Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Dose–Effect of Phloretin and LPS on the Viability of BRECs
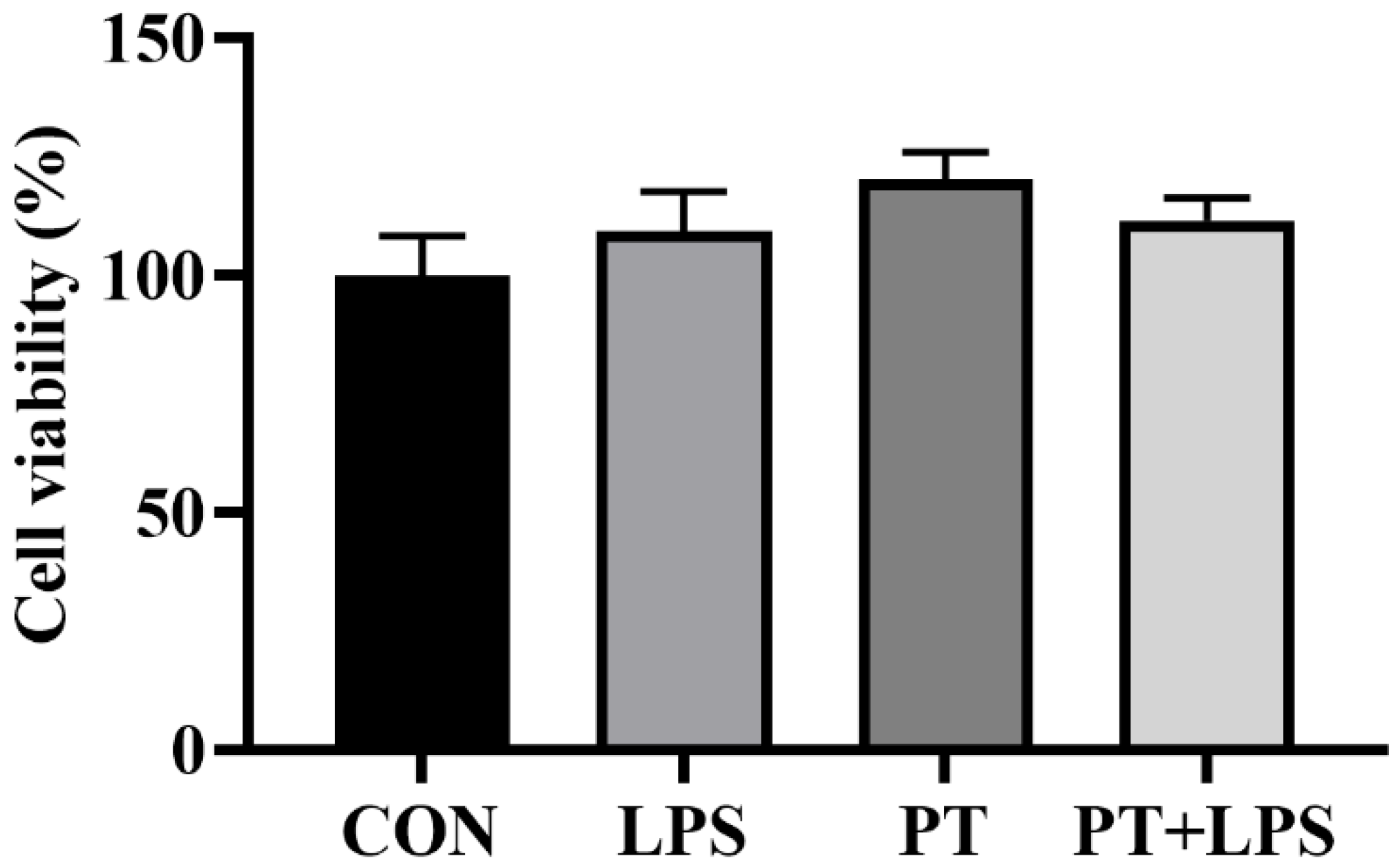
2.2. Effect of Phloretin on LPS-Induced Cell Viability
2.3. Effect of Phloretin on LPS-Induced Oxidative Properties
2.4. Effect of Phloretin on LPS-Induced Inflammatory Cytokine Gene Expression in BRECs
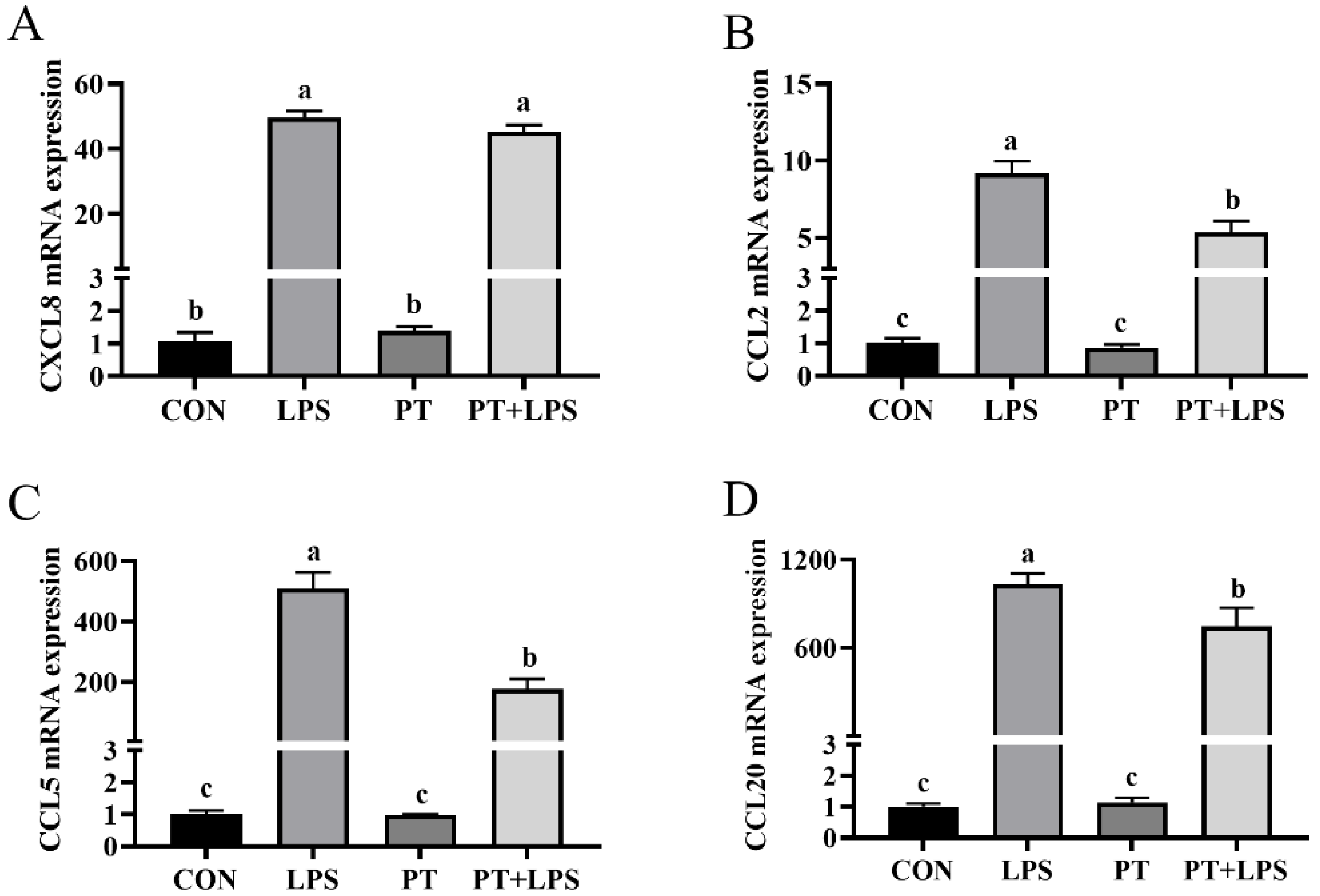
2.5. Effect of Phloretin on LPS-Induced Chemokine Gene Expression in BRECs
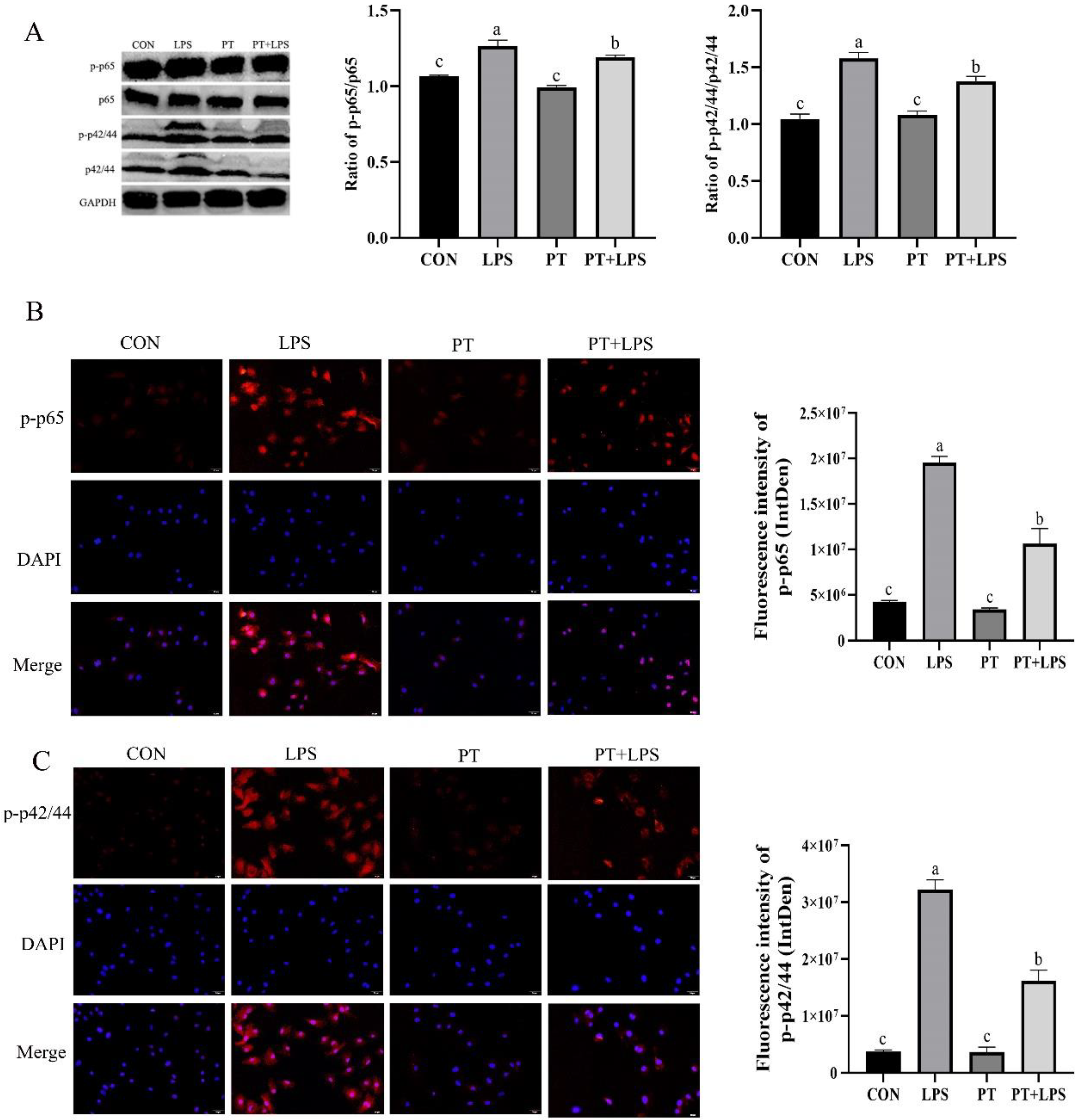
2.6. Effect of Phloretin on LPS-Induced the Expression of p-p65 and p-p42/44 in BRECs
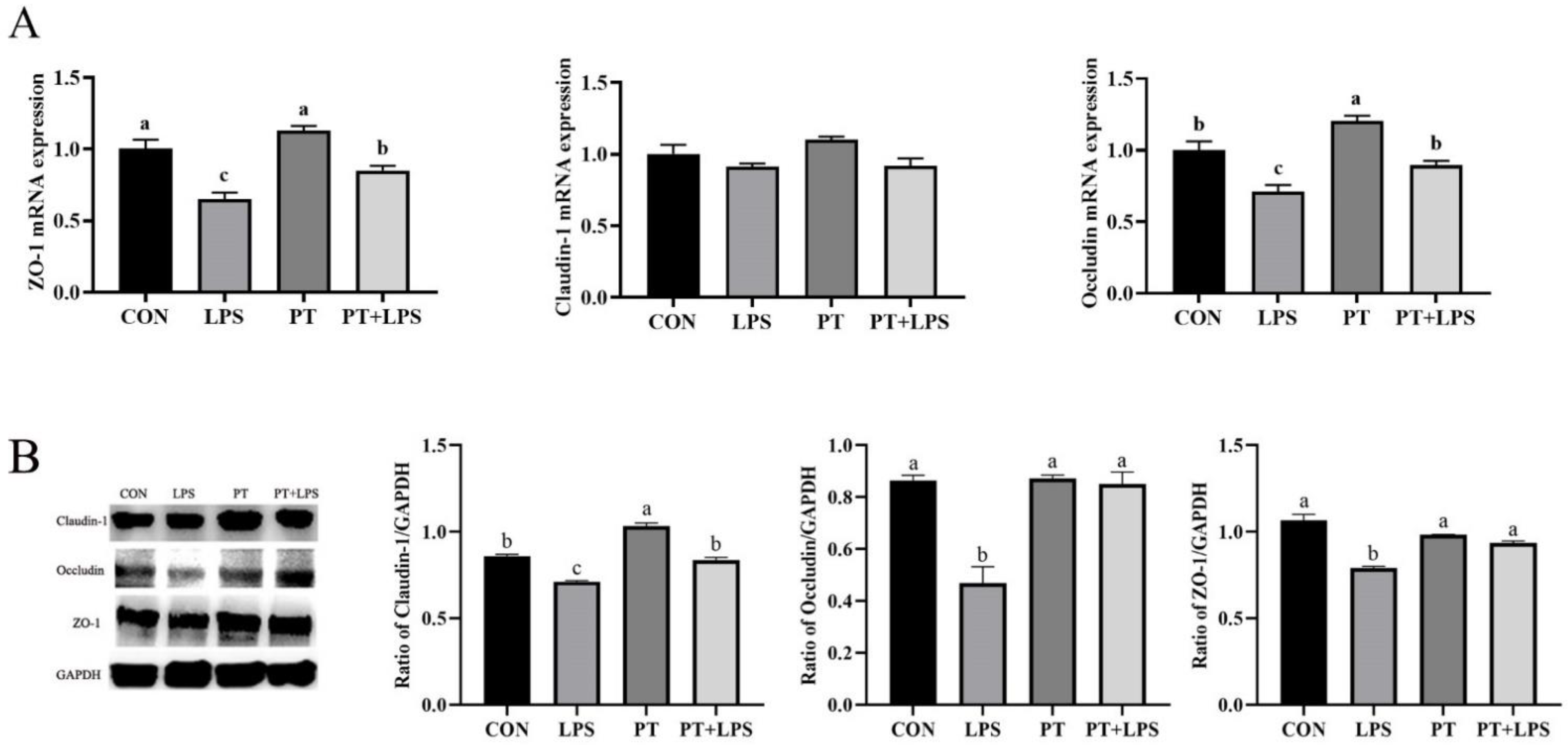
2.7. Effect of Phloretin on LPS-Induced Expression of Tight Junction Proteins in BRECs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Cell Culture
5.3. Treatment Methods for BRECs
5.4. Cell Viability Assay
5.5. Apoptosis Assay
5.6. Antioxidant Index Assay
5.7. RNA Extraction and Quantitative Real-Time PCR
5.8. Immunohistochemical Analysis of NF-κB and ERK1/2
5.9. Western Blotting
5.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, J.; Tian, K.; Sun, Y.; Wu, Y.; Chen, J.; Zhang, R.; He, T.; Dong, G. Effects of the acid–base treatment of corn on rumen fermentation and microbiota, inflammatory response and growth performance in beef cattle fed high-concentrate diet. Animal 2020, 14, 1876–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, K.; Liu, J.; Sun, Y.; Wu, Y.; Chen, J.; Zhang, R.; He, T.; Dong, G. Effects of dietary supplementation of inulin on rumen fermentation and bacterial microbiota, inflammatory response and growth performance in finishing beef steers fed high or low-concentrate diet. Anim. Feed Sci. Technol. 2019, 258, 114299. [Google Scholar] [CrossRef]
- Gozho, G.; Krause, D.; Plaizier, J. Ruminal lipopolysaccharide concentration and inflammatory response during grain-induced subacute ruminal acidosis in dairy cows. J. Dairy Sci. 2007, 90, 856–866. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Liu, S.; Wu, Y.; Lei, C.; Zhou, J.; Zhang, S. Diet-induced bacterial immunogens in the gastrointestinal tract of dairy cows: Impacts on immunity and metabolism. Acta Vet. Scand. 2011, 53, 48. [Google Scholar] [CrossRef] [Green Version]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [Green Version]
- Bertani, B.; Ruiz, N. Function and biogenesis of lipopolysaccharides. EcoSal Plus 2018, 8. [Google Scholar] [CrossRef]
- Aperce, C.; Amachawadi, R.; Van Bibber-Krueger, C.; Nagaraja, T.; Scott, H.; Vinasco-Torre, J.; Drouillard, J. Effects of menthol supplementation in feedlot cattle diets on the fecal prevalence of antimicrobial-resistant Escherichia coli. PLoS ONE 2016, 11, e0168983. [Google Scholar] [CrossRef] [Green Version]
- Steele, N.; Swartz, T.; Enger, K.; Schramm, H.; Cockrum, R.; Lacy-Hulbert, S.; White, R.; Hogan, J.; Petersson-Wolfe, C. The effect of J5 bacterins on clinical, behavioral, and antibody response following an Escherichia coli intramammary challenge in dairy cows at peak lactation. J. Dairy Sci. 2019, 102, 11233–11249. [Google Scholar] [CrossRef]
- Chae, B.S. Pretreatment of low-dose and super-low-dose LPS on the production of in vitro LPS-induced inflammatory mediators. Toxicol. Res. 2018, 34, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Mariadoss, A.V.; Vinyagam, R.; Rajamanickam, V.; Sankaran, V.; Venkatesan, S.; David, E. Pharmacological aspects and potential use of phloretin: A systemic review. Mini Rev. Med. Chem. 2019, 19, 1060–1067. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, J.; Memon, A.H.; Liang, H. Molecular model and in vitro antioxidant activity of a water-soluble and stable phloretin/hydroxypropyl-β-cyclodextrin inclusion complex. J. Mol. Liq. 2017, 236, 68–75. [Google Scholar] [CrossRef]
- Xu, M.; Gu, W.; Shen, Z.; Wang, F. Anticancer activity of phloretin against human gastric cancer cell lines involves apoptosis, cell cycle arrest, and inhibition of cell invasion and JNK signalling pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 6551. [Google Scholar] [CrossRef]
- Huang, W.-C.; Dai, Y.-W.; Peng, H.-L.; Kang, C.-W.; Kuo, C.-Y.; Liou, C.-J. Phloretin ameliorates chemokines and ICAM-1 expression via blocking of the NF-κB pathway in the TNF-α-induced HaCaT human keratinocytes. Int. Immunopharmacol. 2015, 27, 32–37. [Google Scholar] [CrossRef]
- Behzad, S.; Sureda, A.; Barreca, D.; Nabavi, S.F.; Rastrelli, L.; Nabavi, S.M. Health effects of phloretin: From chemistry to medicine. Phytochem. Rev. 2017, 16, 527–533. [Google Scholar] [CrossRef]
- Chauhan, A.; Jang, M.; Kim, Y. Phloretin Protects Macrophages from E. Coli-Induced Inflammation through the TLR4 Signaling Pathway. J. Microbiol. Biotechnol. 2020, 30, 333–340. [Google Scholar] [CrossRef]
- Eslamizad, M.; Albrecht, D.; Kuhla, B. The effect of chronic, mild heat stress on metabolic changes of nutrition and adaptations in rumen papillae of lactating dairy cows. J. Dairy Sci. 2020, 103, 8601–8614. [Google Scholar] [CrossRef]
- Jung, M.; Triebel, S.; Anke, T.; Richling, E.; Erkel, G. Influence of apple polyphenols on inflammatory gene expression. Mol. Nutr. Food Res. 2010, 53, 1263–1280. [Google Scholar] [CrossRef]
- Wu, M.; Li, P.; An, Y.; Ren, J.; Yan, D.; Cui, J.; Li, D.; Li, M.; Wang, M.; Zhong, G. Phloretin ameliorates dextran sulfate sodium-induced ulcerative colitis in mice by regulating the gut microbiota. Pharmacol. Res. 2019, 150, 104489. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.S.; Kwon, J.Y.; Kang, N.J.; Lee, K.W.; Lee, H.J. Phloretin Induces Apoptosis in H-Ras MCF10A Human Breast Tumor Cells through the Activation of p53 via JNK and p38 Mitogen-Activated Protein Kinase Signaling. Ann. N. Y. Acad. Sci. 2010, 1171, 479–483. [Google Scholar] [CrossRef]
- Plaizier, J.; Krause, D.; Gozho, G.; McBride, B. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Li, X.; Guan, Y.; Wang, Y.; Yuan, X.; Sun, G.; Wang, Z.; Li, X. Inflammatory mechanism of Rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef]
- Mao, S.Y.; Huo, W.J.; Zhu, W.Y. Microbiome–metabolome analysis reveals unhealthy alterations in the composition and metabolism of ruminal microbiota with increasing dietary grain in a goat model. Environ. Microbiol. 2016, 18, 525–541. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Zebeli, Q.; Patra, A.K.; Greco, G.; Amasheh, S.; Penner, G.B. Symposium review: The importance of the ruminal epithelial barrier for a healthy and productive cow. J. Dairy Sci. 2019, 102, 1866–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [PubMed]
- Djordjevic, V.B. Free radicals in cell biology. Int. Rev. Cytol. 2004, 237, 57–89. [Google Scholar] [PubMed]
- Hu, Q.; Liu, Z.; Guo, Y.; Lu, S.; Du, H.; Cao, Y. Antioxidant capacity of flavonoids from Folium Artemisiae Argyi and the molecular mechanism in Caenorhabditis elegans. J. Ethnopharmacol. 2021, 279, 114398. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, S.; Huang, J.; Cui, Y.; Liu, Y.; Zhou, Y.; Zhu, Z. Phloretin is protective in a murine salmonella enterica serovar typhimurium infection model. Microb. Pathog. 2021, 161, 105298. [Google Scholar] [CrossRef]
- Shanmugam, N.; Reddy, M.A.; Guha, M.; Natarajan, R. High glucose-induced expression of proinflammatory cytokine and chemokine genes in monocytic cells. Diabetes 2003, 52, 1256–1264. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Fu, X.; Liao, X.; Li, Y. Nrf2 activators as dietary phytochemicals against oxidative stress, inflammation, and mitochondrial dysfunction in autism spectrum disorders: A systematic review. Front. Psychiatry 2020, 11, 1299. [Google Scholar] [CrossRef]
- Rallabhandi, P.; Awomoyi, A.; Thomas, K.E.; Phalipon, A.; Fujimoto, Y.; Fukase, K.; Kusumoto, S.; Qureshi, N.; Sztein, M.B.; Vogel, S.N. Differential activation of human TLR4 by Escherichia coli and Shigella flexneri 2a lipopolysaccharide: Combined effects of lipid A acylation state and TLR4 polymorphisms on signaling. J. Immunol. 2008, 180, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Jacob, B.; Jang, M.; Kwak, C.; Lee, Y.; Son, K.; Lee, S.; Jung, I.D.; Jeong, M.S.; Kwon, S.-H. Development of a novel short 12-meric papiliocin-derived peptide that is effective against Gram-negative sepsis. Sci. Rep. 2019, 9, 3817. [Google Scholar] [CrossRef] [Green Version]
- De Schepper, S.; De Ketelaere, A.; Bannerman, D.D.; Paape, M.J.; Peelman, L.; Burvenich, C. The toll-like receptor-4 (TLR-4) pathway and its possible role in the pathogenesis of Escherichia coli mastitis in dairy cattle. Vet. Res. 2008, 39, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Riollet, C.; Rainard, P.; Poutrel, B. Differential induction of complement fragment C5a and inflammatory cytokines during intramammary infections with Escherichia coli and Staphylococcus aureus. Clin. Diagn. Lab. Immunol. 2000, 7, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Günther, J.; Koczan, D.; Yang, W.; Nürnberg, G.; Repsilber, D.; Schuberth, H.-J.; Park, Z.; Maqbool, N.; Molenaar, A.; Seyfert, H.-M. Assessment of the immune capacity of mammary epithelial cells: Comparison with mammary tissue after challenge with Escherichia coli. Vet. Res. 2009, 40, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kayisoglu, O.; Weiss, F.; Niklas, C.; Pierotti, I.; Pompaiah, M.; Wallaschek, N.; Germer, C.-T.; Wiegering, A.; Bartfeld, S. Location-specific cell identity rather than exposure to GI microbiota defines many innate immune signalling cascades in the gut epithelium. Gut 2021, 70, 687–697. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, W.; Mao, S. High-concentrate feeding upregulates the expression of inflammation-related genes in the ruminal epithelium of dairy cattle. J. Anim. Sci. Biotechnol. 2016, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Fang, H.; Shen, J.; Jin, Y.; Zhang, J. Curcumin Alleviates LPS-Induced Oxidative Stress, Inflammation and Apoptosis in Bovine Mammary Epithelial Cells via the NFE2L2 Signaling Pathway. Toxins 2021, 13, 208. [Google Scholar] [CrossRef]
- Huang, W.-C.; Lai, C.-L.; Liang, Y.-T.; Hung, H.-C.; Liu, H.-C.; Liou, C.-J. Phloretin attenuates LPS-induced acute lung injury in mice via modulation of the NF-κB and MAPK pathways. Int. Immunopharmacol. 2016, 40, 98–105. [Google Scholar] [CrossRef]
- Porta, C.; Ippolito, A.; Consonni, F.M.; Carraro, L.; Celesti, G.; Correale, C.; Grizzi, F.; Pasqualini, F.; Tartari, S.; Rinaldi, M. Protumor steering of cancer inflammation by p50 NF-κB enhances colorectal cancer progression. Cancer Immunol. Res. 2018, 6, 578–593. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, P. Expressions and clinicopathologic significance of Id2 and NF-κB/P65 in gastric cancer. Zhonghua Yi Xue Za Zhi 2018, 98, 846–850. [Google Scholar]
- Shi, H.; Berger, E.A. Characterization of site-specific phosphorylation of NF-κB p65 in retinal cells in response to high glucose and cytokine polarization. Mediat. Inflamm. 2018, 2018, 3020675. [Google Scholar] [CrossRef] [Green Version]
- Birru, R.L.; Bein, K.; Wells, H.; Bondarchuk, N.; Barchowsky, A.; Di, Y.P.; Leikauf, G.D. Phloretin, an Apple Polyphenol, Inhibits Pathogen-Induced Mucin Overproduction. Mol. Nutr. Food Res. 2021, 65, 2000658. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Cao, H.; Shen, P.; Liu, J.; Fu, Y.; Cao, Y.; Zhang, N. The protective role of phloretin against dextran sulfate sodium-induced ulcerative colitis in mice. Food Funct. 2019, 10, 422–431. [Google Scholar] [CrossRef]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- You, Q.; Xu, J.; Zhu, Z.; Hu, Z.; Cai, Q. Phloretin flavonoid exhibits selective antiproliferative activity in doxorubicin-resistant gastric cancer cells by inducing autophagy, inhibiting cell migration and invasion, cell cycle arrest and targeting ERK1/2 MAP pathway. J. BUON 2020, 25, 308–313. [Google Scholar]
- Shen, H.; Xu, Z.; Shen, Z.; Lu, Z. The regulation of ruminal short-chain fatty acids on the functions of rumen barriers. Front. Physiol. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.-C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol improves barrier function and attenuates inflammatory responses in porcine intestinal epithelial cells during lipopolysaccharide (LPS)-induced inflammation. J. Agric. Food Chem. 2018, 67, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol. 2001, 2, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, H.; Zhu, W.; Mao, S. Dynamic changes in rumen fermentation and bacterial community following rumen fluid transplantation in a sheep model of rumen acidosis: Implications for rumen health in ruminants. FASEB J. 2019, 33, 8453–8467. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, T.; Liu, Y.; Zhu, W.; Mao, S. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Ni, X.; Jin, C.; Ren, W.; Li, J.; Deng, J.; Deng, B.; Yin, Y. Cecropin A modulates tight junction-related protein expression and enhances the barrier function of porcine intestinal epithelial cells by suppressing the MEK/ERK pathway. Int. J. Mol. Sci. 2018, 19, 1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, K.; Gong, X.; Chen, Y.; Jiang, M.; Yang, T.; Zhao, G. Short-chain fatty acids regulate the immune responses via G protein-coupled receptor 41 in bovine rumen epithelial cells. Front. Immunol. 2019, 10, 2042. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Liu, X.; Cheng, L.; Wang, D.; Qin, G.; Zhang, X.; Zhen, Y.; Wang, T.; Sun, Z. Protective Mechanism of Leucine and Isoleucine against H2O2-Induced Oxidative Damage in Bovine Mammary Epithelial Cells. Oxidative Med. Cell. Longev. 2022, 2022, 4013575. [Google Scholar] [CrossRef]
- Gong, X.; Su, X.; Zhan, K.; Zhao, G. The protective effect of chlorogenic acid on bovine mammary epithelial cells and neutrophil function. J. Dairy Sci. 2018, 101, 10089–10097. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size (bp) | Accession Number |
|---|---|---|---|
| CXCL8 | F: TGGGCCACACTGTGAAAAT | 136 | NM_173925.2 |
| R: TCATGGATCTTGCTTCTCAGC | |||
| CCL20 | F: TTCGACTGCTGTCTCCGATA | 172 | NM_174263.2 |
| R: GCACAACTTGTTTCACCCACT | |||
| CCL2 | F: GCTCGCTCAGCCAGATGCAA | 117 | NM_174006.2 |
| R: GGACACTTGCTGCTGGTGACTC | |||
| CCL5 | F: CTGCCTTCGCTGTCCTCCTGATG | 217 | NM_175827.2 |
| R: TTCTCTGGGTTGGCGCACACCTG | |||
| TNF-α | F: GCCCTCTGGTTCAGACACTC | 192 | NM_173966.3 |
| R: AGATGAGGTAAAGCCCGTCA | |||
| IL-6 | F: TCCTTGCTGCTTTCACACTC | 129 | NM_173923.2 |
| R: CACCCCAGGCAGACTACTTC | |||
| IL-1β | F: CAGTGCCTACGCACATGTCT | 209 | NM_174093.1 |
| R: AGAGGAGGTGGAGAGCCTTC | |||
| IL-8 | F: TGGGCCACACTGTGAAAAT | 136 | NM_173925.2 |
| R: TCATGGATCTTGCTTCTCAGC | |||
| TLR-4 | F: GACCCTTGCGTACAGGTTGT | 103 | NM_174198.6 |
| R: GGTCCAGCATCTTGGTTGAT | |||
| TLR-2 | F: CAGGCTTCTTCTCTGTCTTGT | 140 | NM_174197.2 |
| R: CTGTTGCCGACATAGGTGATA | |||
| ZO-1 | F: TCTGCAGCAATAAAGCAGCATTTC | 187 | XM_010817146.1 |
| R: TTAGGGCACAGCATCGTATCACA | |||
| Claudin-1 | F: CGTGCCTTGATGGTGAT | 102 | NM_001001854.2 |
| R: CTGTGCCTCGTCGTCTT | |||
| Occludin | F: GAACGAGAAGCGACTGTATC | 122 | NM_001082433.2 |
| R: CACTGCTGCTGTAATGAGG | |||
| GAPDH | F: GGGTCATCATCTCTGCACCT | 176 | NM_001034034.2 |
| R: GGTCATAAGTCCCTCCACGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Lei, Q.; Ma, H.; Jiang, M.; Yang, T.; Ma, Q.; Datsomor, O.; Zhan, K.; Zhao, G. Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury. Toxins 2022, 14, 337. https://doi.org/10.3390/toxins14050337
Wang K, Lei Q, Ma H, Jiang M, Yang T, Ma Q, Datsomor O, Zhan K, Zhao G. Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury. Toxins. 2022; 14(5):337. https://doi.org/10.3390/toxins14050337
Chicago/Turabian StyleWang, Kexin, Qian Lei, Huimin Ma, Maocheng Jiang, Tianyu Yang, Qianbo Ma, Osmond Datsomor, Kang Zhan, and Guoqi Zhao. 2022. "Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury" Toxins 14, no. 5: 337. https://doi.org/10.3390/toxins14050337
APA StyleWang, K., Lei, Q., Ma, H., Jiang, M., Yang, T., Ma, Q., Datsomor, O., Zhan, K., & Zhao, G. (2022). Phloretin Protects Bovine Rumen Epithelial Cells from LPS-Induced Injury. Toxins, 14(5), 337. https://doi.org/10.3390/toxins14050337