Cytotoxicity and Effects on the Synapsis Induced by Pure Cylindrospermopsin in an E17 Embryonic Murine Primary Neuronal Culture in a Concentration- and Time-Dependent Manner

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
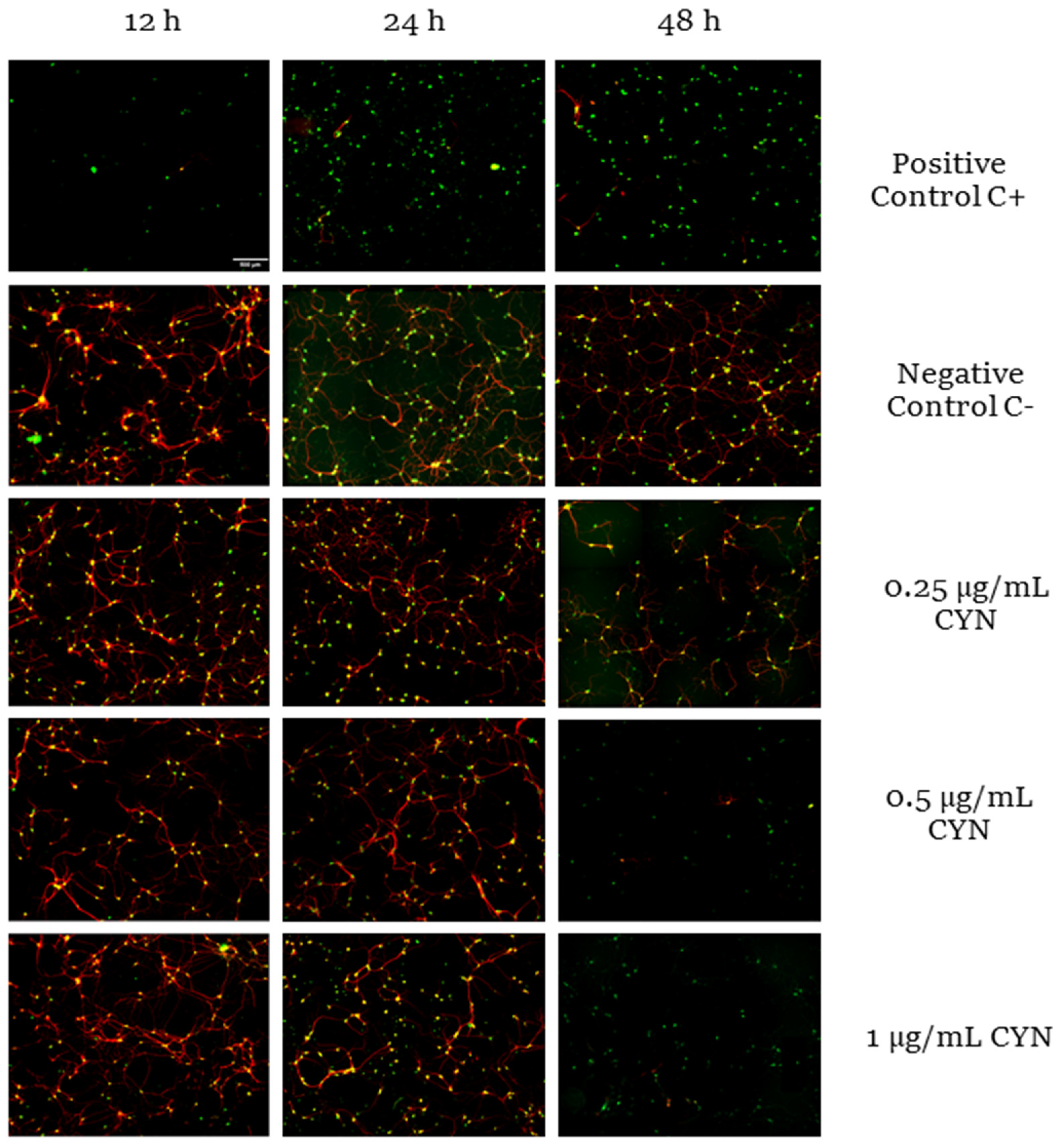
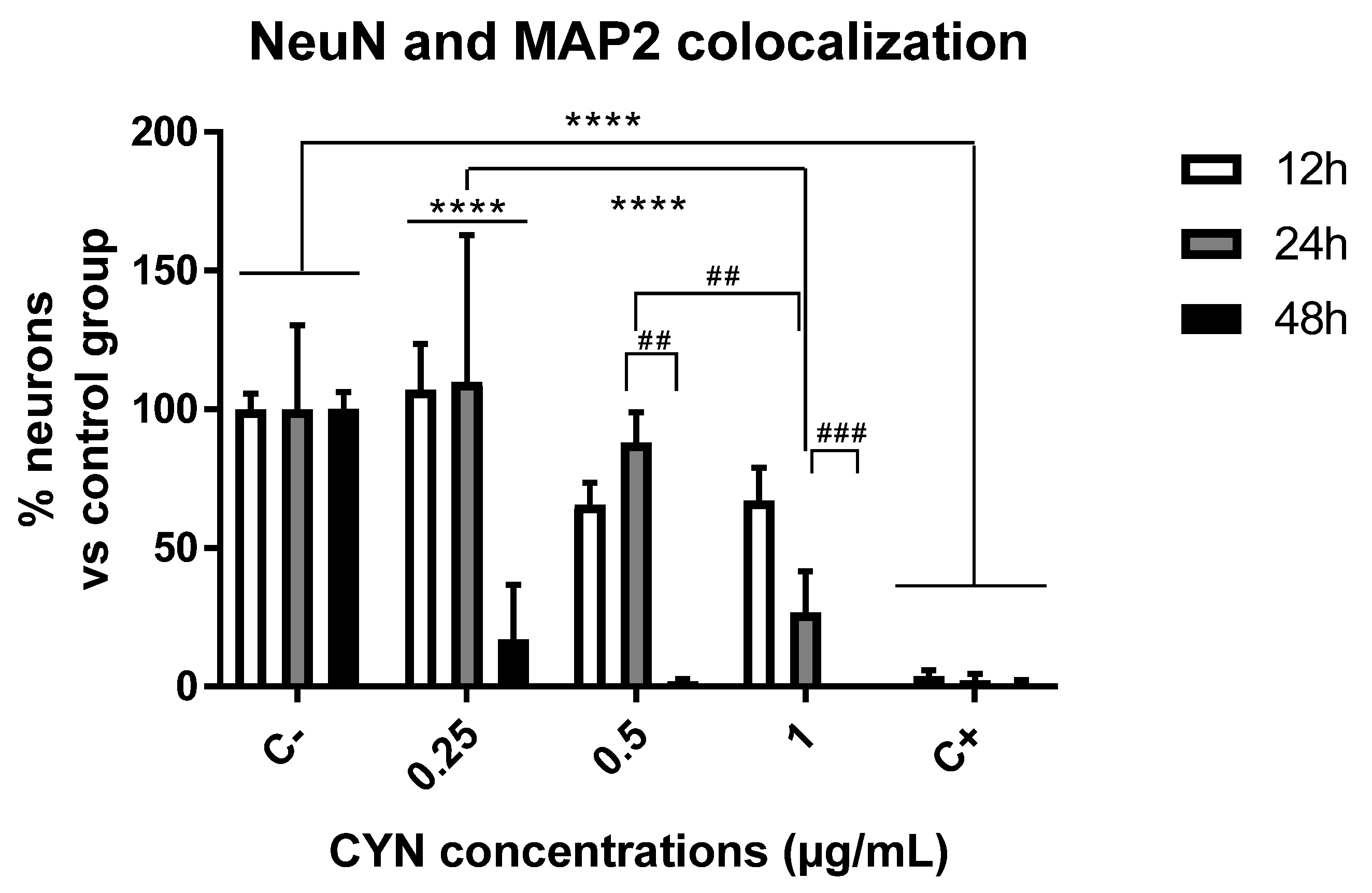
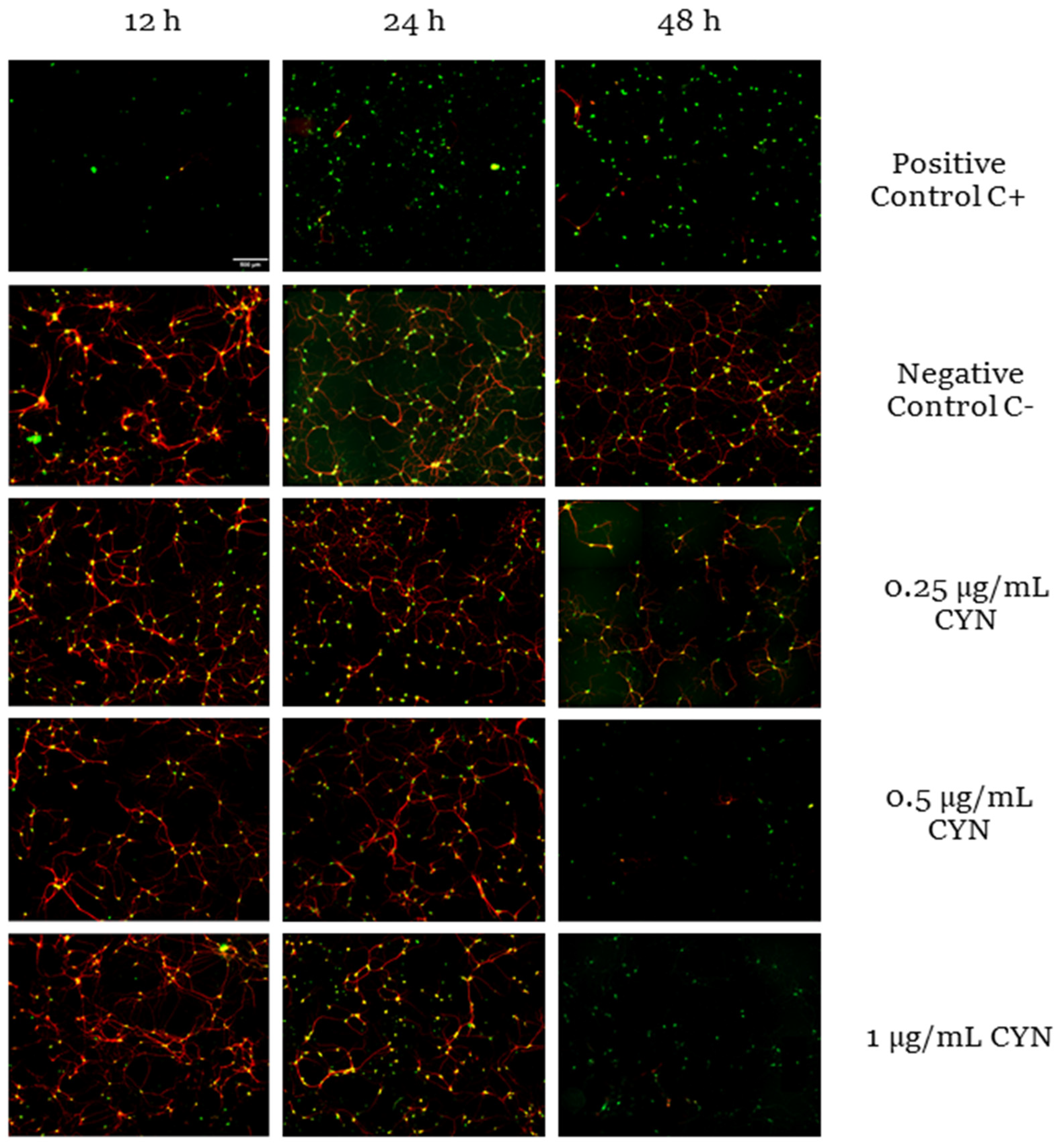
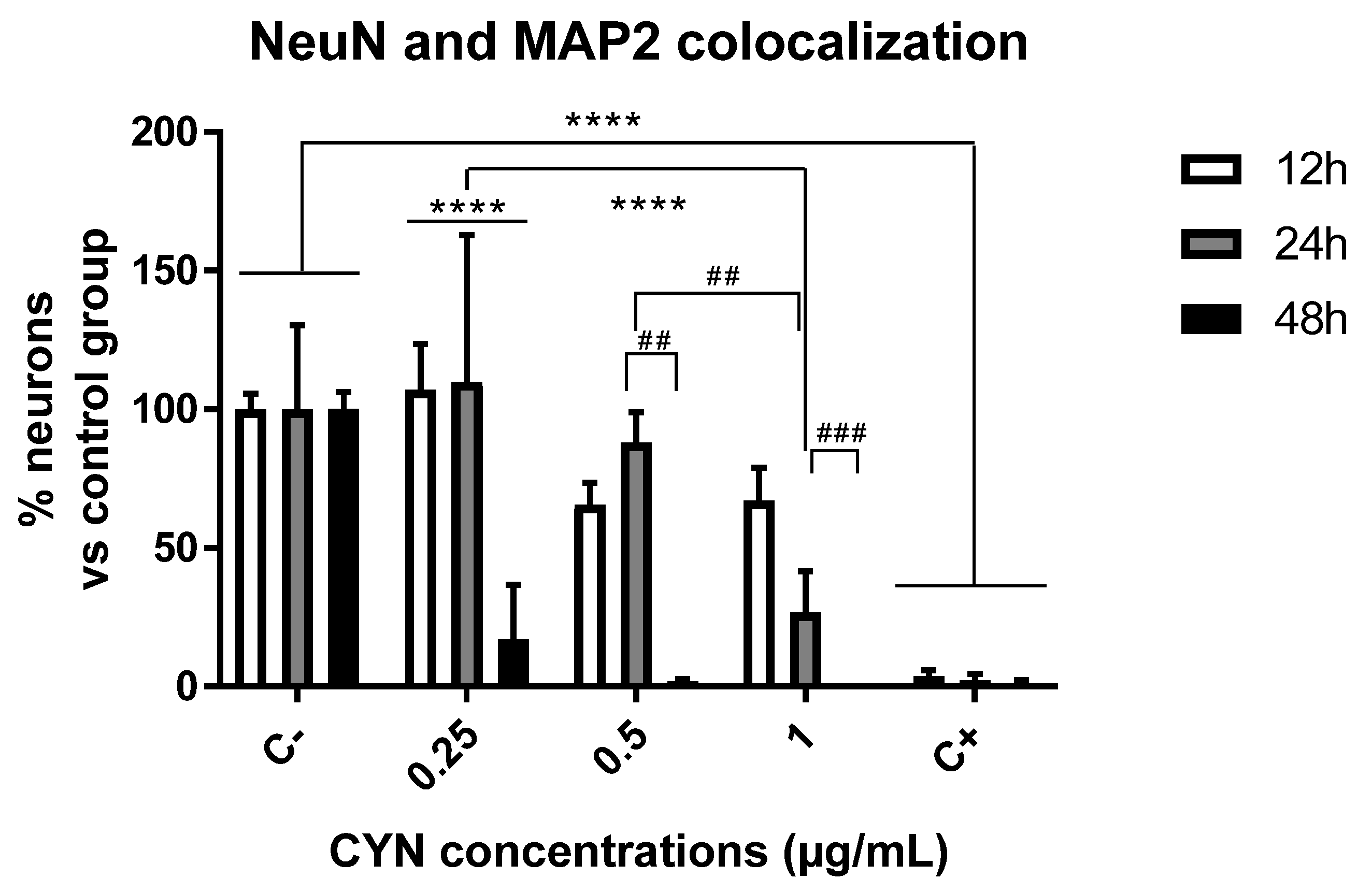
2.1. Cytotoxicity of CYN in Murine Primary Neuronal Culture
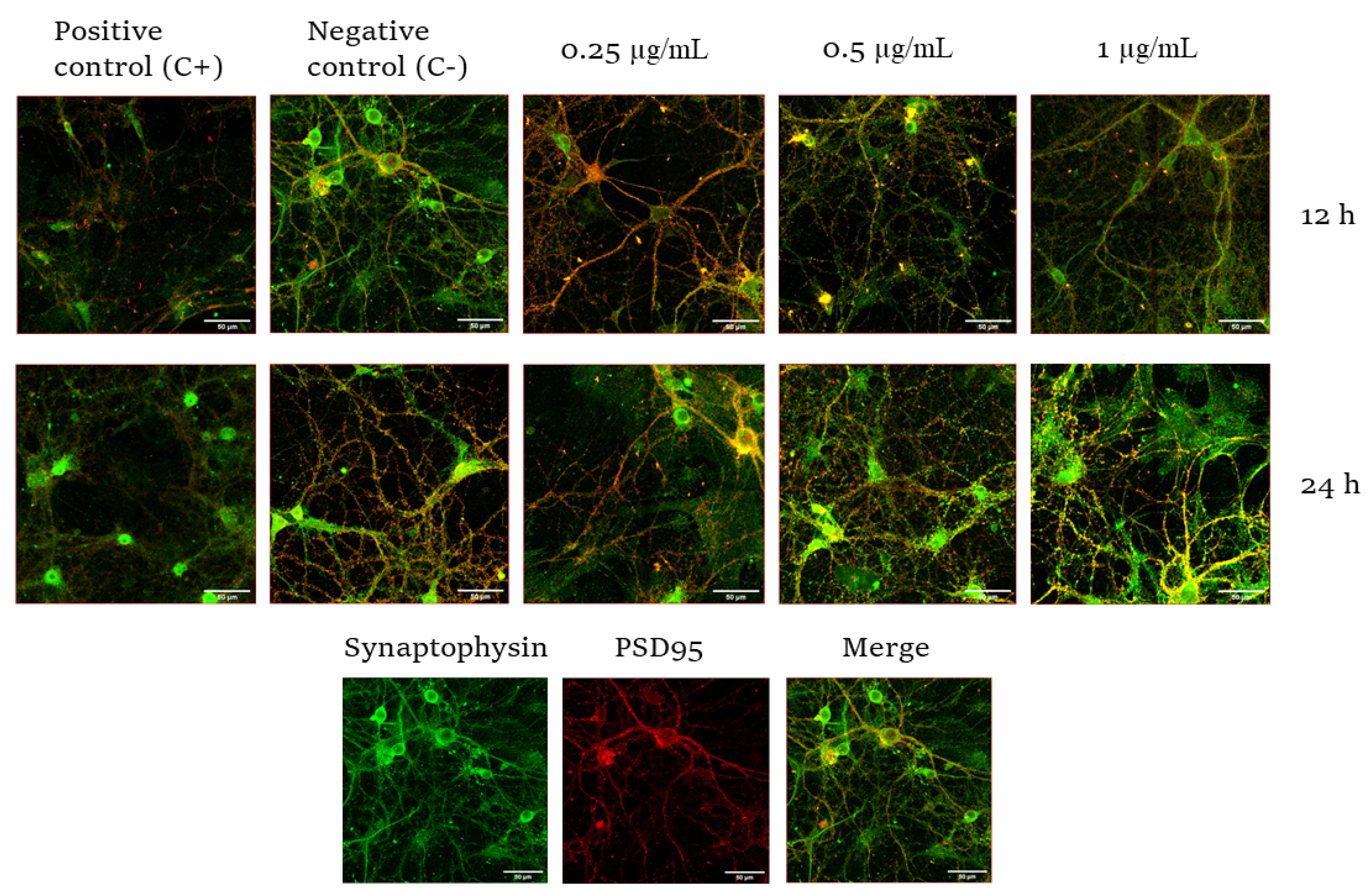
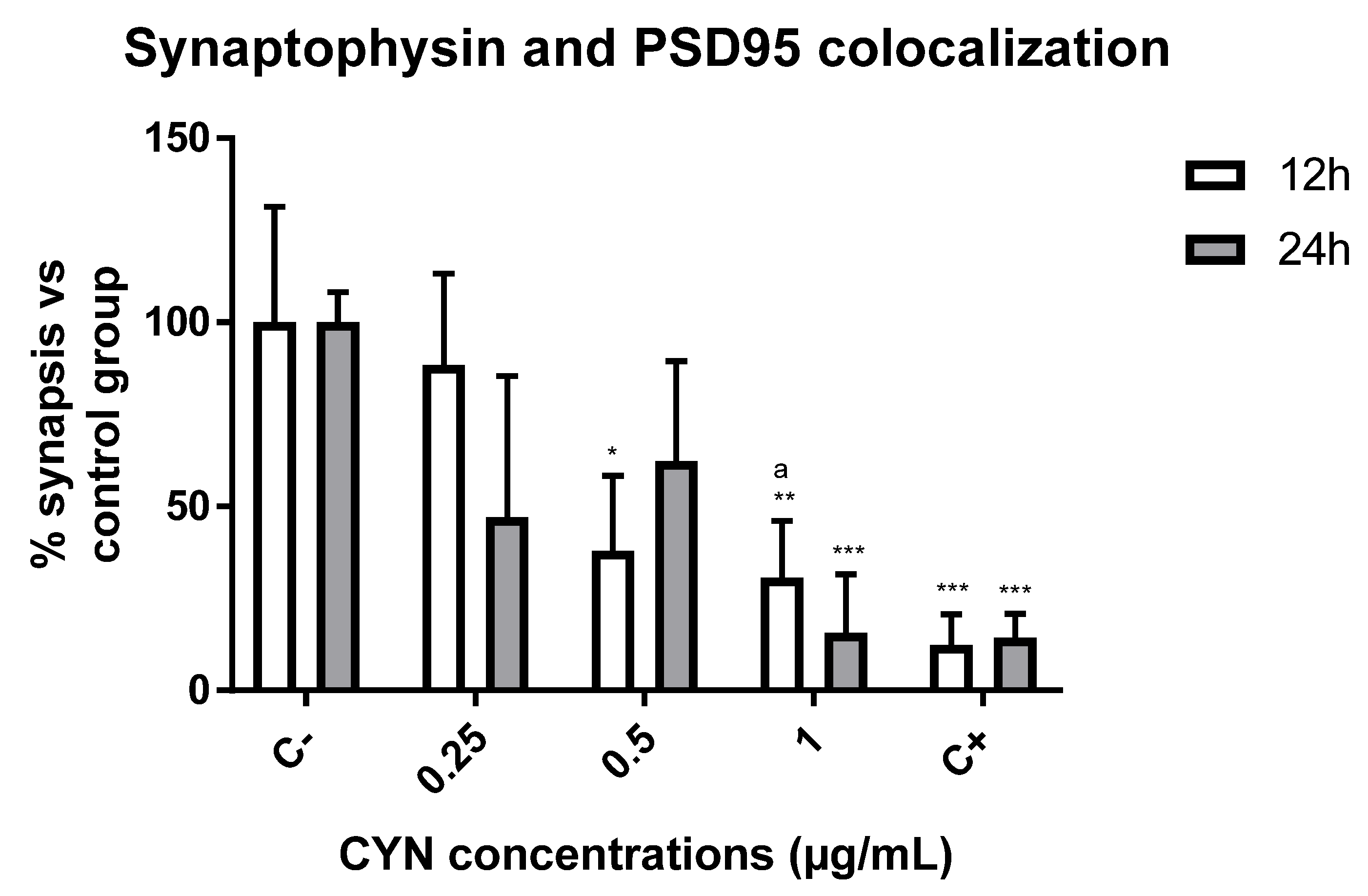
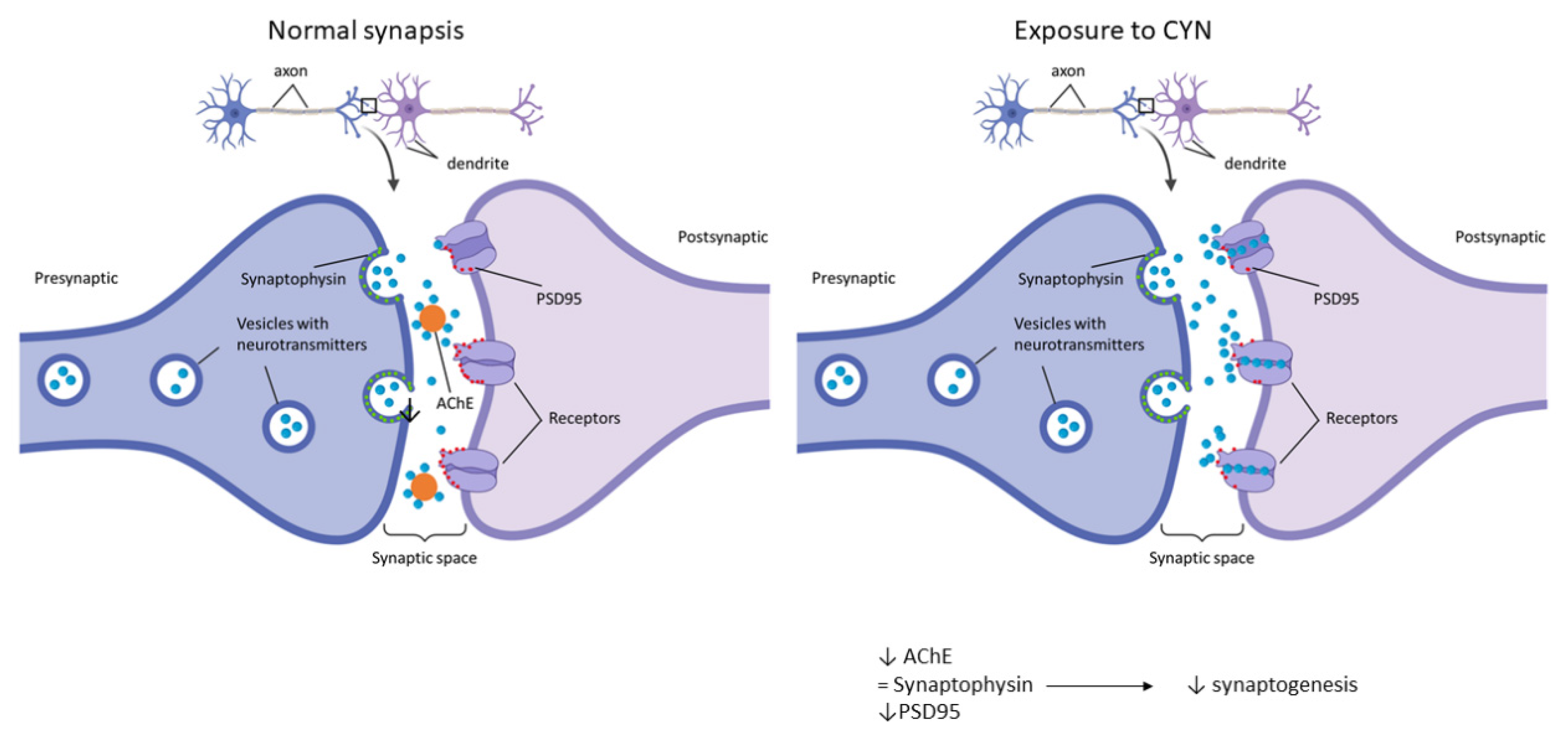
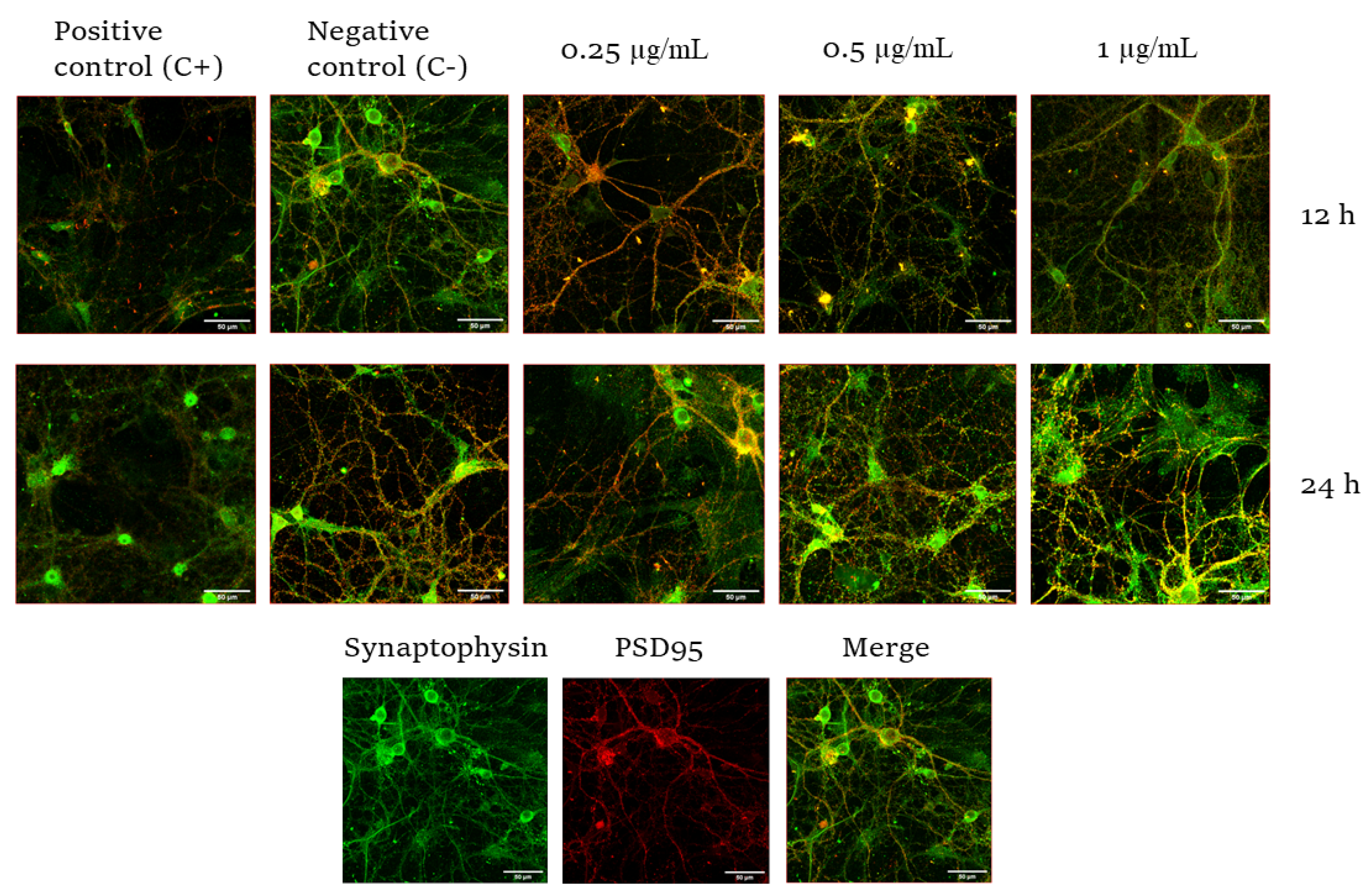
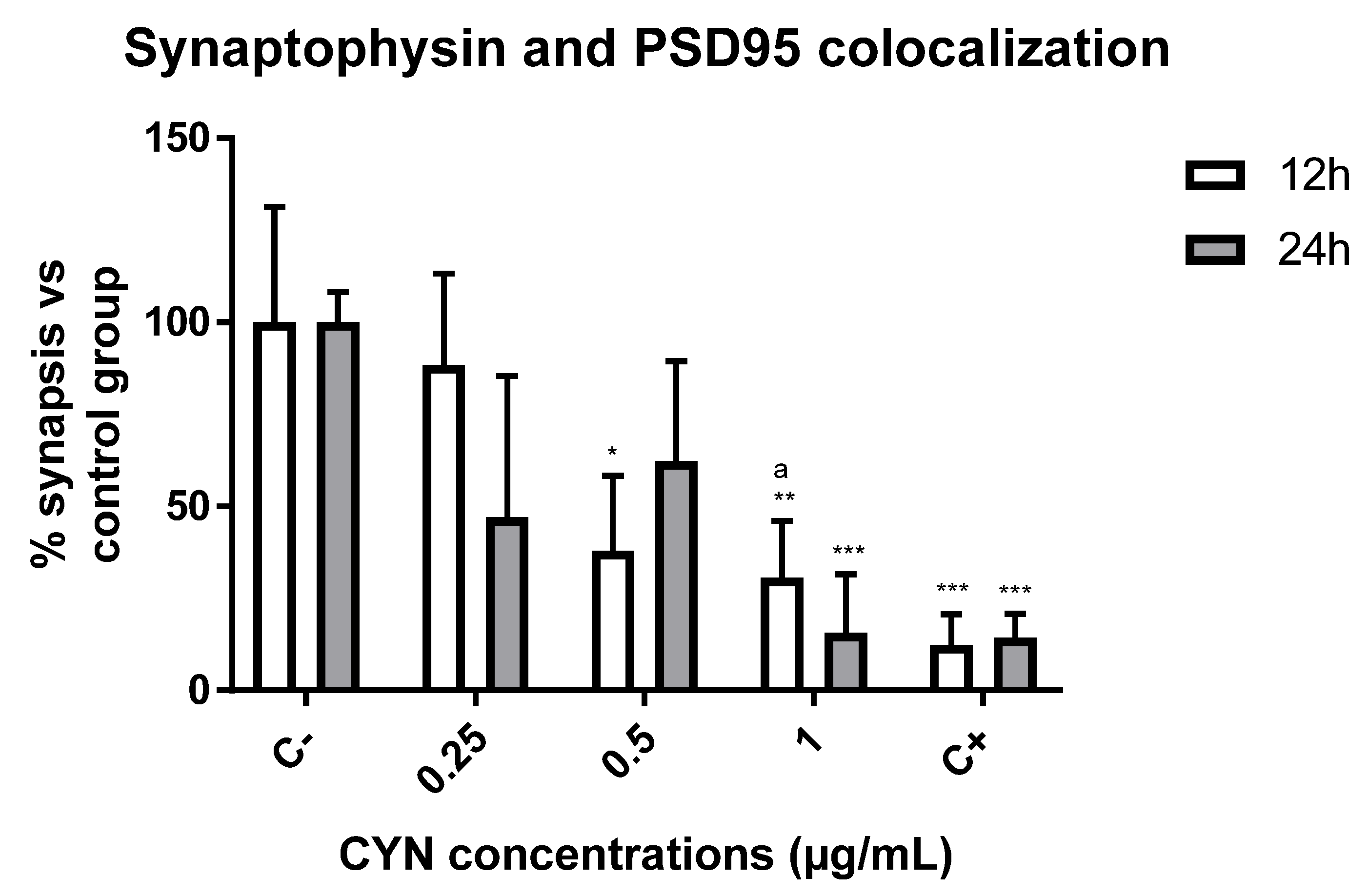
2.2. Effect of CYN on the Number of Synapses in Murine Primary Neuronal Culture
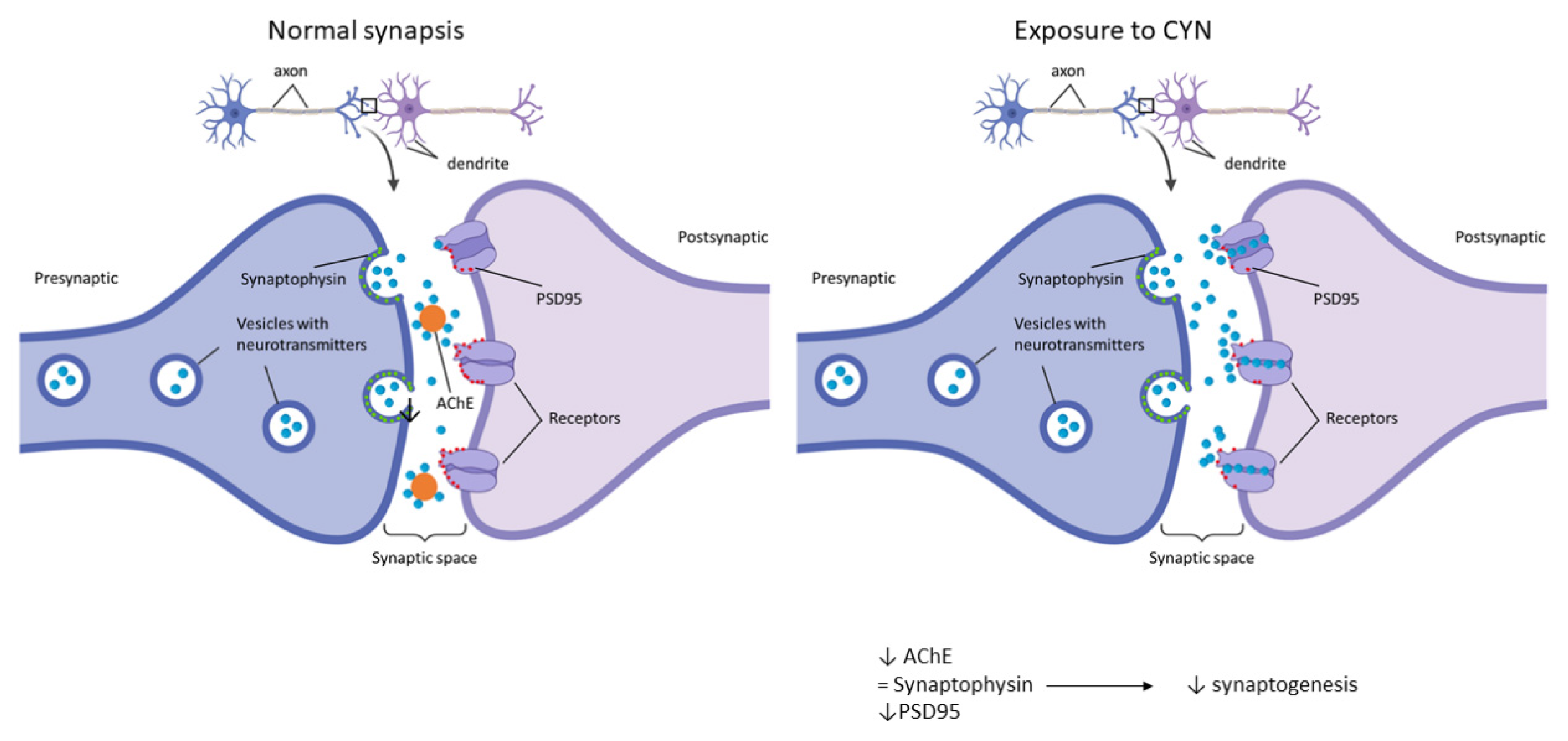
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Toxin Test Solution
5.3. Animals
5.4. Primary Culture of Hippocampal Neurons and Toxin Exposure
5.5. Sample Fixation and Immunocytochemistry
5.6. Image Acquisition and Analysis
5.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Guzmán-Guillén, R.; Prieto Ortega, A.I.; Moreno, I.; González, G.; Soria-Díaz, M.E.; Vasconcelos, V.; Cameán, A.M. Development and optimization of a method for the determination of Cylindrospermopsin production from strain of Aphanizomenon cultures: Intra-laboratory assessment of its accuracy by using validation standards. Talanta 2012, 100, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Cameán, A.M.; Carmeli, S.; Guzmán-Guillén, R.; Jos, A.; Mankiewicz-Boczek, J.; Metcalf, J.S.; Moreno, I.M.; Prieto, A.I.; Sukenik, A. Cylindrospermopsin and Congeners. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 127–137. [Google Scholar]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium. Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar]
- Kinnear, S. Cylindrospermopsin: A decade of progress on bioaccumulation research. Mar. Drugs 2010, 8, 542–564. [Google Scholar] [CrossRef] [Green Version]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, G.; Chen, Y.; Jia, N.; Li, R. Four decades of progress in cylindrospermopsin research: The ins and outs of a potent cyanotoxin. J. Hazard. Marter. 2021, 406, 124653. [Google Scholar] [CrossRef]
- Otero, P.; Silva, M. Chapter 7—The role of toxins: Impact on human health and aquatic environments. In The Pharmacological Potential of Cyanobacteria; Lopes, G., Silva, M., Vasconcelos, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 173–199. [Google Scholar]
- Mutoti, M.; Gumbo, J.; Jideani, A.I.O. Occurrence of cyanobacteria in water used for food production: A review. Phys. Chem. Earth Parts A/B/C 2022, 125, 103101. [Google Scholar] [CrossRef]
- Ríos, V.; Moreno, I.; Prieto, A.I.; Puerto, M.; Gutiérrez-Praena, D.; Soria, M.E.; Cameán, A.M. Analysis of MC-LR and MC-RR in tissue from freshwater fish (Tinca tinca) and crayfish (Procambarus clarkii) in tench ponds (Cáceres, Spain) by Liquid Chromatography-Mass spectrometry (LC-MS). Food Chem. Toxicol. 2013, 57, 170–178. [Google Scholar] [CrossRef]
- Guzmán-Guillen, R.; Moreno, I.; Prieto, A.I.; Soria-Diaz, M.E.; Vasconcelos, V.M.; Cameán, A.M. CYN determination in tissues from freshwater fish by LC-MS/MS: Validation and application in tissues form subchronically exposed tilapia (Oreochromis niloticus). Talanta 2015, 131, 452–459. [Google Scholar] [CrossRef]
- Gutiérrez-Praena, D.; Jos, Á.; Pichardo, S.; Moreno, I.M.; Cameán, A.M. Presence and bioaccumulation of microcystins and cylindrospermopsin in food and the effectiveness of some cooking techniques at decreasing their concentrations: A review. Food Chem. Toxicol. 2013, 53, 139–152. [Google Scholar] [CrossRef]
- Codd, G.A.; Meriluoto, J.; Metcalf, J.S. Introduction: Cyanobacteria, cyanotoxins, their human impact, and risk management. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 3–8. [Google Scholar]
- Byth, S. Palm Island mystery disease. Med. J. Aust. 1980, 2, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Praena, D.; Pichardo, S.; Jos, Á.; Moreno, J.; Cameán, A.M. Alterations observed in the endothelial HUVEC cell line exposed to pure Cylindrospermopsin. Chemosphere 2012, 89, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Pichardo, S.; Cameán, A.M.; Jos, A. In vitro toxicological assessment of cylindropermopsin: A review. Toxins 2017, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichardo, S.; Devesa, V.; Puerto, M.; Vélez, D.; Cameán, A.M. Intestinal transport of Cylindrospermopsin using the Caco-2 cell line. Toxicol. In Vitro 2017, 38, 142–149. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron-microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green-alga Umezakia Natans. Toxicon 1994, 32, 833–843. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Prieto Ortega, A.I.; Moreno, I.M.; Ríos, V.; Moyano, R.; Blanco, A.; Vasconcelos, V.; Cameán, A.M. Effects of Depuration on Histopathological Changes in Tilapia (Oreochromis Niloticus) After Exposure to Cylindrospermopsin. Environmental Toxicology 2016, 1318–1332. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Lomares, I.; Moreno, I.M.; Prieto, A.I.; Moyano, R.; Blanco, A.; Cameán, A.M. Cylindrospermopsin induces neurotoxicity in tilapia fish (Oreochromis niloticus) exposed to Aphanizomenon ovalisporum. Aquat. Toxicol. 2015, 161, 17–24. [Google Scholar] [CrossRef]
- Da Silva, R.C.; Grötzner, S.R.; Moura Costa, D.D.; Esquivel, J.R.; Muelbert, J.; Freitas de Magalhães, V.; Filipak, F.; De Oliveira, C.A. Comparative bioaccumulation and effects of purified and cellular extract of cylindrospermopsin to freshwater fish Hoplias malabaricus. J. Toxicol. Environ. Health Part A Curr. Issues 2018, 81, 620–632. [Google Scholar] [CrossRef]
- Rabelo, J.C.S.; Hanusch, A.L.; De Jesus, L.W.O.; Mesquita, L.A.; Franco, F.C.; Silva, R.A.; Sabóia-Morais, S.M.T. DNA damage induced by cylindrospermopsin on different tissues of the biomonitor fish Poecilia reticulate. Environ. Toxicol. 2021, 36, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, N.; Hill, D.J.; Chorus, I.; Diggs, D.L.; Huang, H.; King, D.; Lang, J.R.; Le, T.-T.; Schmid, J.E.; Travlos, G.S.; et al. Cylindrospermopsin toxicity in mice following a 90-d oral exposure. J. Toxicol. Environ. Health Part A 2018, 81, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Díez-Quijada, L.; Moyano, R.; Molina-Hernández, V.; Cameán, A.M.; Jos, A. Evaluation of toxic effects induced by repeated exposure to Cylindrospermopsin in rats using a 28-day feeding study. Food Chem. Toxicol. 2021, 151, 112108. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environ. Toxicol. Pharm. 2012, 34, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M.; Karczewski, J. Toxic potencies of metabolite(s) of non-cylindrospermopsin producing Cylindrospermopsis raciborskii isolated from temperate zone in human white cells. Chemosphere 2015, 120, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Puerto, M.; Pichardo, S.; Jos, Á.; Gutiérrez-Praena, D.; Cameán, A.M. Acute effects of pure Cylindrospermopsin on the activity and transcription of antioxidant enzymes in Tilapia (Oreochromis niloticus) exposed by gavage. Ecotoxicology 2011, 20, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Žegura, B.; Gajski, G.; Štraser, A.; Garaj-Vrhovac, V. Cylindrospermopsin induced DNA damage and alteration in the expression of genes involved in the response to DNA damage, apoptosis and oxidative stress. Toxicon 2011, 58, 471–479. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Cameán, A.M.; Jos, A. Neurotoxic assessment of microcystin-LR, cylindrospermopsin and their combination on the human neuroblastoma SH-SY5Y cell line. Chemosphere 2019, 224, 751–764. [Google Scholar] [CrossRef]
- Gusel’nikova, V.V.; Korzhevskiy, D.E. NeuN as a neuronal nuclear antigen and neuron differentiation marker. Acta Nat. 2015, 7, 42–47. [Google Scholar] [CrossRef]
- Johnson, G.V.W.; Jope, R.S. The role of microtubule-associated protein 2 (MAP-2) in neuronal growth, plasticity, and degeneration. J. Neurosci. Res. 1992, 33, 505–512. [Google Scholar] [CrossRef]
- Soltani, M.H.; Pichardo, R.; Song, Z.; Sangha, N.; Camacho, F.; Satyamoorthy, K.; Sangueza, O.P.; Setaluri, V. Microtubule-associated protein 2, a marker of neuronal differentiation, induces mitotic defects, inhibits growth of melanoma cells, and predicts metastatic potential of cutaneous melanoma. Am. J. Pathol. 2005, 166, 1841–1850. [Google Scholar] [CrossRef] [Green Version]
- Phelan, P.; Gordon-Weeks, P.R. Widespread Distribution of Synaptophysin, a Synaptic Vesicle Glycoprotein, in Growing Neurites and Growth Cones. Eur. J. Neurosci. 1992, 4, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Béïque, J.C.; Andrade, R. PSD-95 regulates synaptic transmission and plasticity in rat cerebral cortex. J. Physiol. 2003, 546, 859–867. [Google Scholar] [CrossRef]
- Ayeni, E.A.; Gong, Y.; Yuan, H.; Hu, Y.; Bai, X.; Liao, X. Medicinal plants for anti-neurodegenerative diseases in West Africa. J. Ethnopharmacol. 2021, 11, 114468. [Google Scholar] [CrossRef] [PubMed]
- Bondy, S.C. Anthropogenic pollutants may increase the incidence of neurodegenerative disease in an aging population. Toxicology 2016, 3, 41–46. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Cameán, A.M.; Jos, A. In vitro assessment of the combination of cylindrospermopsin and the organophosphate chlorpyrifos on the human neuroblastoma SH-SY5Y cell line. Ecotoxcol. Environm Saf. 2020, 191, 110222. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, R.; Constantinescu, A.T.; Reichmann, H.; Janetzky, B. Neuronal differentiation and long-term culture of the human neuroblastoma line SH-SY5Y. J. Neural Transm. Suppl. 2007, 72, 17–28. [Google Scholar]
- Heravi, M.; Dargahi, L.; Parsafar, S.; Tayaranian Marvian, A.; Aliakbari, F.; Morshedi, D. The primary neuronal cells are more resistant than PC12 cells to α-synuclein toxic aggregates. Neurosci. Lett. 2019, 14, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Giordano, G.; Costa, L.G. Primary neurons in culture and neuronal cell lines for in vitro neurotoxicological studies. Methods Mol. Biol. 2011, 758, 13–27. [Google Scholar] [PubMed]
- Yousefsani, B.S.; Akbarizadeh, N.; Pourahmad, J. The antioxidant and neuroprotective effects of Zolpidem on acrylamide-induced neurotoxicity using Wistar rat primary neuronal cortical culture. Toxicol. Rep. 2020, 7, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Q.; De Felice, F.G.; Fernandez, S.; Chen, H.; Lambert, M.P.; Quon, M.J.; Krafft, G.A.; Klein, W.L. Amyloid beta oligomers induce impairment of neuronal insulin receptors. FASEB J. 2008, 22, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Cossette, M.; Lévesque, D.; Parent, A. Neurochemical characterization of dopaminergic neurons in human striatum. Parkinsonism Relat. Disord. 2005, 11, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Varo, R.; Trujillo-Estrada, L.; Sánchez-Mejias, E.; Torres, M.; Baglietto-Vargas, D.; Moreno-González, I.; De Castro, V.; Jiménez, S.; Ruano, D.; Vizuete, M.; et al. Abnormal accumulation of autophagic vesicles correlates with axonal and synaptic pathology in young Alzheimer’s mice hippocampus. Acta Neuropathol. 2012, 123, 53–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, S.W. Effects of Continuous Exposure of SH-SY5Y Neuroblastoma Cells to Chlorpyrifos on Neuronal Differentiation: Implications for Neurotoxicity. Ph.D. Dissertation, Faculty of the Graduate School of the University of Maryland, Baltimore, MD, USA, 2017. [Google Scholar]
- Takser, L.; Benachour, N.; Husk, B.; Cabana, H.; Gris, D. Cyanotoxins at low doses induce apoptosis and inflammatory effects in murine brain cells: Potential implications for neurodegenerative diseases. Toxicol. Rep. 2016, 3, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glantz, L.A.; Gilmore, J.H.; Hamer, R.M.; Lieberman, J.A.; Jarskog, L.F. Synaptophysin and postsynaptic density protein 95 in the human prefrontal cortex from mid-gestation into early adulthood. Neuroscience 2007, 149, 582–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Kou, J.; Li, F.; Huo, D.; Xu, J.; Zhou, X.; Meng, D.; Ghulam, M.; Artyom, B.; Gao, X.; et al. Lemon essential oil ameliorates age-associated cognitive dysfunction via modulating hippocampal synaptic density and inhibiting acetylcholinesterase. Aging 2020, 12, 8622–8639. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar]
- Mata, G.; Cuesto, G.; Heras, J.; Morales, M.; Romero, A.; Rubio, J. SynapCountJ: A Validated Tool for Analyzing Synaptic Densities in Neurons. In Biomedical Engineering Systems and Technologies; BIOSTEC 2016; Communications in Computer and Information Science; Fred, A., Gamboa, H., Eds.; Springer: Cham, Switzerland, 2017; Volume 690, pp. 41–55. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinojosa, M.G.; Prieto, A.I.; Muñoz-Castro, C.; Sánchez-Mico, M.V.; Vitorica, J.; Cameán, A.M.; Jos, Á. Cytotoxicity and Effects on the Synapsis Induced by Pure Cylindrospermopsin in an E17 Embryonic Murine Primary Neuronal Culture in a Concentration- and Time-Dependent Manner. Toxins 2022, 14, 175. https://doi.org/10.3390/toxins14030175
Hinojosa MG, Prieto AI, Muñoz-Castro C, Sánchez-Mico MV, Vitorica J, Cameán AM, Jos Á. Cytotoxicity and Effects on the Synapsis Induced by Pure Cylindrospermopsin in an E17 Embryonic Murine Primary Neuronal Culture in a Concentration- and Time-Dependent Manner. Toxins. 2022; 14(3):175. https://doi.org/10.3390/toxins14030175
Chicago/Turabian StyleHinojosa, María G., Ana I. Prieto, Clara Muñoz-Castro, María V. Sánchez-Mico, Javier Vitorica, Ana M. Cameán, and Ángeles Jos. 2022. "Cytotoxicity and Effects on the Synapsis Induced by Pure Cylindrospermopsin in an E17 Embryonic Murine Primary Neuronal Culture in a Concentration- and Time-Dependent Manner" Toxins 14, no. 3: 175. https://doi.org/10.3390/toxins14030175
APA StyleHinojosa, M. G., Prieto, A. I., Muñoz-Castro, C., Sánchez-Mico, M. V., Vitorica, J., Cameán, A. M., & Jos, Á. (2022). Cytotoxicity and Effects on the Synapsis Induced by Pure Cylindrospermopsin in an E17 Embryonic Murine Primary Neuronal Culture in a Concentration- and Time-Dependent Manner. Toxins, 14(3), 175. https://doi.org/10.3390/toxins14030175