. Rev. Environ. Sci. Technol. 2013, 43, 1349–1383. [Google Scholar]
- Campos, A.; Vasconcelos, V. Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar]
- Massey, I.Y.; Wu, P.; Wei, J.; Luo, J.; Ding, P.; Wei, H.; Yang, F. A mini-review on detection methods of microcystins. Toxins 2020, 12, 641. [Google Scholar] [PubMed]
- Froscio, S.M.; Humpage, A.R.; Burcham, P.C.; Falconer, I.R. Cell-free protein synthesis inhibition assay for the cyanobacterial toxin cylindrospermopsin. Environ. Toxicol. Int. J. 2001, 16, 408–412. [Google Scholar]
- Devic, E.; Li, D.; Dauta, A.; Henriksen, P.; Codd, G.A.; Marty, J.L.; Fournier, D. Detection of anatoxin-a (s) in environmental samples of cyanobacteria by using a biosensor with engineered acetylcholinesterases. Appl. Environ. Microbiol. 2002, 68, 4102–4106. [Google Scholar] [PubMed]
- Llewellyn, L.E.; Doyle, J.; Negri, A.P. A high-throughput, microtiter plate assay for paralytic shellfish poisons using the saxitoxin-specific receptor, saxiphilin. Anal. Biochem. 1998, 261, 51–56. [Google Scholar] [PubMed]
- Aráoz, R.; Nghiêm, H.O.; Rippka, R.; Palibroda, N.; de Marsac, N.T.; Herdman, M. Neurotoxins in axenic oscillatorian cyanobacteria: Coexistence of anatoxin-a and homoanatoxin-a determined by ligand-binding assay and GC/MS. Microbiology 2005, 151, 1263–1273. [Google Scholar]
- Zhang, H.; Li, B.; Liu, Y.; Chuan, H.; Liu, Y.; Xie, P. Immunoassay technology: Research progress in microcystin-LR detection in water samples. J. Hazard. Mater. 2022, 424, 127406. [Google Scholar]
- Fischer, W.J.; Garthwaite, I.; Miles, C.O.; Ross, K.M.; Aggen, J.B.; Chamberlin, A.R.; Towers, N.R.; Dietrich, D.R. Congener-independent immunoassay for microcystins and nodularins. Environ. Sci. Technol. 2001, 35, 4849–4856. [Google Scholar]
- Mikhailov, A.; Härmälä-Braskén, A.S.; Polosukhina, E.; Hanski, A.; Wahlsten, M.; Sivonen, K.; Eriksson, J.E. Production and specificity of monoclonal antibodies against nodularin conjugated through N-methyldehydrobutyrine. Toxicon 2001, 39, 1453–1459. [Google Scholar]
- do Carmo Bittencourt-Oliveira, M.; Piccin-Santos, V.; Kujbida, P.; do Nascimento Moura, A. Cylindrospermopsin in water supply reservoirs in Brazil determined by immunochemical and molecular methods. J. Water Resour. Prot. 2011, 3, 349. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, H.; Ma, L.; Liu, D.; Wang, Z. A label-free electrochemical impedance aptasensor for cylindrospermopsin detection based on thionine–graphene nanocomposites. Analyst 2015, 140, 5570–5577. [Google Scholar] [CrossRef]
- Cevallos-Cedeño, R.E.; Quiñones-Reyes, G.; Agulló, C.; Abad-Somovilla, A.; Abad-Fuentes, A.; Mercader, J.V. Rapid Immunochemical Methods for Anatoxin-a Monitoring in Environmental Water Samples. Anal. Chem. 2022, 94, 10857–10864. [Google Scholar] [CrossRef]
- Micheli, L.; Di Stefano, S.; Moscone, D.; Palleschi, G.; Marini, S.; Coletta, M.; Draisci, R.; Delli Quadri, F. Production of antibodies and development of highly sensitive formats of enzyme immunoassay for saxitoxin analysis. Anal. Bioanal. Chem. 2002, 373, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Rosso, L.; Sedan, D.; Giannuzzi, L.; Andrinolo, D.; Volmer, D.A. Seven new microcystin variants discovered from a native Microcystis aeruginosa strain–unambiguous assignment of product ions by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2015, 29, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xie, P.; Liang, G.; Chen, J.; Lei, H. Simultaneous determination of microcystin-LR and its glutathione conjugate in fish tissues by liquid chromatography–tandem mass spectrometry. J. Chromatogr. B 2008, 862, 43–50. [Google Scholar] [CrossRef]
- Gallo, P.; Fabbrocino, S.; Cerulo, M.G.; Ferranti, P.; Bruno, M.; Serpe, L. Determination of cylindrospermopsin in freshwaters and fish tissue by liquid chromatography coupled to electrospray ion trap mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 3279–3284. [Google Scholar] [CrossRef]
- Haddad, S.P.; Bobbitt, J.M.; Taylor, R.B.; Lovin, L.M.; Conkle, J.L.; Chambliss, C.K.; Brooks, B.W. Determination of microcystins, nodularin, anatoxin-a, cylindrospermopsin, and saxitoxin in water and fish tissue using isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1599, 66–74. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M. Tumor promoters-microcystin-LR, nodularin and TNF-α and human cancer development. Anti-Cancer Agents Med. Chem. 2011, 11, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, X.; Sun, Q.; Ren, X.; Zhong, J.; Zhou, L.; Zhang, H.; Li, G.; Liu, Y.; Liu, J.; et al. Long term exposure of saxitoxin induced cognitive deficits and YAP1 cytoplasmic retention. Ecotoxicol. Environ. Saf. 2023, 253, 114645. [Google Scholar] [CrossRef]
- Newell, M.E.; Adhikari, S.; Halden, R.U. Systematic and state of the science review of the role of environmental factors in Amyotophic Lateral Sclerosis (ALS) or Lou Gehrig’s Disease. Sci. Total Environ. 2021, 817, 152504. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Banack, S.A.; Bishop, S.L.; Metcalf, J.S.; Murch, S.J.; Davis, D.A.; Stommel, E.W.; Karlsson, O.; Brittebo, E.B.; Chatziefthimiou, A.D.; et al. Is exposure to BMAA a risk factor for neurodegenerative diseases? A response to a critical review of the BMAA hypothesis. Neurotox. Res. 2021, 39, 81–106. [Google Scholar]
- Bishop, S.L.; Murch, S.J. A systematic review of analytical methods for the detection and quantification of β-N-methylamino-l-alanine (BMAA). Analyst 2020, 145, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Euerby, M.R.; Partridge, L.Z.; Nunn, P.B. Resolution of neuroactive non-protein amino acid enantiomers by high-performance liquid chromatography utilizing pre-column derivatisation with o-phthaldialdehyde-chiral thiols. Application to 2-amino-ω-phosphoalkanoic acid homologues and α-amino-β-N-methylaminopropanoic acid (β-methylaminoalanine). J. Chromatogr. 1989, 469, 412–419. [Google Scholar] [PubMed]
- Metcalf, J.S.; Lobner, D.; Banack, S.A.; Cox, G.A.; Nunn, P.B.; Wyatt, P.B.; Cox, P.A. Analysis of BMAA enantiomers in cycads, cyanobacteria, and mammals: In vivo formation and toxicity of D-BMAA. Amino Acids 2017, 49, 1427–1439. [Google Scholar] [CrossRef]
- Zurita, J.; Zguna, N.; Andrýs, R.; Strzelczak, A.; Jiang, L.; Thorsen, G.; Ilag, L.L. Chiral analysis of β-methylaminoalanine (BMAA) enantiomers after (+)-1-(9-fluorenyl)-ethyl chloroformate (FLEC) derivatization and LC-MS/MS. Anal. Meth. 2019, 11, 432–442. [Google Scholar] [CrossRef]
- Vega, A.; Bell, E.A. α-amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Vega, A.; Bell, E.A.; Nunn, P.B. The preparation of L- and D-α-amino-β-methylaminopropionic acid and the identification of the compound isolated from Cycas circinalis as the L-isomer. Phytochemistry 1968, 7, 1885–1887. [Google Scholar] [CrossRef]
- Polsky, F.I.; Nunn, P.B.; Bell, E.A. Distribution and toxicity of α-amino-β-methylaminopropionic acid. Fed. Proc. 1972, 31, 1473–1475. [Google Scholar]
- Friedman, M. Origin, microbiology, nutrition and pharmacology of D-amino acids. Chem. Biodivers. 2010, 7, 1491–1530. [Google Scholar] [CrossRef]
- Baumgart, F.; Rodriguez-Crespo, I. D-amino acids in the brain: The biochemistry of serine racemase. FEBS J. 2008, 275, 3538–3548. [Google Scholar] [CrossRef]
- Kim, P.M.; Duan, X.; Huang, A.S.; Liu, C.Y.; Ming, G.-L.; Song, H.; Snyder, S.H. Aspartate racemase, generating neuronal D-aspartate, regulates adult neurogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3175–3179. [Google Scholar] [CrossRef]
- Krashia, P.; Ledonne, A.; Nobili, A.; Cordella, A.; Errico, F.; Usiello, A.; D’Amelio, M.; Mercuri, N.B.; Guatteo, E.; Carunchio, I. Persistent elevation of D-aspartate enhances NMDA receptor-mediated responses in mouse substantia nigra pars compacta dopamine neurons. Neuropharmacology 2016, 103, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.A.; Watson, A.A.; Nash, R.J. Non-protein amino acids: A review of the biosynthesis and taxonomic significance. Nat. Prod. Comm. 2008, 3, 93–110. [Google Scholar] [CrossRef]
- Tymm, F.J.M.; Bishop, S.L.; Murch, S.J. A single laboratory validation for the analysis of underivatized β-N-methylamino-l-alanine (BMAA). Neurotox. Res. 2019, 39, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.B.; Baker, T.C.; Murch, S.J.; Brown, P.N. Determination of β-N-methylamino-l-alanine, N-(2-aminoethyl)glycine and 2,4-diaminobutyric acid in food products containing cyanobacteria by ultra-performance liquid chromatography and tandem mass spectrometry: Single laboratory validation. J. AOAC Int. 2015, 98, 1559–1565. [Google Scholar] [CrossRef]
- Banack, S.A. Second laboratory validation of β-N-methylamino-l-alanine, N-(2-aminoethyl)glycine and 2,4-diaminobutyric acid by ultra-performance liquid chromatography and tandem mass spectrometry. Neurotox. Res. 2020, 39, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Spacil, Z.; Eriksson, J.; Rydberg, S.; Rasmussen, U.; Ilag, L.L.; Bergman, B. Analytical protocol for identification of BMAA and DAB in biological samples. Analyst 2010, 135, 127–132. [Google Scholar] [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and bacteria isolated from a diatom culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-l-alanine (BMAA) and its isomers by freshwater diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Wessel, R.A.; Lester, M.; Pim, J.G.; Cassani, J.R.; Cox, P.A. Toxin analysis of freshwater cyanobacterial and marine harmful algal blooms on the west coast of florida and implications for estuarine environments. Neurotox. Res. 2021, 39, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Yan, Y.; Qiu, J.; Yan, G.; Zhao, P.; Li, M.; Ji, Y.; Wang, G.; Meng, F.; Li, Y.; et al. Putative biosynthesis mechanism of the neurotoxin β-N-methylamino-l-alanine in marine diatoms based on a transcriptomics approach. J. Hazard. Mater. 2023, 441, 129953. [Google Scholar] [CrossRef]
- Schneider, T.; Simpson, C.; Desai, P.; Tucker, M.; Lobner, D. Neurotoxicity of isomers of the environmental toxin L-BMAA. Toxicon 2020, 184, 175–179. [Google Scholar] [CrossRef]
- Ward, T.J.; Ward, K.D. Chiral separations: Fundamental review. Anal. Chem. 2010, 82, 4712–4722. [Google Scholar] [CrossRef] [PubMed]
- Visser, W.F.; Verhoeven-Duif, N.M.; Ophoff, R.; Bakker, S.; Klomp, L.W.; Berger, R.; de Koning, T.J. A sensitive and simple ultra-high-performance-liquid chromatography-tandem mass spectrometric method for the quantification of D-amino acids in body fluids. J. Chromatogr. A 2011, 1218, 7130–7136. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Nunn, P.B.; Codd, G.A. Environmental distribution of the neurotoxin L-BMAA in Paenibacillus species. Toxicol. Res. 2019, 8, 781–783. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Gobler, C.J.; Hansen, P.J. Dynamic CO2 and pH levels in coastal, estuarine, and inland waters: Theoretical and observed effects on harmful algal blooms. Harmful Algae 2020, 91, 101594. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef]
- Davis, D.A.; Cox, P.A.; Banack, S.A.; Lecusay, P.D.; Garamszegi, S.P.; Hagan, M.J.; Powell, J.T.; Metcalf, J.S.; Palmour, R.M.; Beierschmitt, A.; et al. L-serine reduces spinal cord pathology in a vervet model of preclinical ALS/MND. J. Neuropath. Exp. Neurol. 2020, 79, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Kotut, K.; Krienitz, L.; Codd, G.A. Amino acid neurotoxins in feathers of the Lesser Flamingo, Phoeniconaias minor. Chemosphere 2013, 90, 835–839. [Google Scholar] [CrossRef]

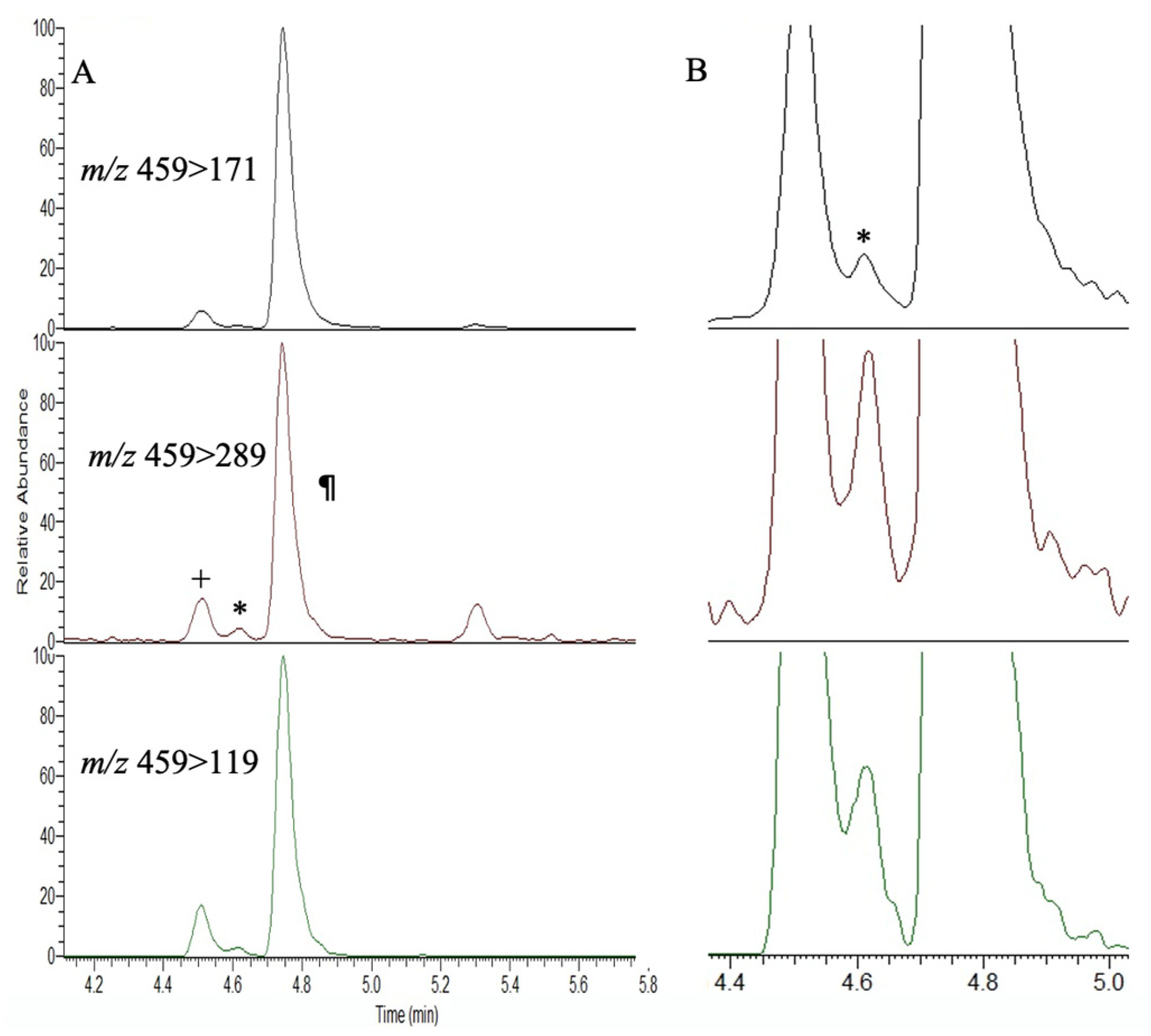
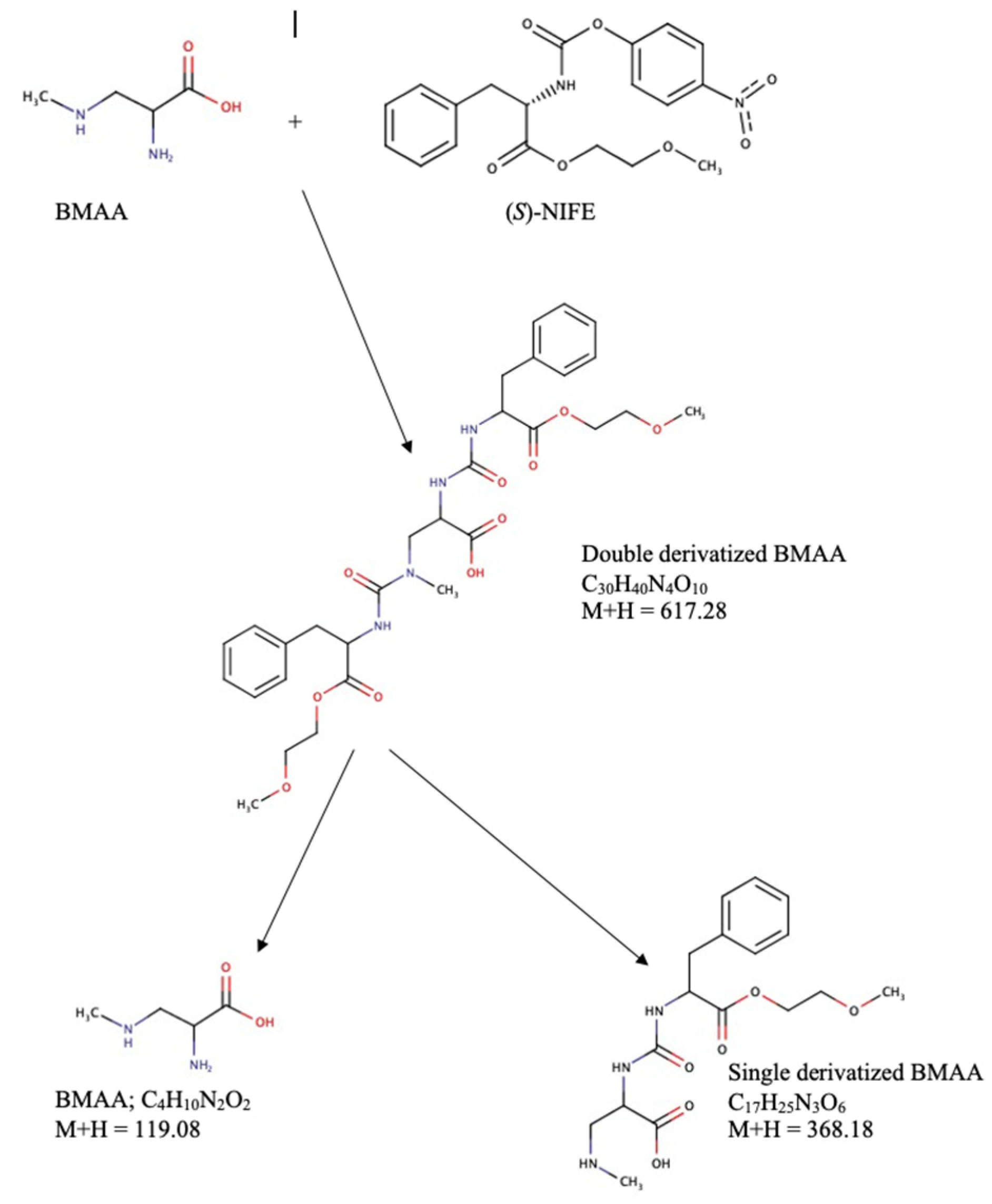

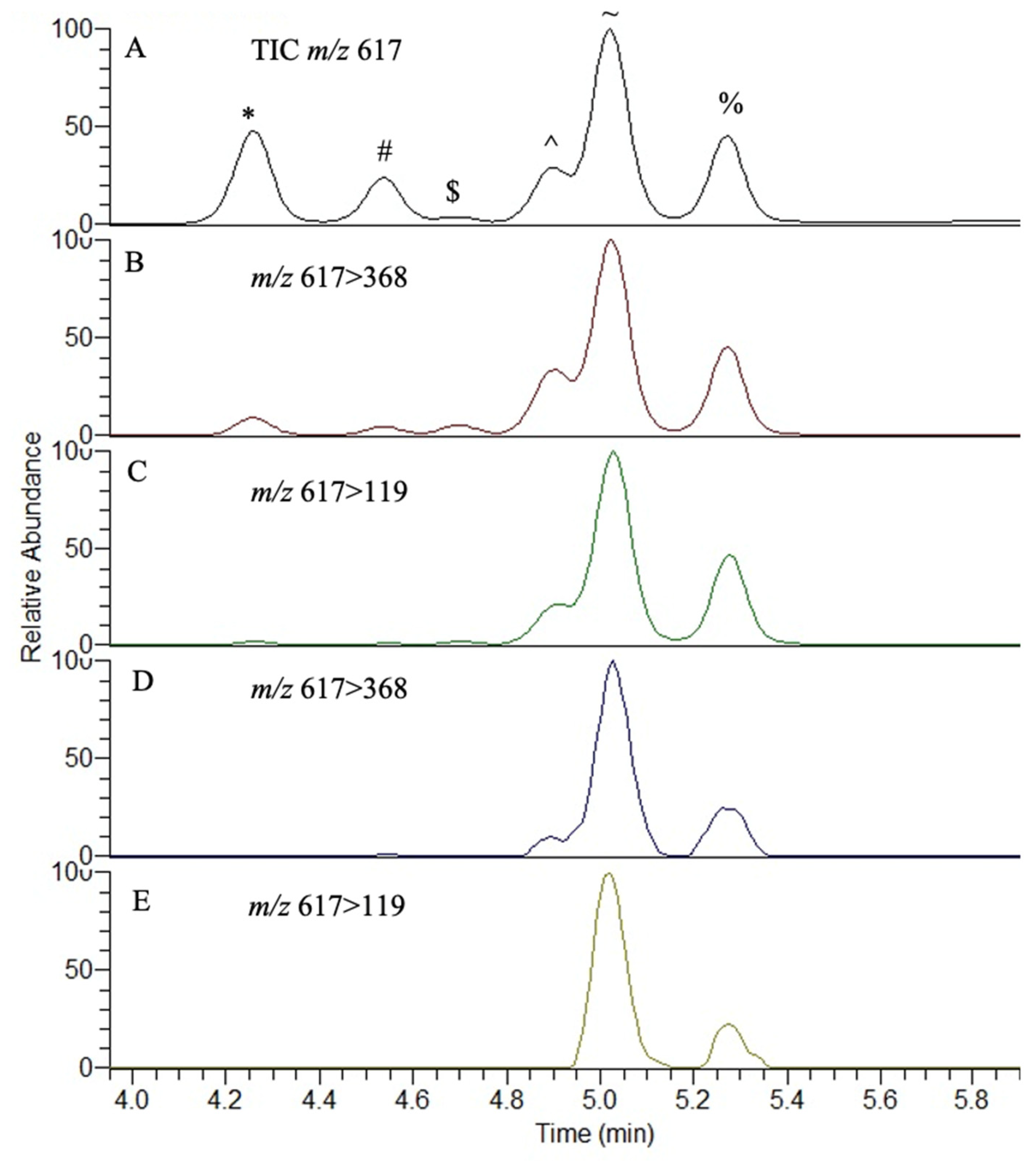


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L-BMAA | D-BMAA | AEG | L-DAB | D-DAB | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Conc (μg/mL) | ng on Column | RT %CV | Area %CV | RT %CV | Area %CV | RT %CV | Area %CV | RT %CV | Area %CV | RT %CV | Area %CV |
| 0.1 | 0.5 | 0.18 | 12.32 | 0.16 | 17.03 | 0.27 | 18.73 | ND | ND | ND | ND |
| 1 | 5 | 0.40 | 22.93 | 0.20 | 12.02 | 0.25 | 10.09 | 0.24 | 11.65 | 0.25 | 9.42 |
| 10 | 50 | 0.26 | 25.70 | 0.25 | 29.06 | 0.24 | 14.14 | 0.26 | 12.68 | 0.23 | 10.66 |
| 100 | 500 | 0.21 | 13.32 | 0.22 | 8.58 | 0.23 | 7.80 | 0.23 | 7.96 | 0.26 | 5.73 |
| L-BMAA | D-BMAA | AEG | L-DAB | D-DAB | |
|---|---|---|---|---|---|
| Conc (μg/mL) | RT range (min) | RT range (min) | RT range (min) | RT range (min) | RT range (min) |
| 0.1 | 5.098–5.127 | 5.268–5.292 | 4.822–4.856 | 4.211–4.240 | 4.482–4.499 |
| 1 | 5.096–5.147 | 5.269–5.298 | 4.824–4.853 | 4.223–4.237 | 4.479–4.507 |
| 10 | 5.062–5.092 | 5.265–5.298 | 4.827–4.856 | 4.213–4.242 | 4.478–4.501 |
| 100 | 5.028–5.058 | 5.255–5.268 | 4.826–4.851 | 4.211–4.239 | 4.472–4.499 |
| LOD (μg/mL) | 0.0013 | 0.0025 | 0.0031 | 0.0025 | 0.0033 |
| LOQ (μg/mL) | 0.0042 | 0.0085 | 0.0103 | 0.00846 | 0.0111 |
| Enantiomer | Linearity (μg/mL) | Intra-Day Precision (%) | Inter-Day Precision (%) |
|---|---|---|---|
| L-BMAA | 0.1–100 | 87–112 | 85–117 |
| D-BMAA | 0.1–100 | 95–104 | 88–117 |
| AEG | 0.1–100 | 95–104 | 91–112 |
| L-DAB | 0.1–100 | 90–109 | 86–112 |
| D-DAB | 0.1–100 | 88–111 | 90–110 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metcalf, J.S.; Banack, S.A.; Wyatt, P.B.; Nunn, P.B.; Cox, P.A. A Direct Analysis of β-N-methylamino-l-alanine Enantiomers and Isomers and Its Application to Cyanobacteria and Marine Mollusks. Toxins 2023, 15, 639. https://doi.org/10.3390/toxins15110639
Metcalf JS, Banack SA, Wyatt PB, Nunn PB, Cox PA. A Direct Analysis of β-N-methylamino-l-alanine Enantiomers and Isomers and Its Application to Cyanobacteria and Marine Mollusks. Toxins. 2023; 15(11):639. https://doi.org/10.3390/toxins15110639
Chicago/Turabian StyleMetcalf, James S., Sandra Anne Banack, Peter B. Wyatt, Peter B. Nunn, and Paul A. Cox. 2023. "A Direct Analysis of β-N-methylamino-l-alanine Enantiomers and Isomers and Its Application to Cyanobacteria and Marine Mollusks" Toxins 15, no. 11: 639. https://doi.org/10.3390/toxins15110639
APA StyleMetcalf, J. S., Banack, S. A., Wyatt, P. B., Nunn, P. B., & Cox, P. A. (2023). A Direct Analysis of β-N-methylamino-l-alanine Enantiomers and Isomers and Its Application to Cyanobacteria and Marine Mollusks. Toxins, 15(11), 639. https://doi.org/10.3390/toxins15110639