Kinetics, Thermodynamics and Mechanism of Enzymatic Degradation of Zearalenone in Degummed Corn Oil



Abstract
1. Introduction
2. Results and Discussion
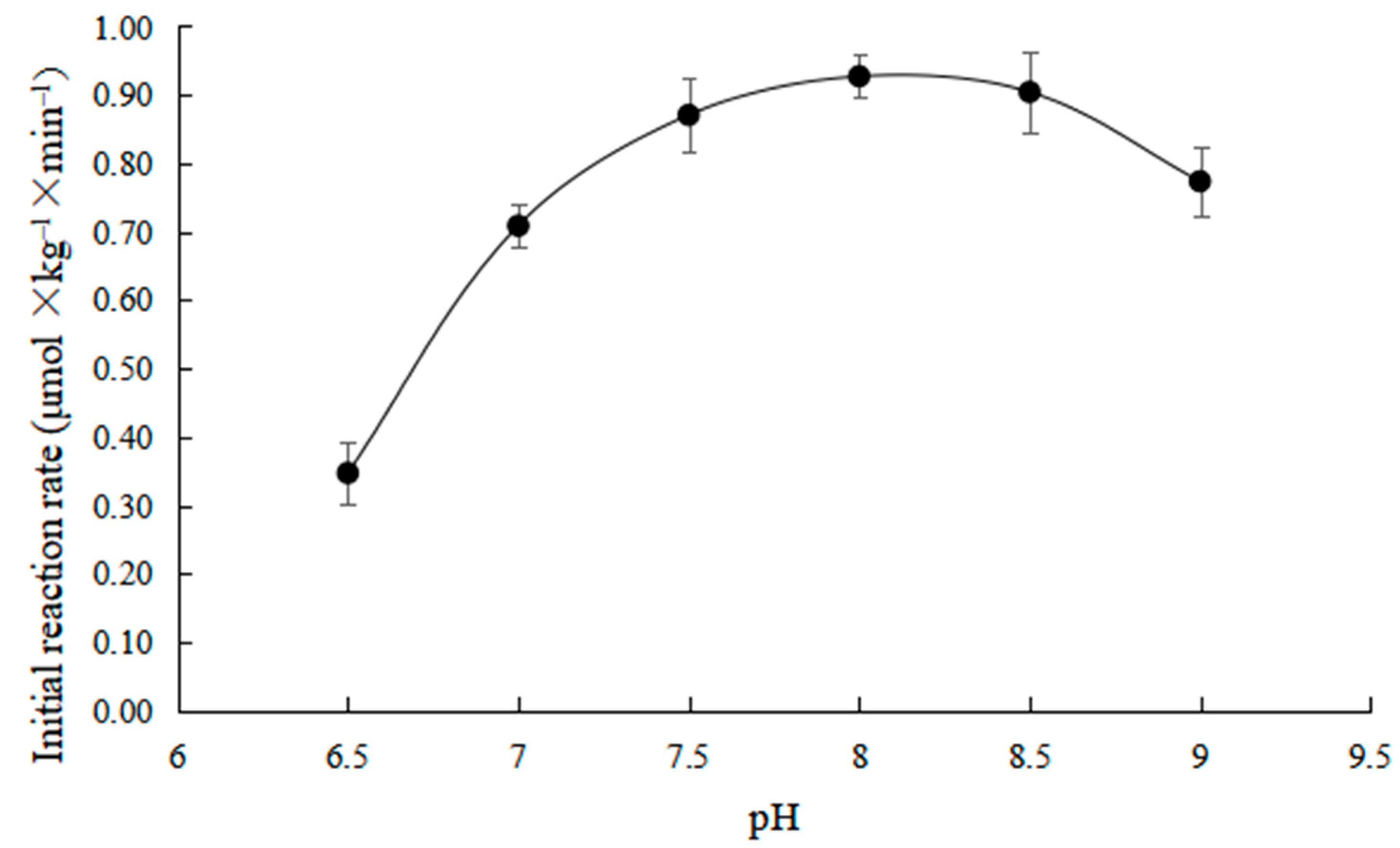
2.1. Effects of pH on Initial Reaction Rate
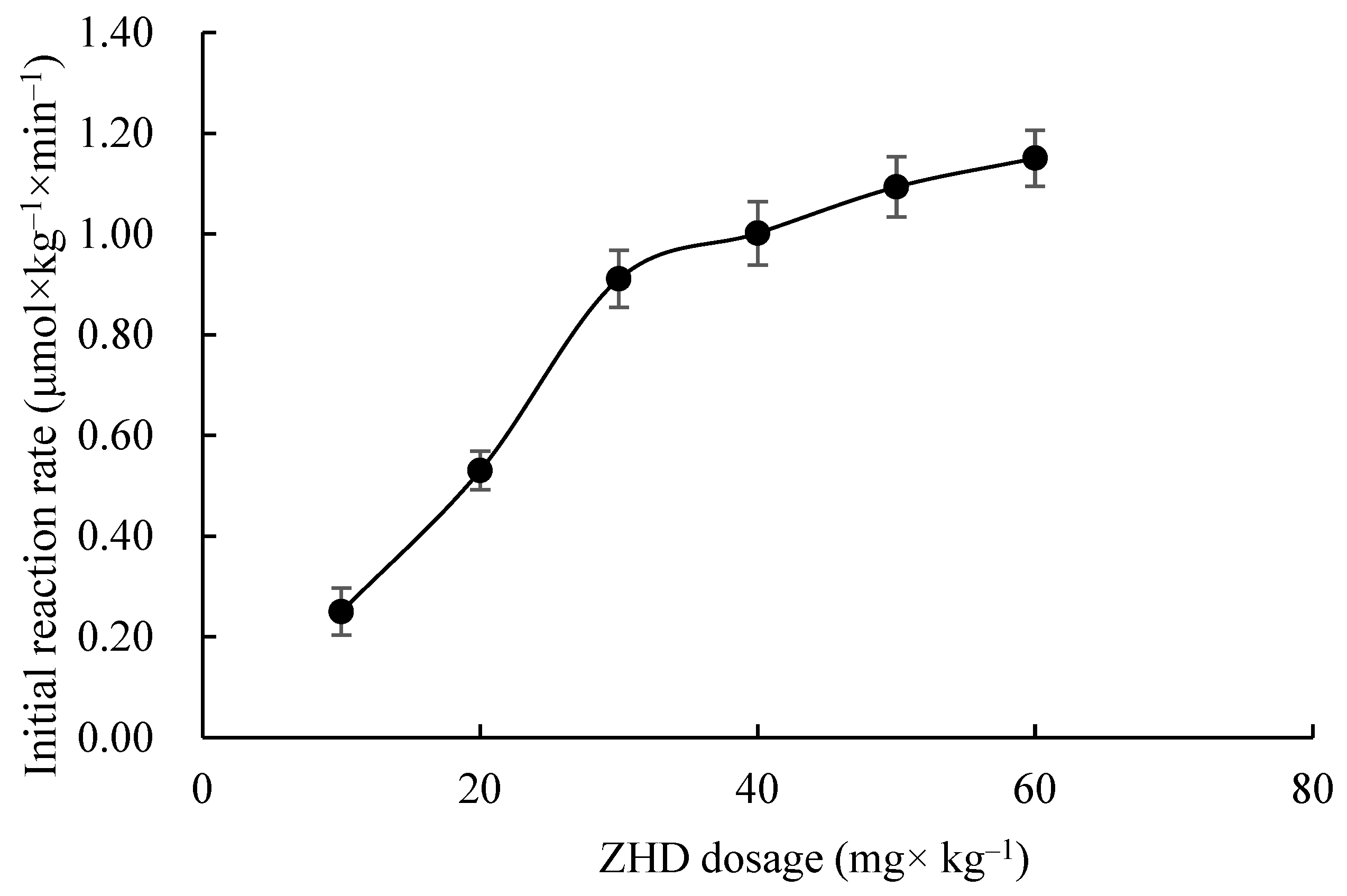
2.2. Effects of ZHD Dosage on Initial Reaction Rate
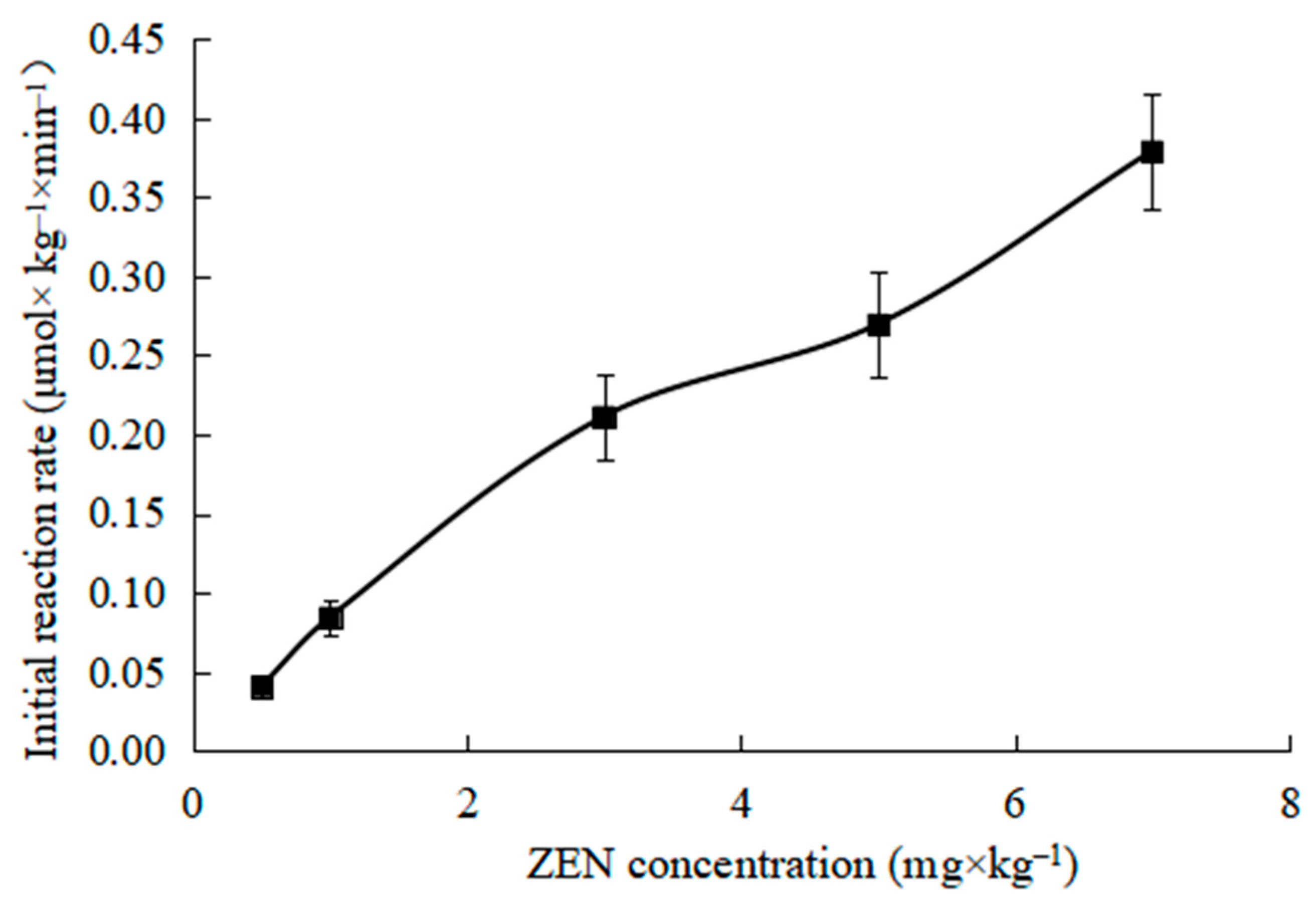
2.3. Effects of ZEN Concentration on Initial Reaction Rate
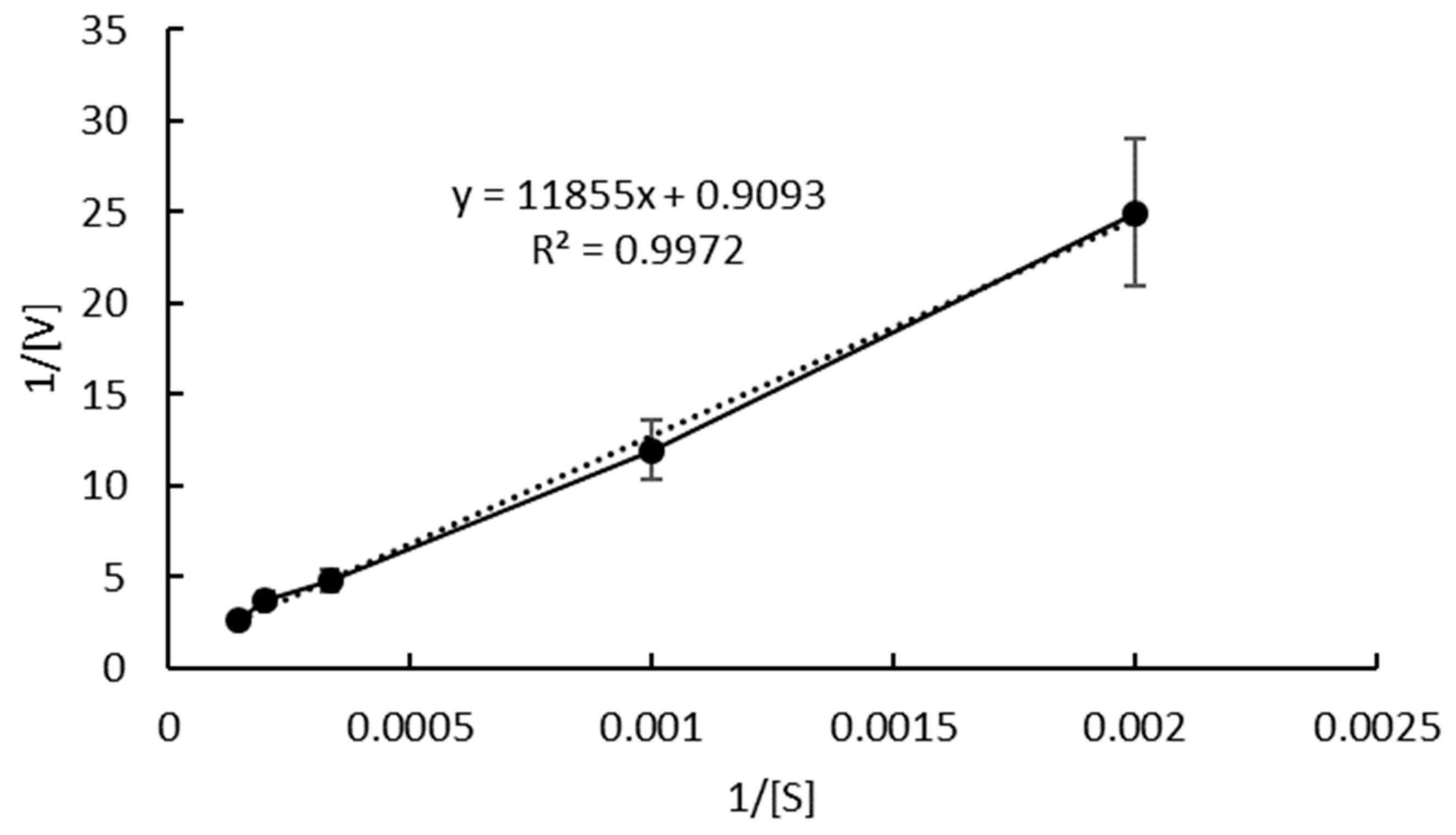
2.4. Enzyme Reaction Kinetics
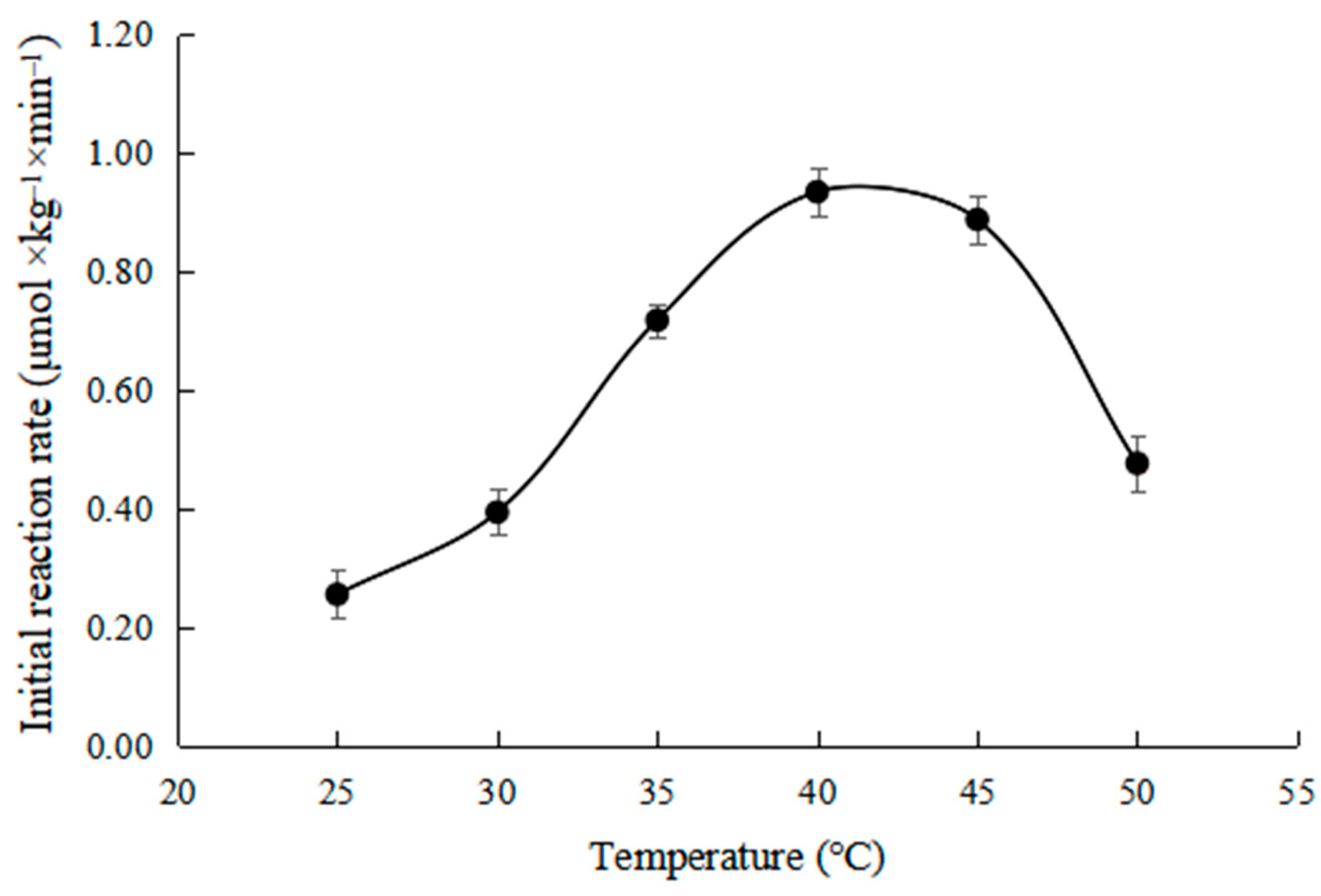
2.5. Effects of Temperature on Initial Reaction Rate
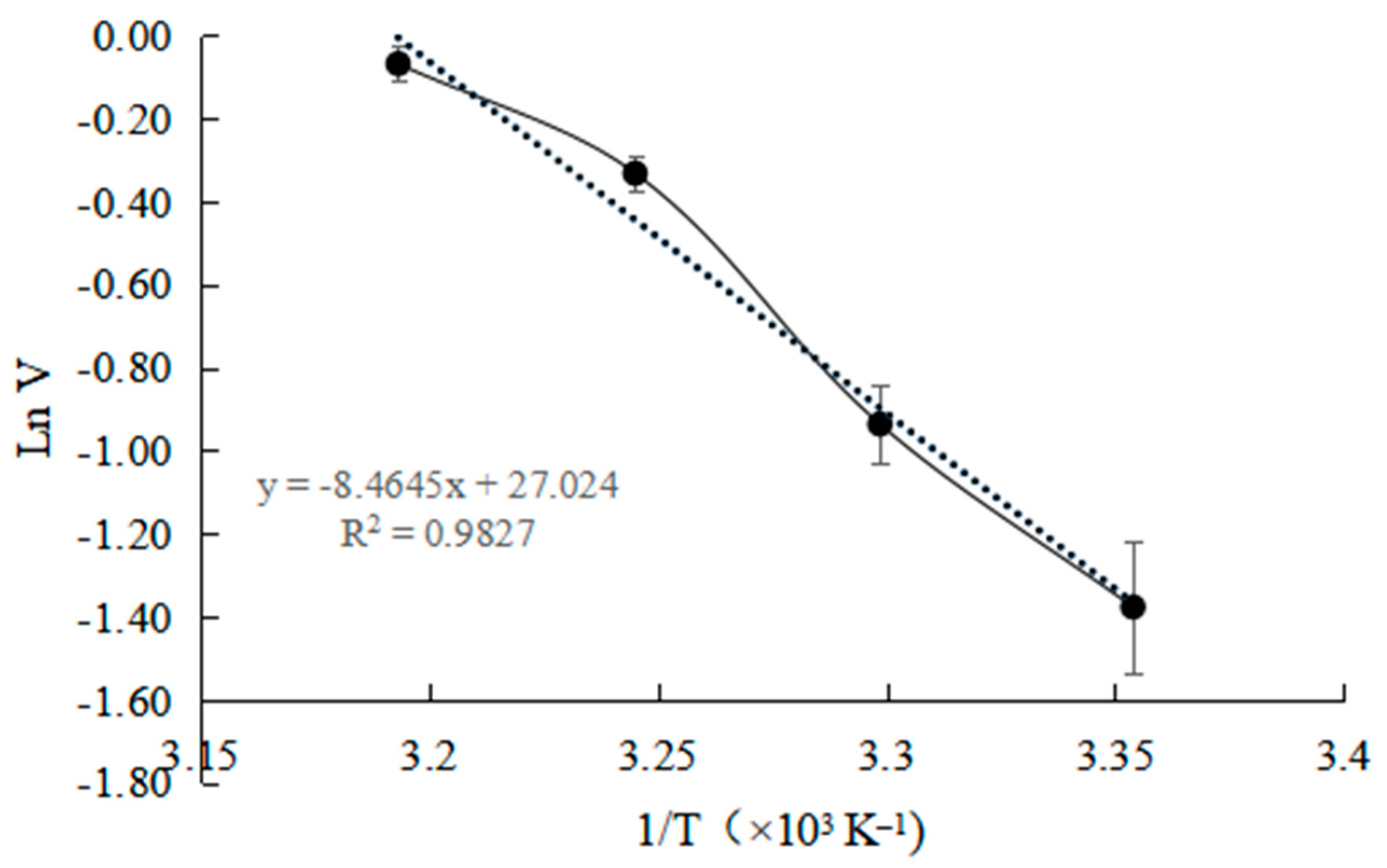
2.6. Determination of Thermodynamic Parameters
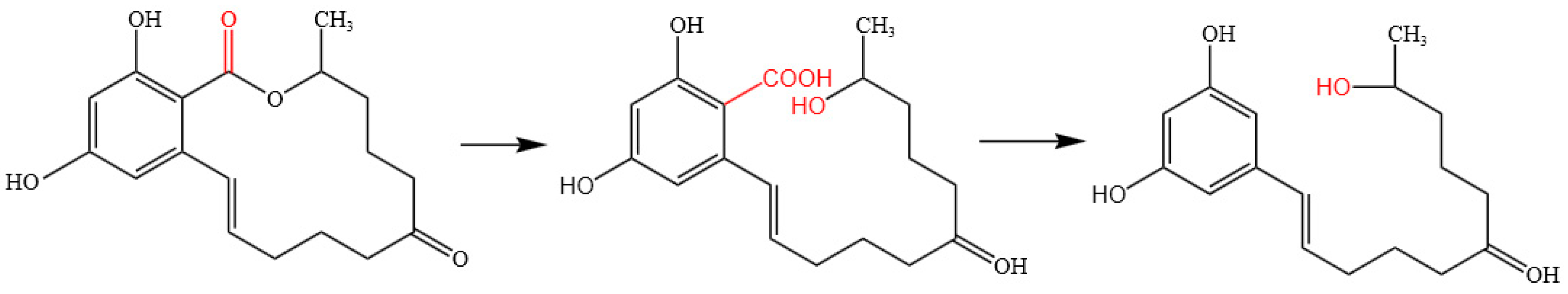
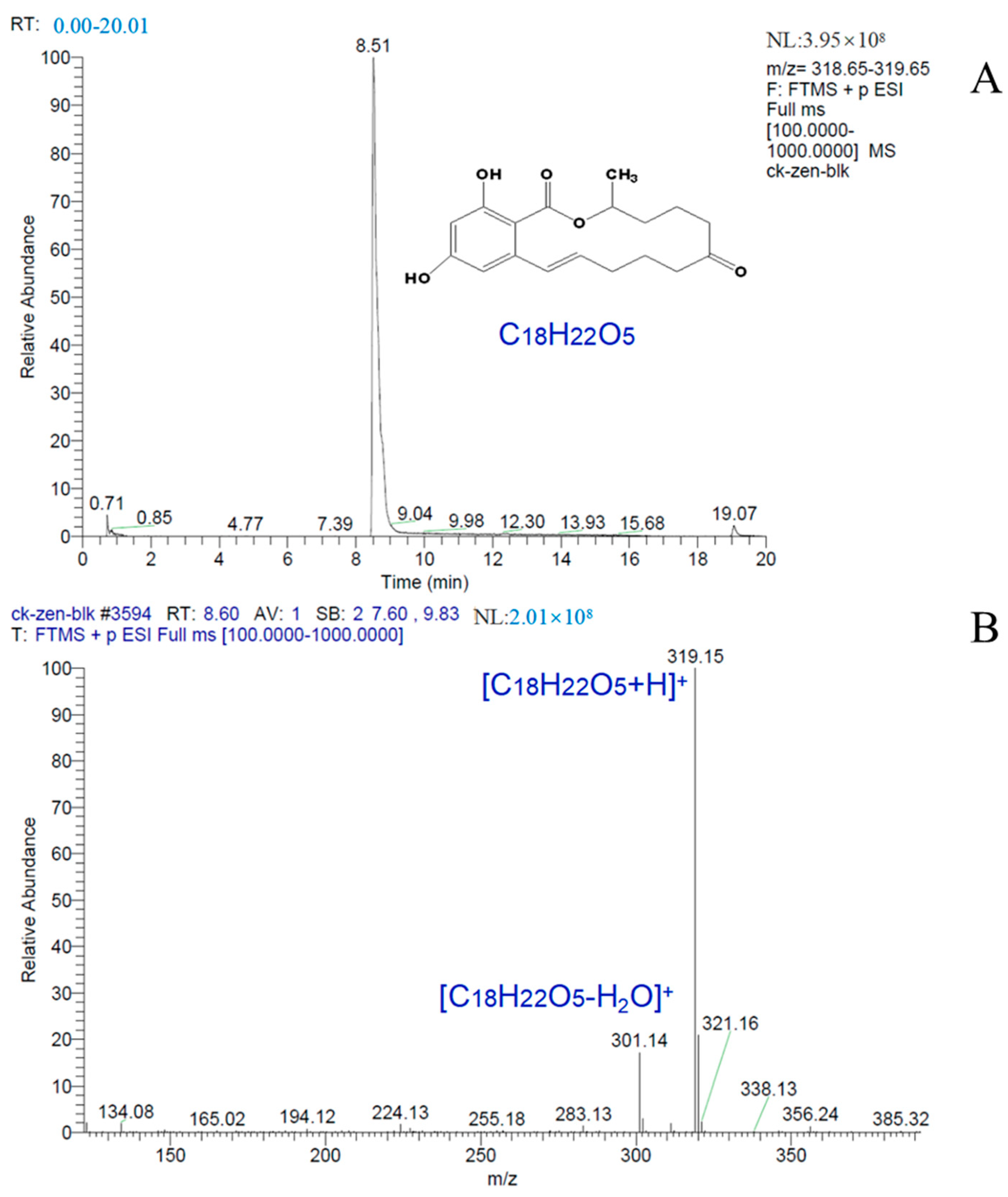
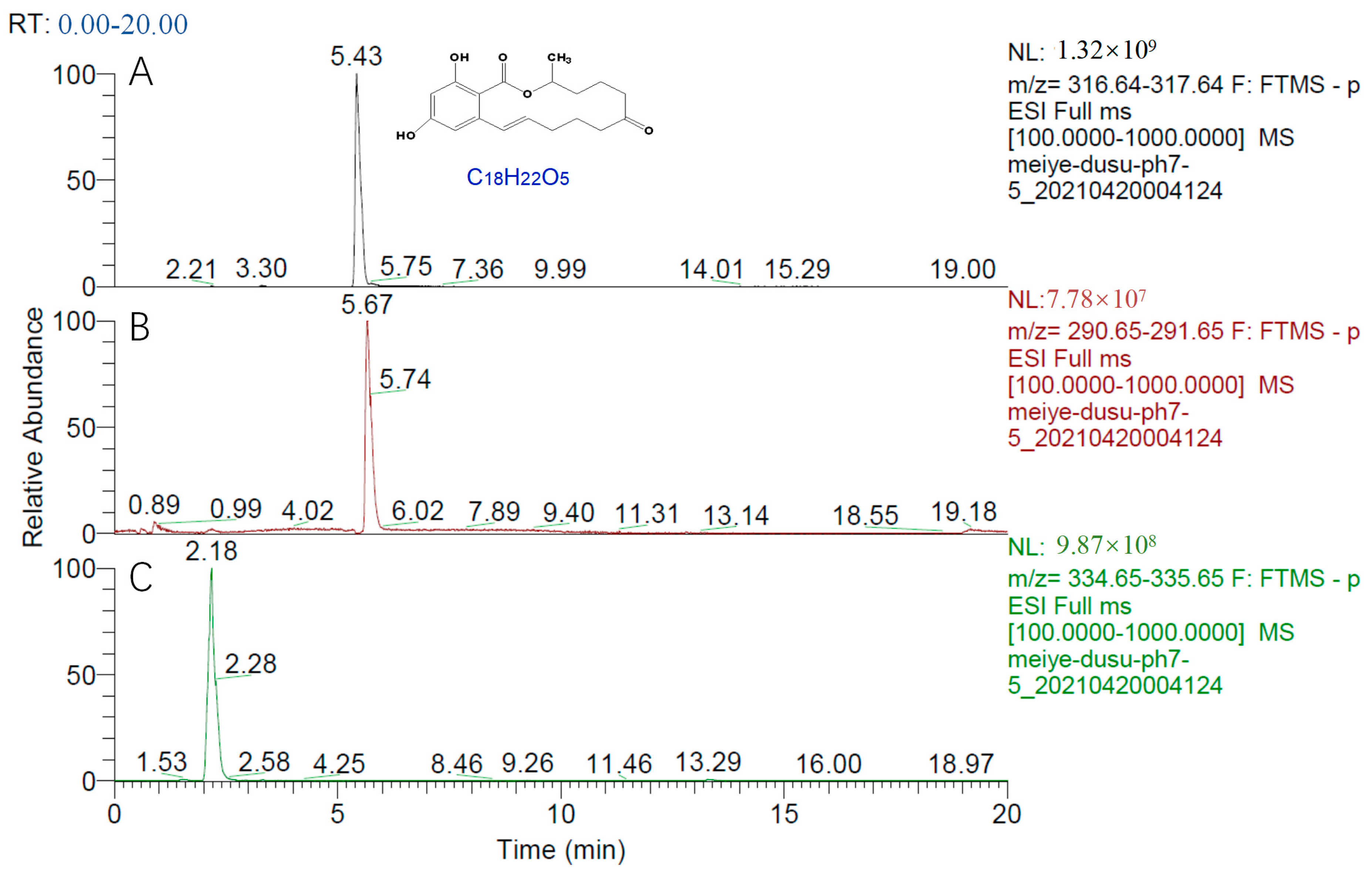
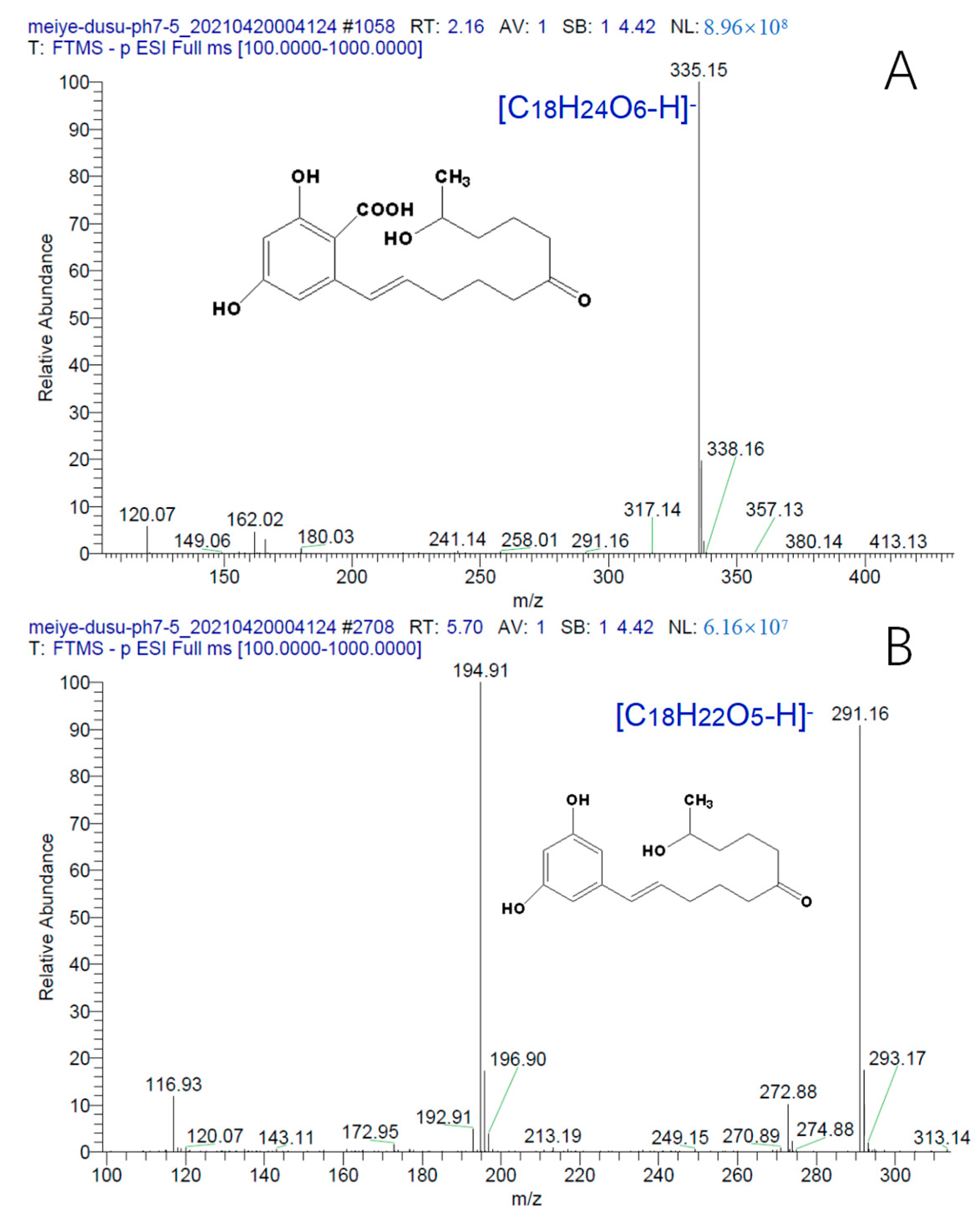
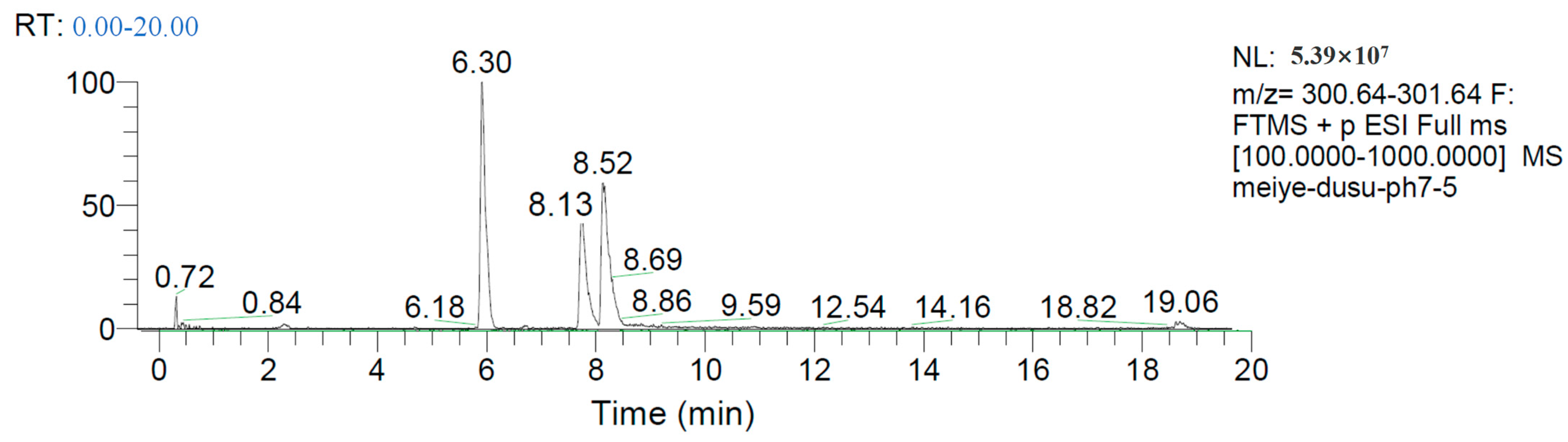
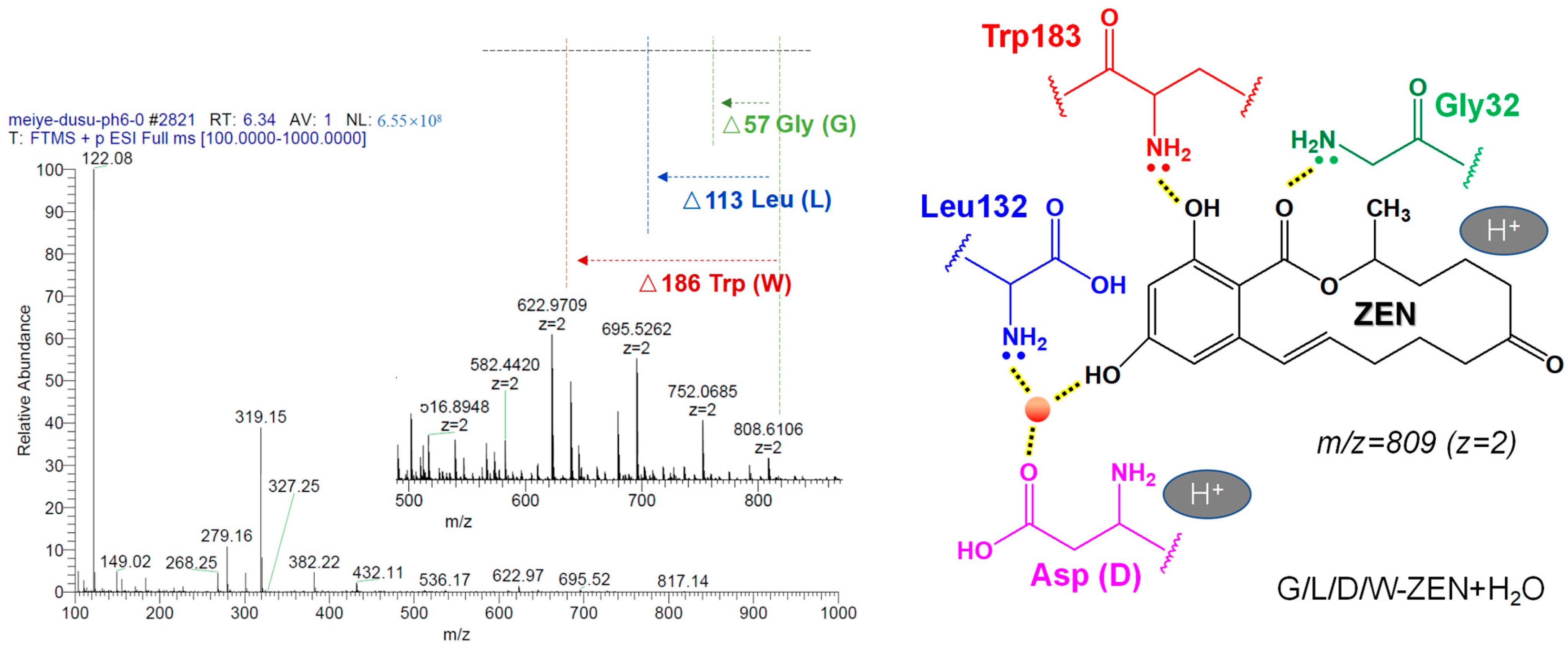
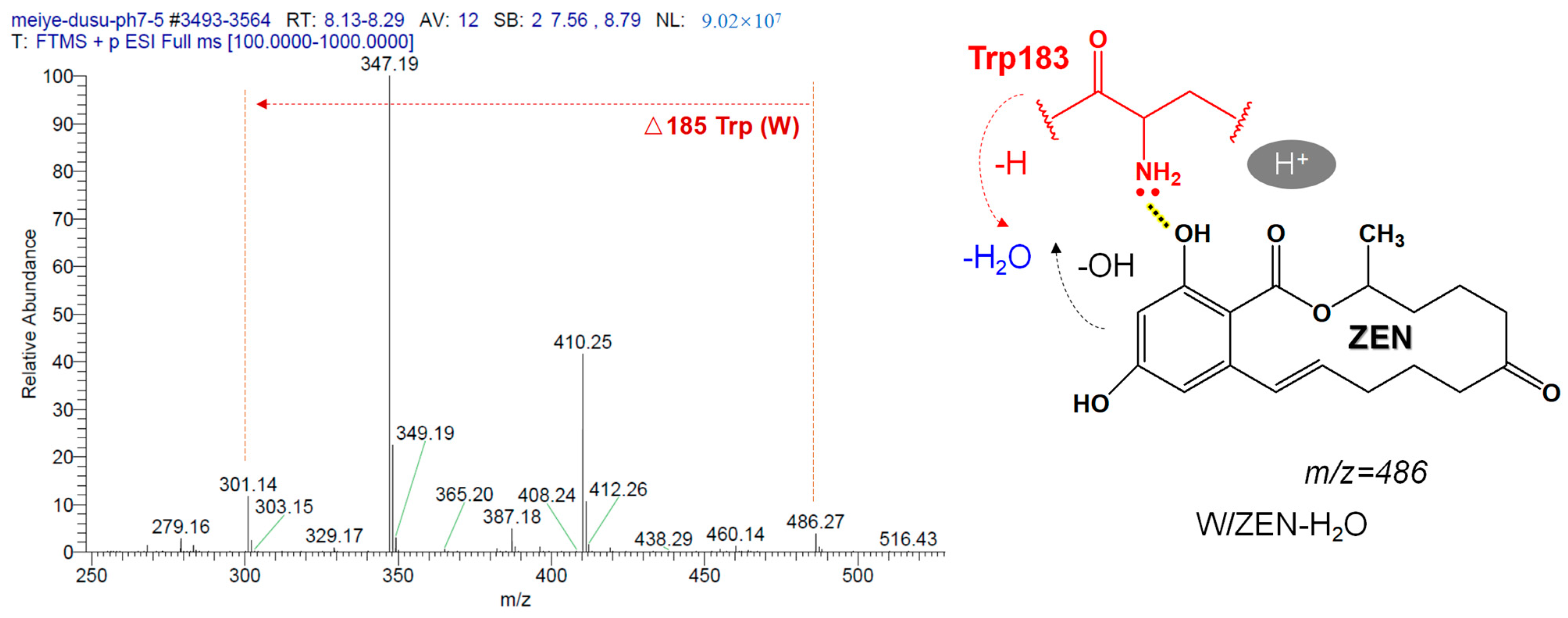
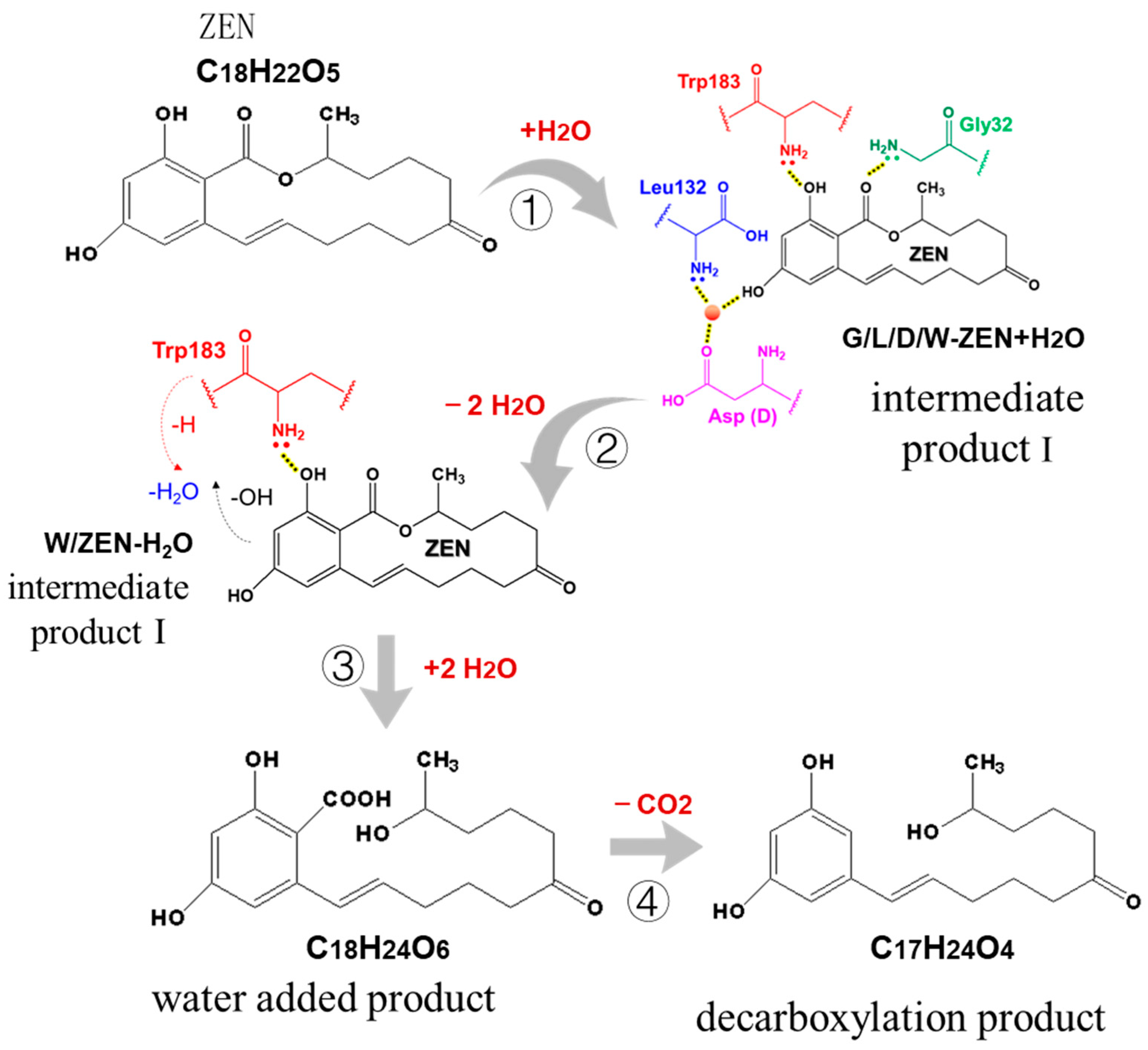
2.7. Mechanism of ZEN Hydrolase
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Detection of ZEN Content
4.2.2. Determination of the Initial Reaction Rate of ZEN Hydrolase
4.2.3. Determination of Kinetic Constants and Thermodynamic Parameters
4.2.4. Determination of ZEN Degradation Products by Hybrid Quadrupole Orbitrap Mass Spectrometer
4.3. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stob, M.; Baldwin, R.; Tuite, J.; Andrews, F.; Gillette, K. Isolation of an anabolic, uterotrophic compound from corn infected with Gibberella zeae. Nature 1962, 196, 1318. [Google Scholar] [CrossRef] [PubMed]
- Fruhauf, S.; Novak, B.; Nagl, V.; Hackl, M.; Hartinger, D.; Rainer, V.; Labudová, S.; Adam, G.; Aleschko, M.; Moll, W.-D. Biotransformation of the mycotoxin zearalenone to its metabolites hydrolyzed zearalenone (HZEN) and decarboxylated hydrolyzed zearalenone (DHZEN) diminishes its estrogenicity in vitro and in vivo. Toxins 2019, 11, 481. [Google Scholar] [CrossRef] [PubMed]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Shiinoki, Y. Microstructure and rheological behavior of whipping cream. J. Texture Stud. 1986, 17, 189–204. [Google Scholar] [CrossRef]
- Szabó-Fodor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Bóta, B.; Kachlek, M.; Mézes, M.; Balogh, K.; Kövér, G.; Nagy, I. Interaction between the three frequently co-occurring Fusarium mycotoxins in rats. J. Anim. Physiol. Anim. Nutr. 2019, 103, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, M.G.; Di Giacomo, A.; Lara, A.; Martinez, M.P.; Cavaglieri, L. Aflatoxin-degrading Bacillus sp. strains degrade zearalenone and produce proteases, amylases and cellulases of agro-industrial interest. Toxicon 2020, 180, 43–48. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulè, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef]
- Kalagatur, N.K.; Kamasani, J.R.; Mudili, V. Assessment of detoxification efficacy of irradiation on zearalenone mycotoxin in various fruit juices by response surface methodology and elucidation of its in-vitro toxicity. Front. Microbiol. 2018, 9, 2937. [Google Scholar] [CrossRef]
- Bai, X.; Sun, C.; Xu, J.; Liu, D.; Han, Y.; Wu, S.; Luo, X. Detoxification of zearalenone from corn oil by adsorption of functionalized GO systems. Appl. Surf. Sci. 2018, 430, 198–207. [Google Scholar] [CrossRef]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A. A review of the mycotoxin adsorbing agents, with an emphasis on their multi-binding capacity, for animal feed decontamination. Food Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.; Ji, J.; Wu, H.; Pi, F.; Zhang, Y.; Sun, X. Chemical and toxicological alterations of zearalenone under ozone treatment. Food Addit. Contam. Part A 2019, 36, 163–174. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, L.; Chen, J.; Wang, W.; Zhang, R.; Li, Y.; Zhang, Q.; Wang, W. Degradation mechanism for Zearalenone ring-cleavage by Zearalenone hydrolase RmZHD: A QM/MM study. Sci. Total Environ. 2020, 709, 135897. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Ando, N.; Kimura, M.; Kakeya, H.; Osada, H.; Yamaguchi, I. A novel lactonohydrolase responsible for the detoxification of zearalenone: Enzyme purification and gene cloning. Biochem. J. 2002, 365, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Tu, T.; Luo, H.; Huang, H.; Su, X.; Wang, Y.; Wang, Y.; Zhang, J.; Bai, Y.; Yao, B. Biochemical characterization and mutational analysis of a lactone hydrolase from Phialophora americana. J. Agric. Food Chem. 2019, 68, 2570–2577. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of zearalenone by genetically modified organisms expressing the detoxification gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [CrossRef]
- Yang, W.-C.; Hsu, T.-C.; Cheng, K.-C.; Liu, J.-R. Expression of the Clonostachys rosea lactonohydrolase gene by Lactobacillus reuteri to increase its zearalenone-removing ability. Microb. Cell Factories 2017, 16, 1–11. [Google Scholar]
- Bi, K.; Zhang, W.; Xiao, Z.; Zhang, D. Characterization, expression and application of a zearalenone degrading enzyme from Neurospora crassa. AMB Express 2018, 8, 194. [Google Scholar] [CrossRef]
- Wang, M.; Yin, L.; Hu, H.; Selvaraj, J.N.; Zhou, Y.; Zhang, G. Expression, functional analysis and mutation of a novel neutral zearalenone-degrading enzyme. Int. J. Biol. Macromol. 2018, 118, 1284–1292. [Google Scholar] [CrossRef]
- Hui, R.; Hu, X.; Liu, W.; Zheng, Y.; Chen, Y.; Guo, R.-T.; Jin, J.; Chen, C.-C. Characterization and crystal structure of a novel zearalenone hydrolase from Cladophialophora bantiana. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 515–519. [Google Scholar] [CrossRef]
- Qi, Q.; Yang, W.-J.; Zhou, H.-J.; Ming, D.-M.; Sun, K.-L.; Xu, T.-Y.; Hu, X.-J.; Lv, H. The structure of a complex of the lactonohydrolase zearalenone hydrolase with the hydrolysis product of zearalenone at 1.60 Å resolution. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Liu, H.; Sun, J.; Wang, J.; Zhao, C.; Zhang, W.; Zhang, J.; Sun, C. Zearalenone removal from corn oil by an enzymatic strategy. Toxins 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Baici, A. Kinetics of Enzyme-Modifier Interactions; Springer: Vienna, Austria, 2015. [Google Scholar]
- Ray, J.C.J.; Wickersheim, M.L.; Jalihal, A.P.; Adeshina, Y.O.; Cooper, T.F.; Balázsi, G. Cellular growth arrest and persistence from enzyme saturation. PLoS Comput. Biol. 2016, 12, 1553–1734X. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Wang, M.; Li, X.; Wang, H.; Zhao, G.; Wu, P.; Lu, Z.; Zhang, G. The replacement of main cap domain to improve the activity of a ZEN lactone hydrolase with broad substrate spectrum. Biochem. Eng. J. 2022, 182, 108418. [Google Scholar] [CrossRef]
- Glick, N.; Landman, A.; Roufogalis, B. Correcting Lineweaver—Burt calculations of V and Km. Trends Biochem. Sci. 1979, 4, N82–N83. [Google Scholar] [CrossRef]
- Johnson, K.A. A century of enzyme kinetic analysis, 1913 to 2013. FEBS Lett. 2013, 587, 2753–2766. [Google Scholar] [CrossRef]
- Feller, G. Molecular adaptations to cold in psychrophilic enzymes. Cell. Mol. Life Sci. 2003, 60, 648–662. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Nerurkar, A.S. Biological properties and thermal behavior of pectate lyase produced by Pectobacterium carotovorum subsp. carotovorum BR1 with industrial potentials. Biochem. Eng. J. 2012, 63, 22–30. [Google Scholar] [CrossRef]
- Souza, P.M.; Aliakbarian, B.; Ferreira, E.X.; Magalhaes, P.O.; Pessoa, A.; Converti, A.; Perego, P. Kinetic and thermodynamic studies of a novel acid protease from Aspergillus foetidus. Int. J. Biol. Macromol. 2015, 3, 17–21. [Google Scholar] [CrossRef]
- Mohapatra, B.R. Kinetic and thermodynamic properties of alginate lyase and cellulase co-produced by Exiguobacterium species Alg-S5. Int. J. Biol. Macromol. 2017, 98, 103–110. [Google Scholar] [CrossRef]
- Muley, A.B.; Chaudhari, S.A.; Singhal, R.S. Non-covalent conjugation of cutinase from Fusarium sp. ICT SAC1 with pectin for enhanced stability: Process minutiae, kinetics, thermodynamics and structural study. Int. J. Biol. Macromol. 2017, 102, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Buiarelli, F.; Di Filippo, P.; Riccardi, C.; Pomata, D.; Rumolo, E.; Giannetti, L.; Neri, B. Analytical method for the determination of mycotoxins in indoor/outdoor airborne particulate matter by HPLC-MS-MS. Int. J. Environ. Anal. Chem. 2015, 95, 713–729. [Google Scholar] [CrossRef]
- Popiel, D.; Koczyk, G.; Dawidziuk, A.; Gromadzka, K.; Blaszczyk, L.; Chelkowski, J. Zearalenone lactonohydrolase activity in Hypocreales and its evolutionary relationships within the epoxide hydrolase subset of a/b-hydrolases. BMC Microbiol. 2014, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.M.d.M.; Costa Batista, L.H.; Gouveia, M.J.; Leite, T.C.C.; de Mello, M.R.F.; de Assis, S.A.; de Sena, A.R. Kinetic and thermodynamic parameters, and partial characterization of the crude extract of tannase produced by Saccharomyces cerevisiae CCMB 520. Nat. Prod. Res. 2018, 32, 1068–1075. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [S] | V | X = V2 | Y = V2/[S] | VX | X2 | VY | XY | Y2 |
|---|---|---|---|---|---|---|---|---|
| 500 | 0.04 | 0.001663 | 3.33 × 10−6 | 6.78 × 10−5 | 2.76 × 10−6 | 1.356 × 10−7 | 5.529 × 10−9 | 1.106 × 10−11 |
| 1000 | 0.08 | 0.007156 | 7.16 × 10−6 | 0.61 × 10−4 | 5.12 × 10−5 | 6.053 × 10−7 | 5.120 × 10−8 | 5.120 × 10−11 |
| 3000 | 0.21 | 0.044701 | 1.49 × 10−5 | 0.0095 | 0.00199 | 3.150 × 10−6 | 6.660 × 10−7 | 2.220 × 10−10 |
| 5000 | 0.27 | 0.072798 | 1.46 × 10−5 | 0.0196 | 0.0053 | 3.928 × 10−6 | 1.059 × 10−6 | 2.119 × 10−10 |
| 7000 | 0.38 | 0.143907 | 2.06 × 10−5 | 0.0546 | 0.0207 | 7.798 × 10−6 | 2.958 × 10−6 | 4.226 × 10−10 |
| Σ | 0.0844 | 0.0281 | 1.561 × 10−5 | 4.741 × 10−6 | 9.189 × 10−10 | |||
| code | α | β | γ | δ | ε |
| [S] | [S] + Km0 | Vm[S] | f | f` | f2 | f`f | vf | vf` |
|---|---|---|---|---|---|---|---|---|
| 500 | 11,551.78 | 476.9 | 0.0413 | −3.574 × 10−6 | 0.0017 | 1.277 × 10−11 | −1.475 × 10−7 | 0.00168 |
| 1000 | 12,051.78 | 953.8 | 0.0791 | −6.567 × 10−6 | 0.0063 | 4.312 × 10−11 | −5.197 × 10−7 | 0.00669 |
| 3000 | 14,051.78 | 2861.4 | 0.2036 | −1.449 × 10−5 | 0.0415 | 2.100 × 10−10 | −2.951 × 10−6 | 0.04305 |
| 5000 | 16,051.78 | 4769 | 0.2971 | −1.851 × 10−5 | 0.0883 | 3.420 × 10−10 | −5.499 × 10−6 | 0.08016 |
| 7000 | 18,051.78 | 6676.6 | 0.3699 | −2.049 × 10−5 | 0.1368 | 4.198 × 10−10 | −7.578 × 10−6 | 0.14030 |
| Σ | 0.2745 | 1.028 × 10−9 | −1.669 × 10−5 | 0.27189 | ||||
| code | α` | β` | γ` | δ` | ε` |
| Temp. (°C) | Kcat (sec−1) | Ea (kJ × mol−1) | ΔH (kJ × mol−1) | ΔG (kJ × mol−1) | ΔS (kJ × mol−1 × K−1) |
|---|---|---|---|---|---|
| 25 | 0.2679 | 70.37 | 67.46 | 70.58 | −0.0104 |
| 30 | 0.2569 | 70.37 | 67.42 | 71.91 | −0.0148 |
| 35 | 0.2463 | 70.37 | 67.38 | 73.24 | −0.0190 |
| 40 | 0.2362 | 70.37 | 67.34 | 74.58 | −0.0231 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.; Xie, P.; Jin, J.; Jin, Q.; Wang, X. Kinetics, Thermodynamics and Mechanism of Enzymatic Degradation of Zearalenone in Degummed Corn Oil. Toxins 2023, 15, 19. https://doi.org/10.3390/toxins15010019
Zhao C, Xie P, Jin J, Jin Q, Wang X. Kinetics, Thermodynamics and Mechanism of Enzymatic Degradation of Zearalenone in Degummed Corn Oil. Toxins. 2023; 15(1):19. https://doi.org/10.3390/toxins15010019
Chicago/Turabian StyleZhao, Chenwei, Pengkai Xie, Jun Jin, Qingzhe Jin, and Xingguo Wang. 2023. "Kinetics, Thermodynamics and Mechanism of Enzymatic Degradation of Zearalenone in Degummed Corn Oil" Toxins 15, no. 1: 19. https://doi.org/10.3390/toxins15010019
APA StyleZhao, C., Xie, P., Jin, J., Jin, Q., & Wang, X. (2023). Kinetics, Thermodynamics and Mechanism of Enzymatic Degradation of Zearalenone in Degummed Corn Oil. Toxins, 15(1), 19. https://doi.org/10.3390/toxins15010019