Oxidative Stress and DNA Damage in Pagrus major by the Dinoflagellate Karenia mikimotoi


 and
and
Abstract
:1. Introduction
2. Results
2.1. Acute Toxicity
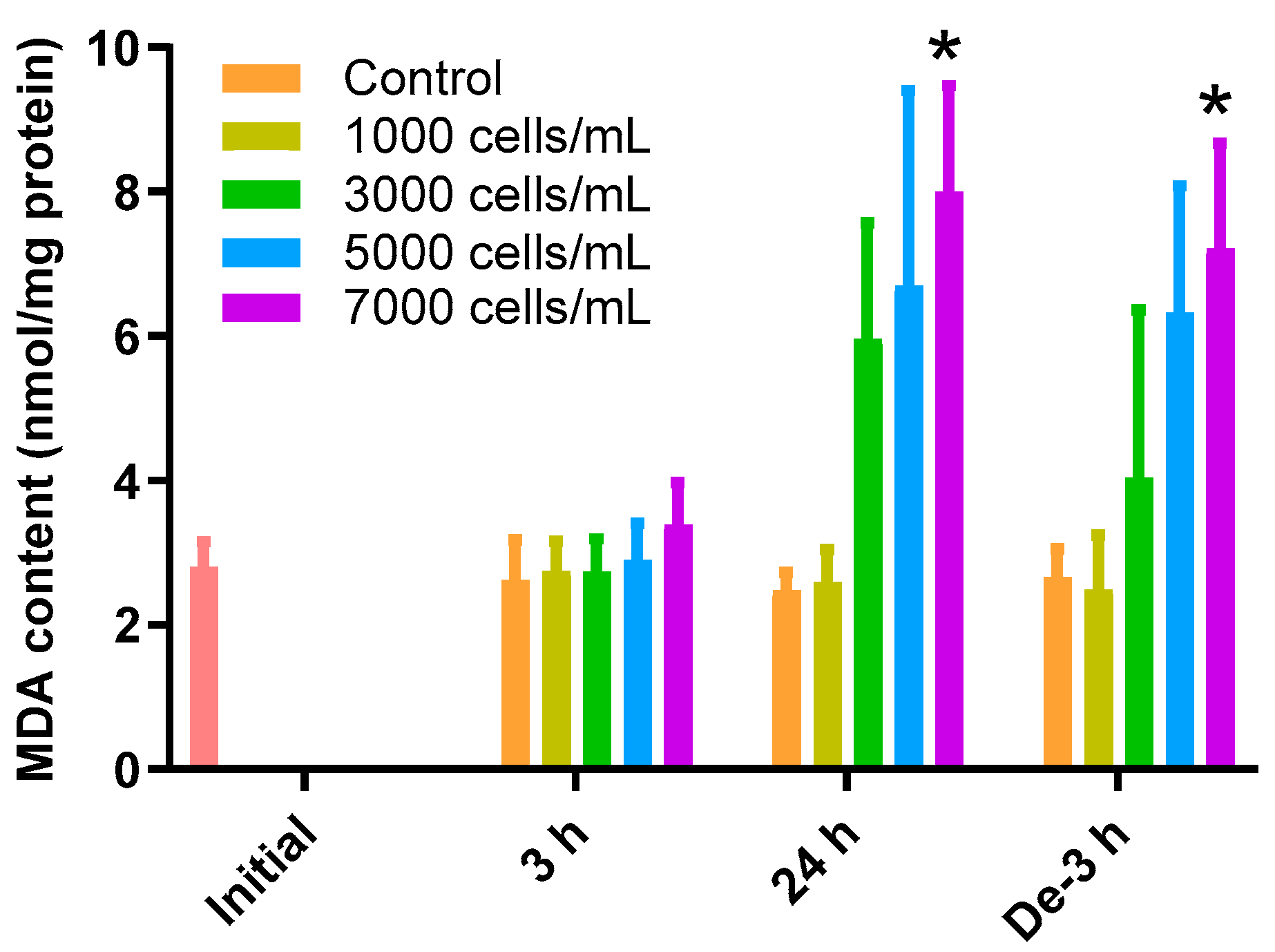
2.2. MDA Content as an Oxidative Index
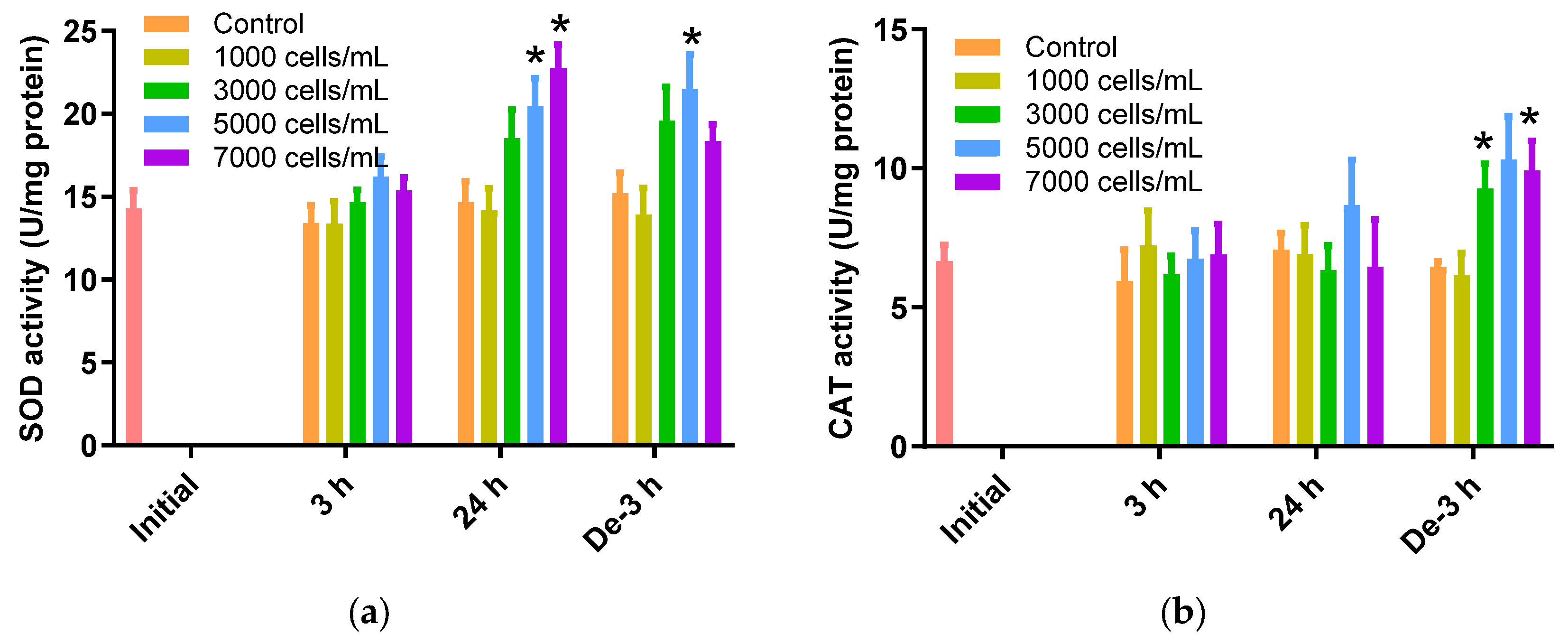
2.3. ROS Scavenging Enzymes Activities
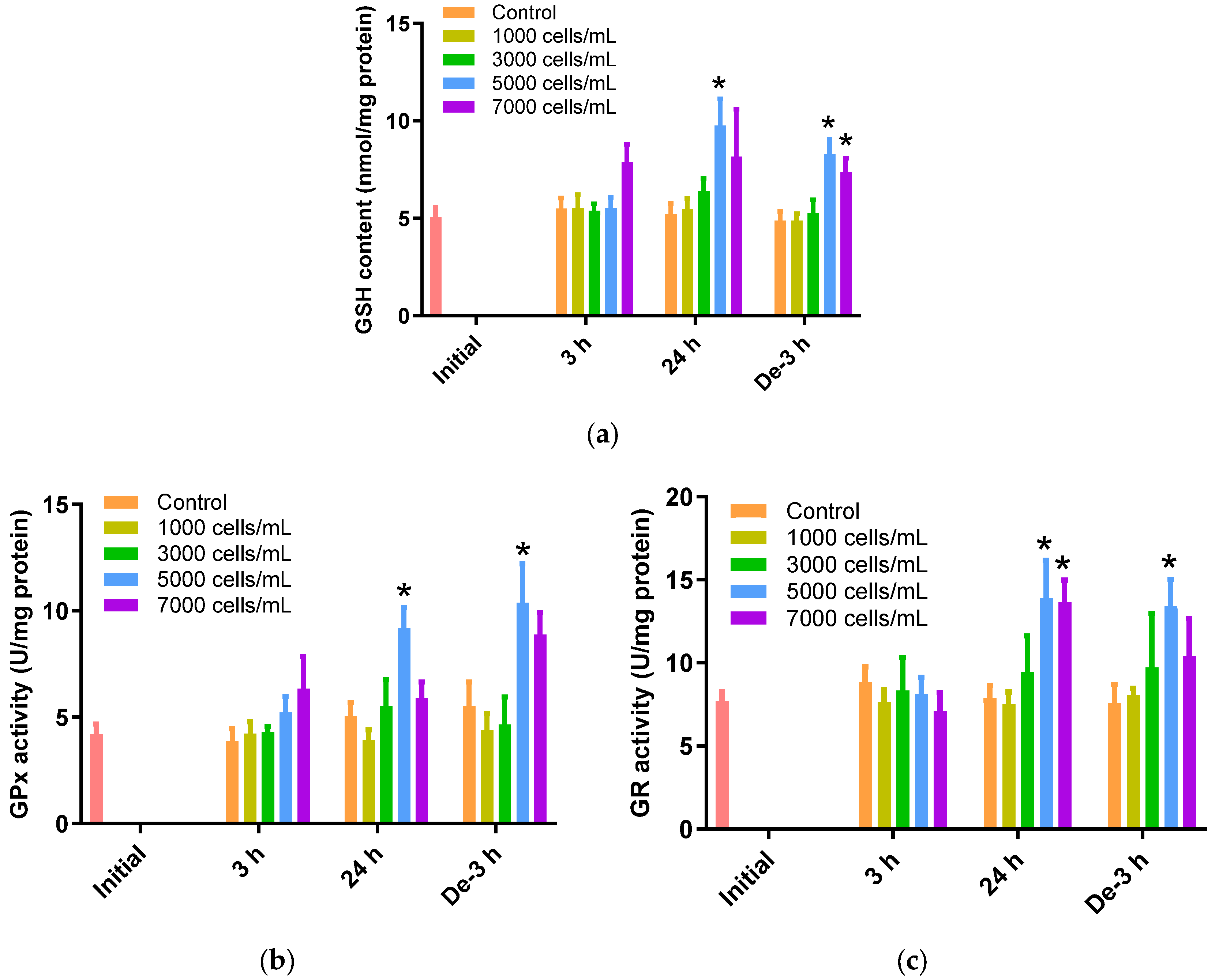
2.4. Glutathione Dependent Metabolic Pathway
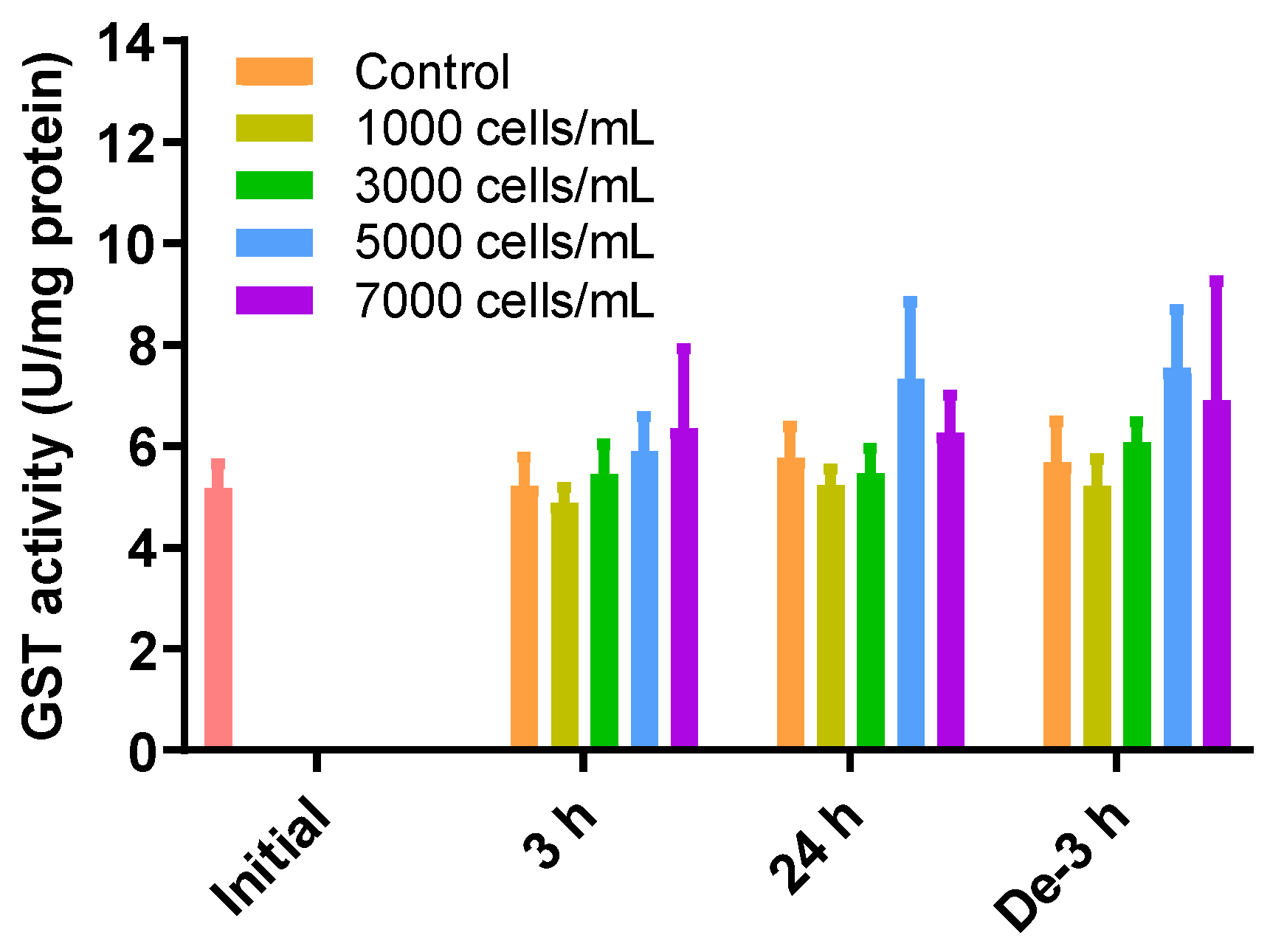
2.5. GST Activity as a Detoxification Enzyme
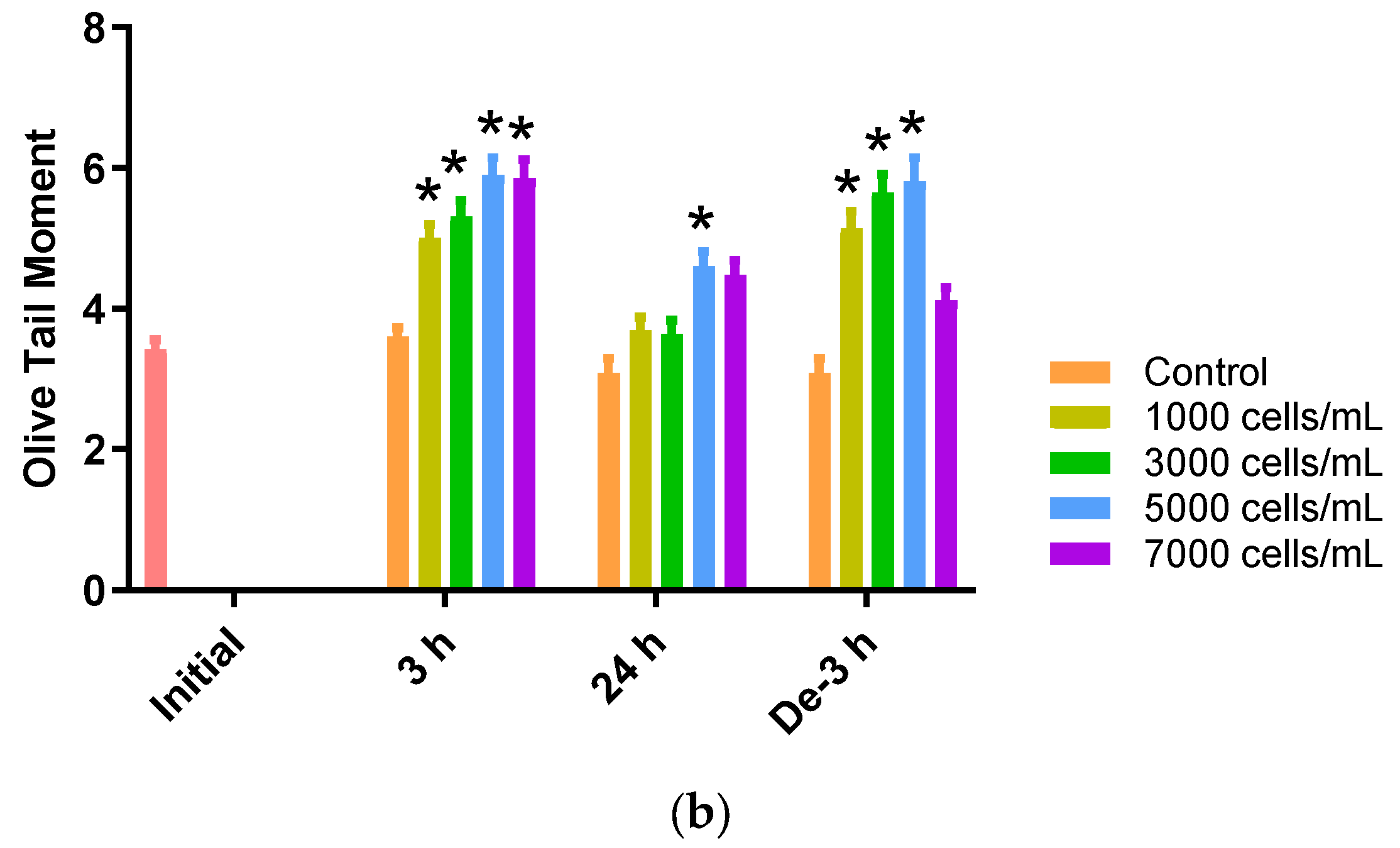
2.6. DNA Damage in Blood Samples of P. major
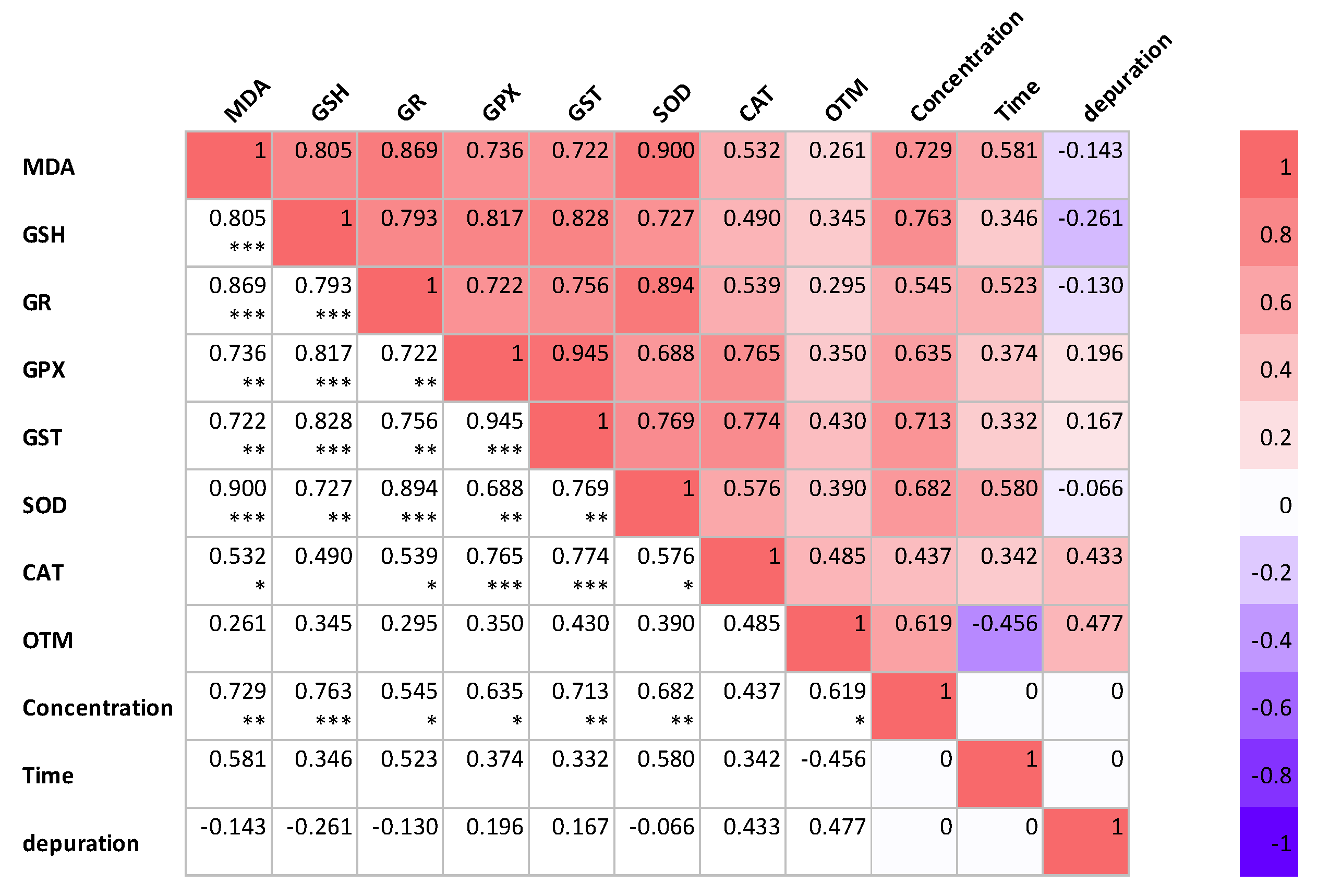
2.7. Correlation Analysis
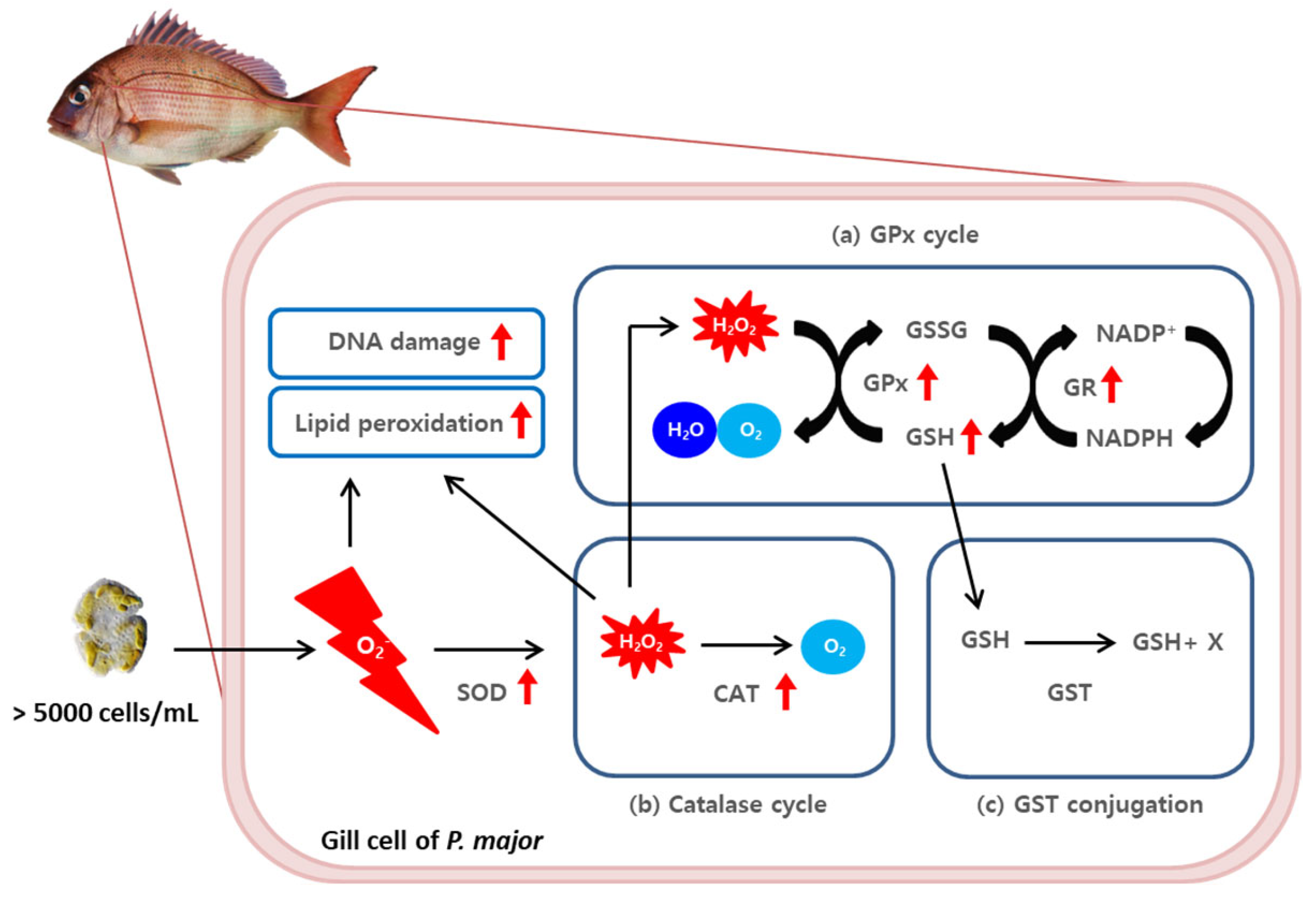
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Fish and K. mikimotoi Cultivation Conditions
5.2. Exposure to K. mikimotoi and Sample Preparation
5.3. Measurements of Lipid Peroxidation Marker
5.4. Measurements of Antioxidant Molecule
5.5. Enzyme Activity Analysis
5.6. Comet Assay
5.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hallegraeff, G. Harmful Algal Blooms: A Global Overview. In Manual on Harmful Marine Microalgae, 2nd; Monographs on Oceanographic Methodology; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; IOC-UNESCO: Paris, France, 2003; pp. 25–49. [Google Scholar] [CrossRef]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Backer, L.C.; Fleming, L.E. Impacts of Climate Variability and Future Climate Change on Harmful Algal Blooms and Human Health. Environ. Health 2008, 7, S4. [Google Scholar] [CrossRef] [PubMed]
- McLeod, D.J.; Hallegraeff, G.M.; Hosie, G.W.; Richardson, A.J. Climate-Driven Range Expansion of the Red-Tide Dinoflagellate Noctiluca scintillans into the Southern Ocean. J. Plankton Res. 2012, 34, 332–337. [Google Scholar] [CrossRef]
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Imai, I.; Yamaguchi, M.; Hori, Y. Eutrophication and Occurrences of Harmful Algal Blooms in the Seto Inland Sea, Japan. Plankton Benthos Res. 2006, 1, 71–84. [Google Scholar] [CrossRef]
- Yang, Z.; Hodgkiss, I. Hong Kong’s Worst “Red Tide”—Causative Factors Reflected in a Phytoplankton Study at Port Shelter Station in 1998. Harmful Algae 2004, 3, 149–161. [Google Scholar] [CrossRef]
- Cho, K.; Ueno, M.; Liang, Y.; Kim, D.; Oda, T. Generation of Reactive Oxygen Species (ROS) by Harmful Algal Bloom (HAB)-Forming Phytoplankton and Their Potential Impact on Surrounding Living Organisms. Antioxidants 2022, 11, 206. [Google Scholar] [CrossRef]
- O’Boyle, S.; McDermott, G.; Silke, J.; Cusack, C. Potential Impact of an Exceptional Bloom of Karenia mikimotoi on Dissolved Oxygen Levels in Waters off Western Ireland. Harmful Algae 2016, 53, 77–85. [Google Scholar] [CrossRef]
- Barnes, M.K.; Tilstone, G.H.; Smyth, T.J.; Widdicombe, C.E.; Gloël, J.; Robinson, C.; Kaiser, J.; Suggett, D.J. Drivers and Effects of Karenia mikimotoi Blooms in the Western English Channel. Prog. Oceanogr. 2015, 137, 456–469. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine Algal Toxins: Origins, Health Effects, and Their Increased Occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Byun, C.K.; Jo, J.-Y. Seed Production of Red Sea-Bream, Pagrus major. Korean J. Fish. Aquat. Sci. 1982, 15, 161–170. [Google Scholar]
- Jang, Y.L.; Lee, H.J.; Moon, H.-B.; Lee, W.-C.; Kim, H.C.; Kim, G.B. Marine Environmental Characteristics in the Coastal Area Surrounding Tongyeong Cage-Fish Farms. J. Korean Soc. Mar. Environ. Energy 2015, 18, 74–80. Available online: https://www.dbpia.co.kr/journal/articleDetail?nodeId=NODE06298479&language=ko_KR&hasTopBanner=true#a (accessed on 16 January 2023).
- Lee, D.C.; Won, K.; Park, M.A.; Sung, C.H.; Jung, S.H. An Analysis of Mass Mortalities in Aquaculture Fish Farms on the Southern Coast in Korea. Ocean Policy Res. 2018, 33, 1–16. [Google Scholar] [CrossRef]
- Lee, C.-K.; Park, T.-G.; Park, Y.-T.; Lim, W.-A. Monitoring and Trends in Harmful Algal Blooms and Red Tides in Korean Coastal Waters, with Emphasis on Cochlodinium polykrikoides. Harmful Algae 2013, 30, S3–S14. [Google Scholar] [CrossRef]
- Sakamoto, S.; Lim, W.E.; Lu, D.; Dai, X.; Orlova, T.; Iwataki, M. Harmful Algal Blooms and Associated Fisheries Damage in East Asia: Current Status and Trends in China, Japan, Korea, and Russia. Harmful Algae 2021, 102, 101787. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Campbell, L.; Bresnan, E. Karenia: The Biology and Ecology of a Toxic Genus. Harmful Algae 2012, 14, 156–178. [Google Scholar] [CrossRef] [PubMed]
- Siswanto, E.; Ishizaka, J.; Tripathy, S.C.; Miyamura, K. Detection of Harmful Algal Blooms of Karenia mikimotoi Using MODIS Measurements: A Case Study of Seto-Inland Sea, Japan. Remote Sens. Environ. 2013, 129, 185–196. [Google Scholar] [CrossRef]
- Kakumu, A.; Morita, K.; Shimada, H.; Yamaguchi, A.; Imai, I. First Detection of the Noxious Red Tide Dinoflagellate Karenia mikimotoi and Bloom Dynamics in 2015 and 2016 in Hakodate Bay, Hokkaido, Northern Japan. Bull. Plankton Soc. Jpn. 2018, 65, 1–11. [Google Scholar]
- Wang, J.; Wu, J. Occurrence and Potential Risks of Harmful Algal Blooms in the East China Sea. Sci. Total Environ. 2009, 407, 4012–4021. [Google Scholar] [CrossRef]
- Lu, S.; Hodgkiss, I. Harmful algal bloom causative collected from Hong Kong waters. In Asian Pacific Phycology in the 21st Century: Prospects and Challenges; Developments in Hydrobiology; Ang, P.O., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 173, pp. 231–238. [Google Scholar] [CrossRef]
- Silke, J.; O’Beirn, F.; Cronin, M. Karenia mikimotoi: An Exceptional Dinoflagellate Bloom in Western Irish Waters, Summer 2005. In Marine Environment and Health Series 21; Marine Institute: Galway, Ireland, 2005. [Google Scholar]
- Kurekin, A.; Miller, P.; Van der Woerd, H. Satellite Discrimination of Karenia mikimotoi and Phaeocystis Harmful Algal Blooms in European Coastal Waters: Merged Classification of Ocean Colour Data. Harmful Algae 2014, 31, 163–176. [Google Scholar] [CrossRef]
- Kim, H. An Overview on the Occurrences of Harmful Algal Blooms (HABs) and Mitigation Strategies in Korean Coastal Waters. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; TERRAPUB and Nagasaki University: Nagasaki, Japan, 2010; pp. 121–131. [Google Scholar]
- Steidinger, K.; Garccés, E. Importance of Life Cycles in the Ecology of Harmful Microalgae. In Ecology of Harmful Algae; Springer: Berlin/Heidelberg, Germany, 2006; pp. 37–49. [Google Scholar] [CrossRef]
- Lei, Q.-Y.; Lü, S.-H. Molecular Ecological Responses of Dinoflagellate, Karenia mikimotoi to Environmental Nitrate Stress. Mar. Pollut. Bull. 2011, 62, 2692–2699. [Google Scholar] [CrossRef]
- Lei, Q.-Y.; Lu, S.-H. Molecular Ecological Responses of the Dinoflagellate Karenia mikimotoi to Phosphate Stress. Harmful Algae 2011, 12, 39–45. [Google Scholar] [CrossRef]
- Kim, D.; Wencheng, L.; Matsuyama, Y.; Cho, K.; Yamasaki, Y.; Takeshita, S.; Yamaguchi, K.; Oda, T. Extremely High Level of Reactive Oxygen Species (ROS) Production in a Newly Isolated Strain of the Dinoflagellate Karenia mikimotoi. Eur. J. Phycol. 2019, 54, 632–640. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Kim, D.I.; Matsuyama, Y.; Oda, T.; Honjo, T. Production of Superoxide Anion and Hydrogen Peroxide by the Red Tide Dinoflagellate Karenia mikimotoi. J. Biosci. Bioeng. 2004, 97, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, J.; Zhuang, Y.; Yu, W.; Liu, G. Reduced Fitness and Elevated Oxidative Stress in the Marine Copepod Tigriopus japonicus Exposed to the Toxic Dinoflagellate Karenia mikimotoi. Antioxidants 2022, 11, 2299. [Google Scholar] [CrossRef]
- Li, X.; Yan, T.; Yu, R.; Zhou, M. A Review of Karenia mikimotoi: Bloom Events, Physiology, Toxicity and Toxic Mechanism. Harmful Algae 2019, 90, 101702. [Google Scholar] [CrossRef]
- Niu, X.; Xu, S.; Yang, Q.; Xu, X.; Zheng, M.; Li, X.; Guan, W. Toxic Effects of the Dinoflagellate Karenia mikimotoi on Zebrafish (Danio rerio) Larval Behavior. Harmful Algae 2021, 103, 101996. [Google Scholar] [CrossRef]
- Zhang, P.P.; Song, X.X.; Zhang, Y.; Zhu, J.A.; Shen, H.H.; Yu, Z.M. Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism. Toxics 2022, 10, 105. [Google Scholar] [CrossRef]
- Botes, L.; Smit, A.; Cook, P. The Potential Threat of Algal Blooms to the Abalone (Haliotis midae) Mariculture Industry Situated Around the South African Coast. Harmful Algae 2003, 2, 247–259. [Google Scholar] [CrossRef]
- Lin, J.; Yan, T.; Zhang, Q.; Wang, Y.; Liu, Q.; Zhou, M. Effects of Karenia mikimotoi Blooms on Antioxidant Enzymes in Gastropod Abalone, Haliotis discus hannai. Mar. Sci. 2016, 40, 17–22. [Google Scholar]
- Davidson, K.; Miller, P.; Wilding, T.A.; Shutler, J.; Bresnan, E.; Kennington, K.; Swan, S. A Large and Prolonged Bloom of Karenia mikimotoi in Scottish Waters in 2006. Harmful Algae 2009, 8, 349–361. [Google Scholar] [CrossRef]
- Zou, Y.; Yamasaki, Y.; Matsuyama, Y.; Yamaguchi, K.; Honjo, T.; Oda, T. Possible Involvement of Hemolytic Activity in the Contact-Dependent Lethal Effects of the Dinoflagellate Karenia mikimotoi on the Rotifer Brachionus plicatilis. Harmful Algae 2010, 9, 367–373. [Google Scholar] [CrossRef]
- Chen, Y.; Yan, T.; Yu, R.; Zhou, M. Toxic Effects of Karenia mikimotoi Extracts on Mammalian Cells. Chinese J. Oceanol. Limnol. 2011, 29, 860–868. [Google Scholar] [CrossRef]
- Helm, M.M.; Hepper, B.T.; Spencer, B.E.; Walne, B.R. Lugworm Mortalities and a Bloom of Gyrodinium aureolum Hulburt in Eastern Irish Sea, Autumn 1971. J. Mar. Biol. Assoc. UK 1974, 54, 857–869. [Google Scholar] [CrossRef]
- Jones, K.J.; Ayres, P.; Bullock, A.M.; Roberts, R.J.; Tett, P. A Red Tide of Gyrodinium aureolum in Sea Lochs of the Firth of Clyde and Associated Mortality of Pond-Reared Salmon. J. Mar. Biol. Assoc. UK 1982, 62, 771–782. [Google Scholar] [CrossRef]
- Mooney, B.D.; De Salas, M.; Hallegraeff, G.M.; Place, A.R. Survey for Karlotoxin Production in 15 Species of Gymnodinioid Dinoflagellates (Kareniaceae, Dinophyta). J. Phycol. 2009, 45, 164–175. [Google Scholar] [CrossRef]
- Ou, X.Y. Histopathological Effect of Karenia mikimotoi Bloom on Gills of Common Farmed Fishes in the East China Sea. Master’s Thesis, Ji Nan University, Guangzhou, China, 2006. [Google Scholar]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.P.; Liu, W.; Au, D.W.; Anderson, D.M.; Wu, R.S. Antioxidant Responses and Lipid Peroxidation in Gills and Erythrocytes of Fish (Rhabdosarga sarba) upon Exposure to Chattonella marina and Hydrogen Peroxide: Implications on the Cause of Fish Kills. J. Exp. Mar. Biol. Ecol. 2006, 336, 230–241. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Bagnyukova, T.V. Effects of Different Environmental Oxygen Levels on Free Radical Processes in Fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 144, 283–289. [Google Scholar] [CrossRef]
- Belhamra, R.; Tichati, L.; Trea, F.; Kada, S.; Mebarkia, A. Effect of Subacute Treatment with Bisphenol A on Oxidative Stress Biomarkers and Lipid Peroxidation in Gambusia affinis Mosquitofish. Toxicol. Environ. Health Sci. 2022, 15, 61–72. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Oxidants and Antioxidants. Exp. Physiol.: Transl. Integr. 1997, 82, 291–295. [Google Scholar] [CrossRef] [PubMed]
- White, C.C.; Viernes, H.; Krejsa, C.M.; Botta, D.; Kavanagh, T.J. Fluorescence-Based Microtiter Plate Assay for Glutamate–Cysteine Ligase Activity. Anal. Biochem. 2003, 318, 175–180. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, J.-C. Oxidative Stress, Neurotoxicity, and Non-Specific Immune Responses in Juvenile Red Sea Bream, Pagrus major, Exposed to Different Waterborne Selenium Concentrations. Chemosphere 2015, 135, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Aranda, J.J.; Seger, A.; Mardones, J.I.; Nichols, P.D.; Hallegraeff, G.M. Progress in Understanding Algal Bloom-Mediated Fish Kills: The Role of Superoxide Radicals, Phycotoxins and Fatty Acids. PLoS ONE 2015, 10, e0133549. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.E.D.; Dos Reis Martinez, C.B. The Pyrethroid λ-Cyhalothrin Induces Biochemical, Genotoxic, and Physiological Alterations in the Teleost Prochilodus lineatus. Chemosphere 2018, 210, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Nwani, C.D.; Lakra, W.S.; Nagpure, N.S.; Kumar, R.; Kushwaha, B.; Srivastava, S.K. Toxicity of the Herbicide Atrazine: Effects on Lipid Peroxidation and Activities of Antioxidant Enzymes in the Freshwater Fish Channa punctatus (Bloch). Int. J. Environ. Res. Public Health 2010, 7, 3298–3312. [Google Scholar] [CrossRef] [PubMed]
- Winston, G.W.; Di Giulio, R.T. Prooxidant and Antioxidant Mechanisms in Aquatic Organisms. Aquat. Toxicol. 1991, 19, 137–161. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA Damage: Mechanisms, Mutation, and Disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef]
- Mitchelmore, C.; Chipman, J. DNA Strand Breakage in Aquatic Organisms and the Potential Value of the Comet Assay in Environmental Monitoring. Mutat. Res. 1998, 399, 135–147. [Google Scholar] [CrossRef]
- Murphy, M.P.; Bayir, H.; Belousov, V.; Chang, C.J.; Davies, K.J.A.; Davies, M.J.; Dick, T.P.; Finkel, T.; Forman, H.J.; Janssen-Heininger, Y. Guidelines for Measuring Reactive Oxygen Species and Oxidative Damage in Cells and in Vivo. Nat. Metab. 2022, 4, 651–662. [Google Scholar] [CrossRef]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Walsh, C.J.; Nierenberg, K.; Clark, J.; Reich, A.; Hollenbeck, J.; Benson, J.; Cheng, Y.S.; et al. Review of Florida Red Tide and Human Health Effects. Harmful Algae 2011, 10, 224–233. [Google Scholar] [CrossRef]
- Satake, M.; Shoji, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Yasumoto, T. Gymnocin-A, a Cytotoxic Polyether from the Notorious Red Tide Dinoflagellate, Gymnodinium mikimotoi. Tetrahedron Lett. 2002, 43, 5829–5832. [Google Scholar] [CrossRef]
- Satake, M.; Tanaka, Y.; Ishikura, Y.; Oshima, Y.; Naoki, H.; Yasumoto, T. Gymnocin-B with the Largest Contiguous Polyether Rings from the Red Tide Dinoflagellate, Karenia (formerly Gymnodinium) mikimotoi. Tetrahedron Lett. 2005, 46, 3537–3540. [Google Scholar] [CrossRef]
- Liepelt, A.; Karbe, L.; Westendorf, J. Induction of DNA Strand Breaks in Rainbow Trout Oncorhynchus mykiss under Hypoxic and Hyperoxic Conditions. Aquat. Toxicol. 1995, 33, 177–181. [Google Scholar] [CrossRef]
- Dasgupta, S.; DiGiulio, R.T.; Drollette, B.D.; Plata, D.L.; Brownawell, B.J.; McElroy, A.E. Hypoxia Depresses CYP1A Induction and Enhances DNA Damage, but Has Minimal Effects on Antioxidant Responses in Sheepshead Minnow (Cyprinodon variegatus) Larvae Exposed to Dispersed Crude Oil. Aquat. Toxicol. 2016, 177, 250–260. [Google Scholar] [CrossRef]
- Shin, Y.K.; Nam, S.-E.; Kim, W.J.; Seo, D.Y.; Kim, Y.-J.; Rhee, J.-S. Red Tide Dinoflagellate Cochlodinium polykrikoides Induces Significant Oxidative Stress and DNA Damage in the Gill Tissue of the Red Seabream Pagrus major. Harmful Algae 2019, 86, 37–45. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, X.; Zhang, P. Combined Effects of Toxic Karenia mikimotoi and Hypoxia on the Juvenile Abalone Haliotis discus hannai. Front. Mar. Sci. 2023, 10, 1029512. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of Marine Planktonic Diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Kim, J.; Rhee, J.S. Biochemical and Physiological Responses of the Water Flea Moina macrocopa to Microplastics: A Multigenerational Study. Mol. Cell. Toxicol. 2021, 17, 523–532. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | 2014 | 2015 | 2016 | 2017~2022 |
|---|---|---|---|---|
| Initiation | 11 August | 4 August | 2 August | No occurrence |
| Termination | 22 August | 2 September | 26 August | - |
| Duration (d) | 12 | 30 | 25 | - |
| Affected area | Goseong | Yeosu, Goseong, Tongyeong, Busan | Yeosu | - |
| Max. density (cells/mL) | 2000 | 9000 | 2200 | - |
| Water temperature (°C) | 24.2~25.9 | 22.7~26.7 | 24.7~31.1 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.K.; Seo, D.Y.; Eom, H.-J.; Park, M.; Lee, M.; Choi, Y.-E.; Han, Y.-S.; Rhee, J.-S.; Kim, Y.-J. Oxidative Stress and DNA Damage in Pagrus major by the Dinoflagellate Karenia mikimotoi. Toxins 2023, 15, 620. https://doi.org/10.3390/toxins15100620
Shin YK, Seo DY, Eom H-J, Park M, Lee M, Choi Y-E, Han Y-S, Rhee J-S, Kim Y-J. Oxidative Stress and DNA Damage in Pagrus major by the Dinoflagellate Karenia mikimotoi. Toxins. 2023; 15(10):620. https://doi.org/10.3390/toxins15100620
Chicago/Turabian StyleShin, Yun Kyung, Do Yeon Seo, Hye-Jin Eom, Mira Park, Minji Lee, Young-Eun Choi, Young-Seok Han, Jae-Sung Rhee, and Youn-Jung Kim. 2023. "Oxidative Stress and DNA Damage in Pagrus major by the Dinoflagellate Karenia mikimotoi" Toxins 15, no. 10: 620. https://doi.org/10.3390/toxins15100620
APA StyleShin, Y. K., Seo, D. Y., Eom, H.-J., Park, M., Lee, M., Choi, Y.-E., Han, Y.-S., Rhee, J.-S., & Kim, Y.-J. (2023). Oxidative Stress and DNA Damage in Pagrus major by the Dinoflagellate Karenia mikimotoi. Toxins, 15(10), 620. https://doi.org/10.3390/toxins15100620