The Role of Heterocysts in Cyanotoxin Production during Nitrogen Limitation

Abstract
:1. Introduction
2. Results
2.1. Growth Curve Profile during Nitrogen (N) and Phosphorus (P) Starvation
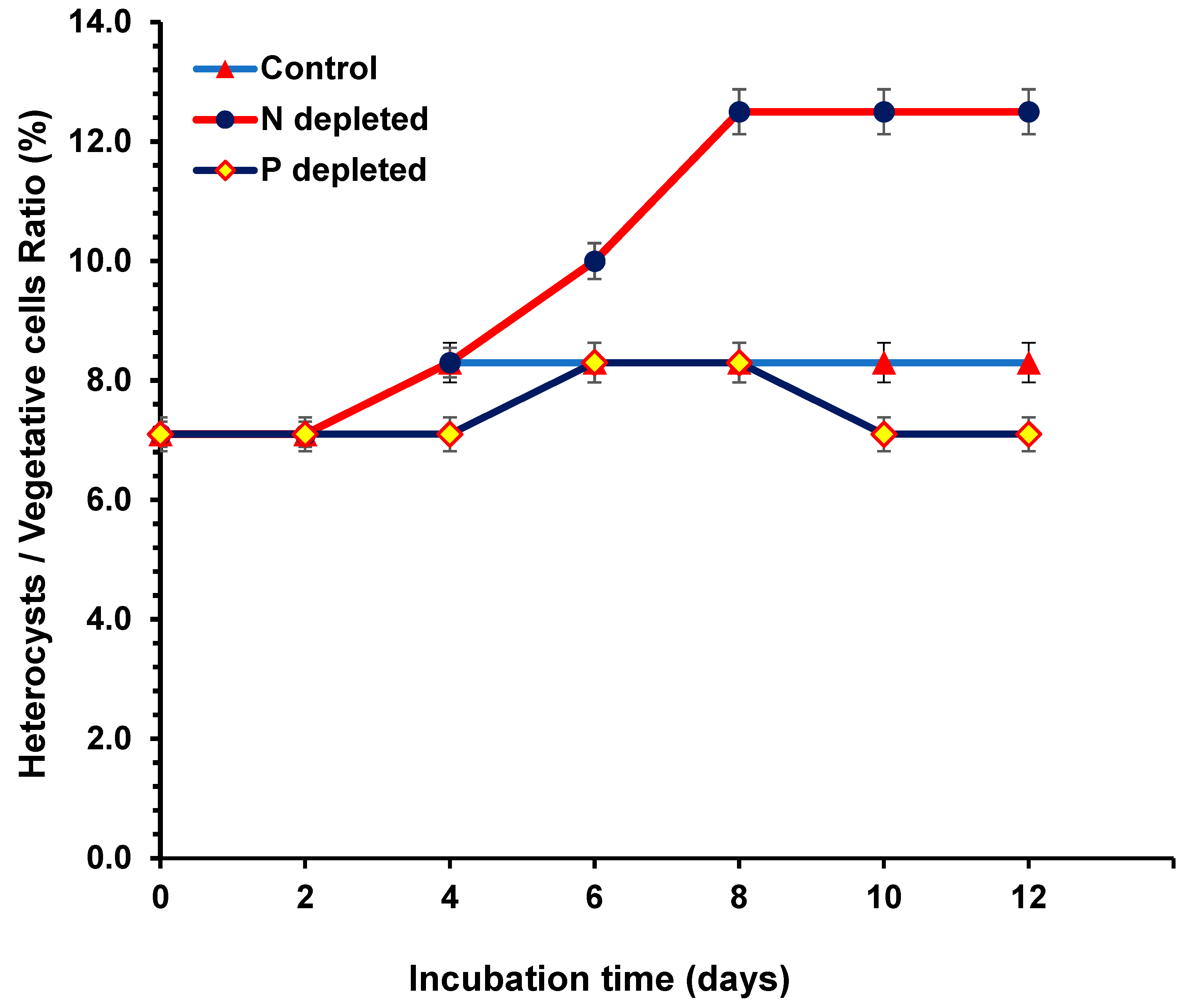
2.2. Heterocyst/Vegetative Ratio during N and P Starvation
2.3. Toxin Concentrations during Nitrogen and Phosphorus Starvation
2.4. Toxin Ratio Profile during Nitrogen and Phosphorus Starvation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cultures of Aphan sp. NH-5
5.1.1. Preparations of Nitrogen (N)- and Phosphorus (P)-Depleted Cultures
5.1.2. Determination of Dry Weight and Cell Count
5.2. Heterocyst Separation
5.3. Saxitoxin and Neosaxitoxin Determination by HPLC-FLD with Post-Column Derivatization
5.4. Quantitation and Calibration of Saxitoxin and Neosaxitoxin Determination by HPLC-FLD with Post-Column Derivatization
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watson, S.B.; Molet, L. Harmful Algal Blooms. In Encyclopedia of Aquatic Ecotoxicology; Férard, J.F., Blaise, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 13, pp. 13–22. Available online: http://www.springerreference.com/index/chapterdbid/379970 (accessed on 21 August 2022).
- USEPA (United States Environmental Protection Agency). Cyanobacterial Harmful Algal Blooms (CyanoHABs) in Water Bodies. 2021. Available online: https://www.epa.gov/nutrientpollution/harmful-algal-blooms (accessed on 21 August 2022).
- Schindler, D. Evolution of phosphorus limitation in lakes. Science 1977, 195, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Nakahara, H. Life Cycle of Cyanobacterium Aphanizomenon flos-aquae. Taiwania 2009, 54, 113–117. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, C.C. The Making of a Heterocyst in Cyanobacteria. Annu. Rev. Microbiol. 2022, 8, 597–618. [Google Scholar] [CrossRef]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef]
- Fuhs, G.W. Cytology of blue-green algae: Light microscopic aspects. In Algae, Man, and the Environment; Jackson, D.F., Ed.; Syracuse University Press: New York, NY, USA, 1967; pp. 213–233. [Google Scholar]
- Laamanen, M.; Kuosa, H. Annual variability of biomass and heterocysts of the N2-fixing cyanobacterium Aphanizomenon flos-aquae in the Baltic Sea with reference to Anabaena spp. and Nodularia spumigena. Boreal. Environ. Res. 2005, 10, 19–30. [Google Scholar]
- Ecker, M.M.; Foxall, T.L.; Sasner, J.J., Jr. Morphology of toxic versus non-toxic strains of Aphanizomenon flos-aquae. In The Water Environment; Carmichael, W.W., Ed.; Plenum Press: New York, NY, USA, 1981; pp. 113–126. [Google Scholar] [CrossRef]
- Sawyer, P.J.; Gentile, J.H.; Sasner, J.J., Jr. Demonstration of a toxin from Aphanizomenon flos-aquae (L.) Ralfs. Can. J. Microbiol. 1986, 14, 199–1204. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Mahmood, N.A. Toxins from freshwater cyanobacteria. In Seafood Toxins; Ragelis, E.P., Ed.; American Chemical Society: Washington, DC, USA, 1984; pp. 377–389. [Google Scholar]
- Ikawa, M.; Wegener, K.; Foxall, T.J.L.; Sasner, J.J., Jr. Comparison of the toxins of the blue-green alga Aphanizomenon flos-aquae with the Gonyaulax tamarensis var excavata toxins. Toxicon 1982, 20, 747–752. [Google Scholar] [CrossRef]
- Mahmood, N.A.; Carmichael, W.W. Paralytic shellfish poisons produced by the freshwater cyanobacterium Aphanizomenon flos-aquae NH-5. Toxicon 1986, 24, 175–186. [Google Scholar] [CrossRef]
- Gomaa, M.N. Toxins and Toxicity of a Blue-Green Alga Aphanizomenon flos-aquae. Ph.D. Thesis, Marine Estuarine Environment Science, University of Maryland, College Park, MD, USA, 1990. [Google Scholar]
- Hall, S. Toxins and Toxicity of Protogonyaulax from the Northeast Pacific. Ph.D. Thesis, University of Alaska, Fairbanks, AK, USA, 1982. [Google Scholar]
- Watanabe, M.F.; Oishi, S. Effect of environmental factors on toxicity of a cyanobacterium Microcystis aeruginosa under culture conditions. Appl. Environ. Microbiol. 1985, 49, 1342–1344. [Google Scholar] [CrossRef]
- Codd, G.A.; Poon, G.K. Cyanobacterial Toxins. In Biochemistry of the Algae and Cyanobacteria; Rogers, L.J., Gallon, J.R., Eds.; Clarendon Press: Oxford, UK, 1988; pp. 283–296. [Google Scholar] [CrossRef]
- Sivonen, K. Effect of light, temperature, nitrate, orthophosphate, and bacteria on growth of and hepatotoxin production by Oscillaioria agardhii strains. Appl. Environ. Microbiol. 1990, 56, 2658–2666. [Google Scholar] [CrossRef] [PubMed]
- Vezie, C.; Rapala, J.; Vaitomaa, J.; Seitsonen, J.; Sivonen, K. Effect of nitrogen and phosphorus on growth of toxic and non-toxic Microcystis strains and on intracellular microcystin concentrations. Microb. Ecol. 2002, 43, 443–454. [Google Scholar] [CrossRef]
- Downing, T.G.; Meyer, C.; Gehringer, M.M.; van de Venter, M. Microcystin content of Microcystis aeruginosa is modulated by nitrogen uptake rate relative to specific growth rate or carbon fixation rate. Environ. Toxicol. 2005, 20, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Sivonen, K.; Luukkainen, R.; Niemela, S.I. Anatoxin-a concentration in Anabaena and Aphanizomenon under different environmental-conditions and comparison of growth by toxic and nontoxic Anabaena-strains—A laboratory study. J. Appl. Phycol. 1993, 5, 581–591. [Google Scholar] [CrossRef]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Neilan, B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (nostocales, cyanophyceae) from Northern Australia. Appl. Environ. Microbiol. 2001, 67, 1839–1845. [Google Scholar] [CrossRef]
- Repka, S.; Mehtonen, J.; Vaitomaa, J.; Saari, L.; Sivonen, K. Effects of nutrients on growth and nodularin production of Nodularia strain GR8b. Microb. Ecol. 2001, 42, 606–613. [Google Scholar] [CrossRef]
- de Figueiredo, D.R.; Reboleira, A.S.S.P.; Antunes, S.C.; Abrantes, N.; Azeiteiro, U.; Gonçalves, F.; Pereira, M.J. The effect of environmental parameters and cyanobacterial blooms on phytoplankton dynamics of a Portuguese temperate Lake. Hydrobiologia 2006, 568, 145–157. [Google Scholar] [CrossRef]
- Teubner, K.; Feyerabend, R.; Henning, M.; Nicklisch, A.; Woitke, P.; Kohl, J.-G. Alternative blooming of Aphanizomenon flos-aquae or Planktothrix agardhii induced by the timing of the critical nitrogen: Phosphorus ratio in hypertrophic riverine lakes. Arch. Hydrobiol. Spec. Issues Adv. Limnol. 1999, 54, 325–344. [Google Scholar]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Oliver, R.; Ganf, G. Freshwater Blooms. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B., Potts, M., Eds.; Kluwer Academic Publishers: Kluwer, The Netherlands, 2000; pp. 149–194. [Google Scholar] [CrossRef]
- Camargo, S.; Valladares, A.; Flores, E.; Herrero, A. Transcription activation by NtcA in the absence of consensus NtcA-binding sites in an anabaena heterocyst differentiation gene promoter. J. Bacteriol. 2012, 194, 2939–2948. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, J.D.; Westrick, J.A.; Reitz, L.A.; Bridgeman, T.B. Microcystin congeners in Lake Erie follow the seasonal pattern of nitrogen availability. Harmful Algae 2023, 127, 102466. [Google Scholar] [CrossRef] [PubMed]
- Yunes, J.S.; De La Rocha, S.; Giroldo, D.; Silveira, S.B.; Comin, R.; Bicho, M.; Melcher, S.S.; Sant’Anna, C.L.; Vieira, A.A. Release of carbohydrates and proteins by a subtropical strain of Raphidiopsis brookii (cyanobacteria) able to produce saxitoxin at three nitrate concentrations. J. Phycol. 2009, 45, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, G.; Wallström, K.; Brattberg, G. Short-term variations in nitrogen fixation in a coastal area of the northern Baltic. Arch. Hydrobiol. 1980, 89, 88–100. [Google Scholar]
- Riddolls, A. Aspects of nitrogen fixation in Lough Neagh. I. Acetylene reduction and the frequency of Aphanizomenon flos-aquae heterocytes. Freshwat. Biol. 1985, 15, 289–298. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef]
- Dolman, A.M.; Rücker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and Cyanotoxins: The Influence of Nitrogen versus Phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef]
- Carmichael, W.W. Freshwater Blue-Green Algae (Cyanobacteria) Toxins—A Review. In The Water Environment. Environmental Science Research; Carmichael, W.W., Ed.; Springer: Boston, MA, USA, 1981. [Google Scholar] [CrossRef]
- Li, R.; Carmichael, W.W.; Liu, Y.; Watanabe, M.M. Taxonomic re-evaluation of Aphanizomenon flos-aquae NH-5 based upon morphology and 16S rRNA gene sequences. Hydrobiologia 2000, 438, 99–105. [Google Scholar] [CrossRef]
- Dreher, T.W.; Davis, E.W., II; Muller, R.S.; Otten, T.G. Comparative genomics of the ADA clade within the Nostocales. Harmful Algae 2021, 104, 102037. [Google Scholar] [CrossRef]
- Allen, M.M. Methods for cyanophyceae. In Handbook of Phycological Methods, Culture Methods, and Growth Measurements; Stein, L.R., Ed.; Cambridge University Press: London, UK, 1973; pp. 127–238. ISBN 9780521297479. [Google Scholar]
- Rippka, R.; Deruelles, J.; Waterbury, J.; Herdman, M.; Stanier, R. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Martinez, M.R.; Chakroff, R.P.; Pantastico, J. Direct phytoplankton counting techniques using the haemocytometer. Philipp. Agric. 1975, 59, 43–50. Available online: https://eurekamag.com/research/018/746/018746327.php (accessed on 14 June 2023).
- Smith, R.L.; Van Baalen, C.; Tabita, F.R. Isolation of metabolically active heterocysts from cyanobacteria. Methods Enzymol. 1988, 167, 490–495. [Google Scholar] [CrossRef]
- Oshima, Y.; Itakura, H.; Lee, K.C.; Yosumoto, T.; Blackburn, S.; Hallegraeff, G. Toxin production by the dinoflagellate Gymnodium catenatun. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier Science Publishers, B.V.: Amsterdam, The Netherlands, 1993; pp. 907–912. [Google Scholar]
- SAS Institute: SAS/STAT User’s Guide Release 6.03 Edition; SAS Institute: Cary, NC, USA, 1988.
- Winer, B.J. Statistical Principles in Experimental Design, 2nd ed.; Mi Graw-Hill Kogakusha, LTD: Tokyo, Japan, 1971. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Component | Stock Solution | Quantity | Molar Concentration in Final Medium |
|---|---|---|---|---|
| 1. | MgNa2 EDTA 3H2O | -------- | 1.000 g | 2.26 µM |
| 2. | H3BO3 | -------- | 2.860 g | 46.3 µM |
| 3. | MnCl2 4H2O | -------- | 1.810 g | 9.15 µM |
| 4. | ZnSO4 7H2O | -------- | 0.220 g | 0.765 µM |
| 5. | CuSO4 5H2O | 79.0 g L−1 H2O | 1 mL | 0.316 µM |
| No. | Constituent | Volume | Concentration of the Stock Solution | Final Molar Concentration |
|---|---|---|---|---|
| 1 | NaNO3 | 10 mL/L | 30 g/200 mL dH2O | 17.6 mM |
| 2 | K2HPO4 | 10 mL/L | 0.8 g/200 mL dH2O | 0.23 mM |
| 3 | MgSO4 7H2O | 10 mL/L | 1.5 g/200 mL dH2O | 0.3 mM |
| 4 | CaCl2 2H2O | 10 mL/L | 0.72 g/200 mL dH2O | 0.24 mM |
| 5 | Citric Acid H2O | 10 mL/L | 0.12 g/200 mL dH2O | 0.031 mM |
| 6 | Ferric Ammonium Citrate | 10 mL/L | 0.12 g/200 mL dH2O | 0.021 mM |
| 7 | Na2EDTA 2H2O | 10 mL/L | 0.02 g/200 mL dH2O | 0.0027 mM |
| 8 | Na2CO3 | 10 mL/L | 0.4 g/200 mL dH2O | 0.19 mM |
| 9 | BG-11 Trace Metals Solution | 1 mL/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomaa, M.N.; Carmichael, W.W. The Role of Heterocysts in Cyanotoxin Production during Nitrogen Limitation. Toxins 2023, 15, 611. https://doi.org/10.3390/toxins15100611
Gomaa MN, Carmichael WW. The Role of Heterocysts in Cyanotoxin Production during Nitrogen Limitation. Toxins. 2023; 15(10):611. https://doi.org/10.3390/toxins15100611
Chicago/Turabian StyleGomaa, Mohamed N., and Wayne W. Carmichael. 2023. "The Role of Heterocysts in Cyanotoxin Production during Nitrogen Limitation" Toxins 15, no. 10: 611. https://doi.org/10.3390/toxins15100611
APA StyleGomaa, M. N., & Carmichael, W. W. (2023). The Role of Heterocysts in Cyanotoxin Production during Nitrogen Limitation. Toxins, 15(10), 611. https://doi.org/10.3390/toxins15100611