Low-Concentration T-2 Toxin Attenuates Pseudorabies Virus Replication in Porcine Kidney 15 Cells


Abstract
:1. Introduction
2. Results
2.1. T2 Cytotoxicity
2.2. T2 Inhibits the Cytotoxicity and Cell Death Caused by PRV Infection
2.2.1. The Number of Cells Infected by PRV
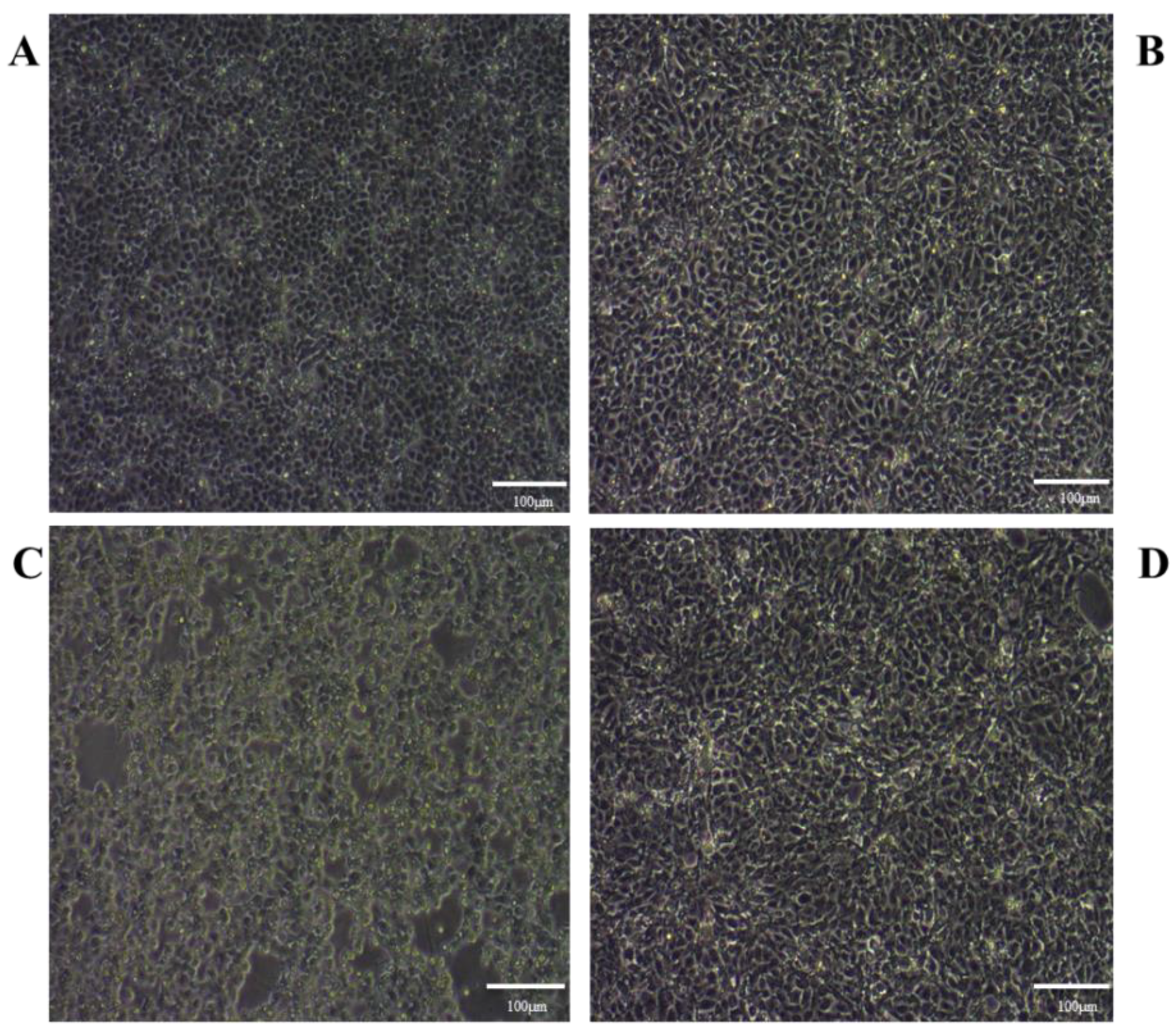
2.2.2. The Mode of Action Assay
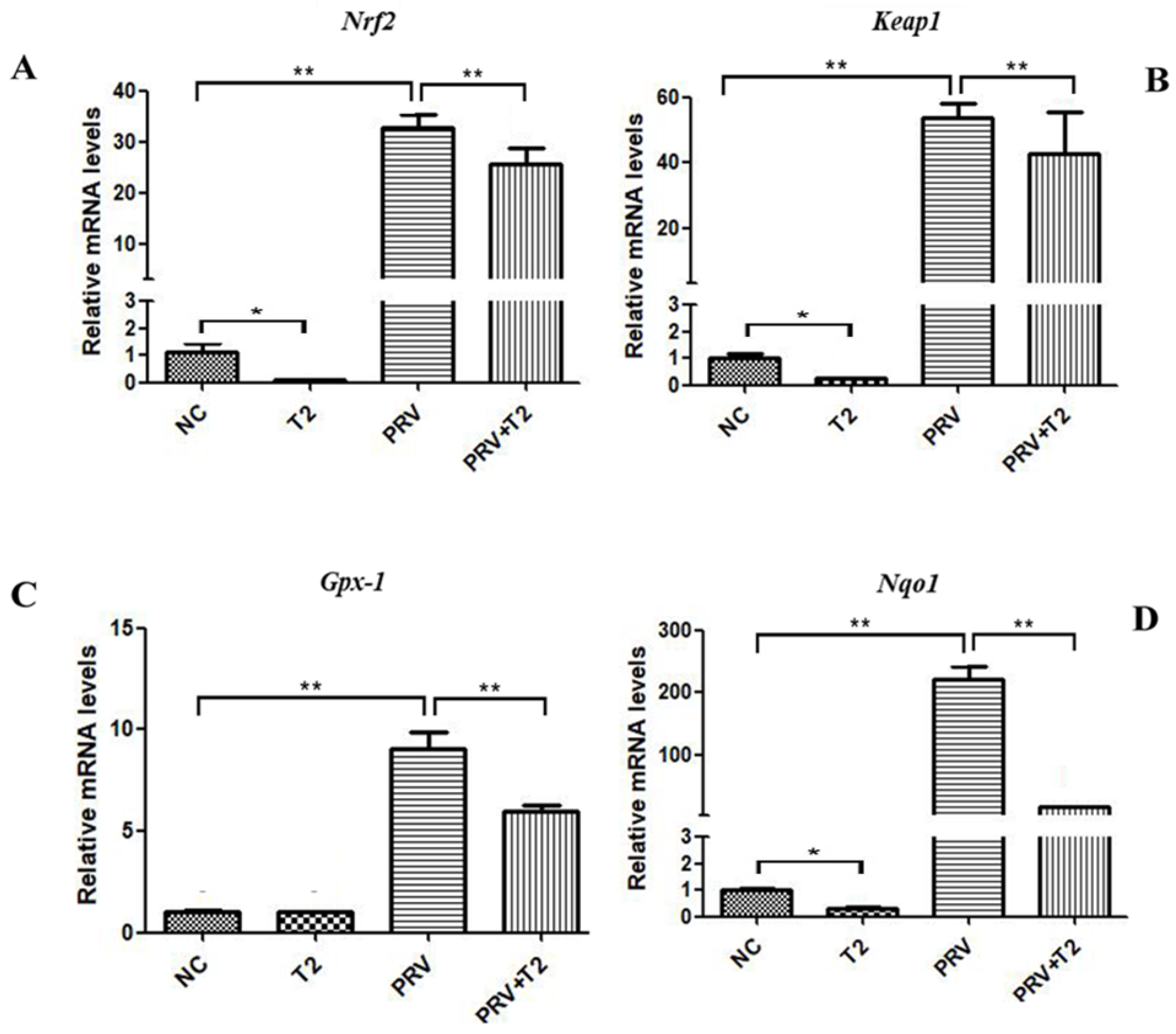
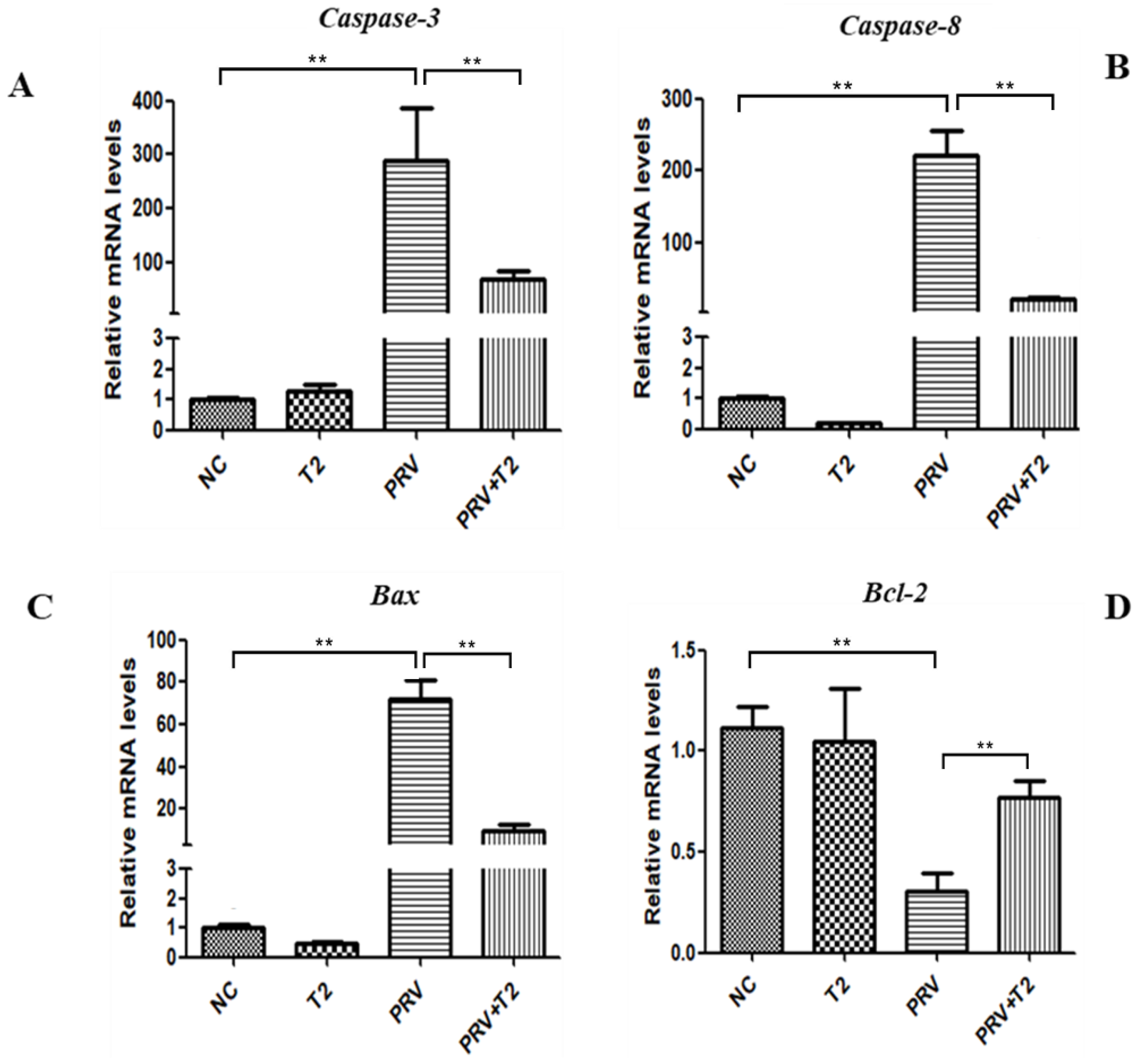
2.3. Oxidative Stress and Apoptosis Are Involved in the Inhibition of PRV Replication in PK15 Cells by Low-Concentration T2
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cells and Viruses
5.2. Reagents and Antibodies
5.3. Cell Cytotoxicity Assay
5.4. Morphological Investigation
5.5. Immunofluorescence Staining of Glycoprotein B (gB)
5.6. Determination of PRV DNA Copies by Real-Time PCR (qPCR)
5.7. The Mode of Action Assays
5.8. DNA and RNA Extraction and qPCR
5.9. Western Blot
5.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morcia, C.; Tumino, G.; Ghizzoni, R.; Badeck, F.; Lattanzio, V.; Pascale, M.; Terzi, V. Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley. Toxins 2016, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Gao, S.; Liu, Y.; Zhang, S.; Hu, R. Pseudorabies in farmed foxes fed pig offal in Shandong province, China. Arch. Virol. 2016, 161, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A novel human acute encephalitis caused by pseudorabies virus variant strain. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 73, e3690–e3700. [Google Scholar] [CrossRef]
- He, W.; Auclert, L.; Zhai, X.; Wong, G.; Zhang, C.; Zhu, H.; Xing, G.; Wang, S.; He, W.; Li, K.; et al. Interspecies Transmission, Genetic Diversity, and Evolutionary Dynamics of Pseudorabies Virus. J. Infect. Dis. 2019, 219, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Lai, I.; Chang, C.; Shih, W. Apoptosis Induction by Pseudorabies Virus via Oxidative Stress and Subsequent DNA Damage Signaling. Intervirology 2019, 62, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ma, G.; Fu, L.; Jia, H.; Zhu, M.; Li, X.; Zhao, S. Pseudorabies viral replication is inhibited by a novel target of miR-21. Virology 2014, 456, 319–328. [Google Scholar] [CrossRef]
- Huang, D.; Cui, L.; Sajid, A.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. The epigenetic mechanisms in Fusarium mycotoxins induced toxicities. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 123, 595–601. [Google Scholar] [CrossRef]
- Park, J.; Chang, H.; Kim, D.; Chung, S.; Lee, C. Long-Term Occurrence of Deoxynivalenol in Feed and Feed Raw Materials with a Special Focus on South Korea. Toxins 2018, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.C.; Liu, X.D.; Liu, J.C.; Wang, G.; Wan, K.Y. Contamination Level of T-2 and HT-2 Toxin in Cereal Crops from Aba Area in Sichuan Province, China. Bull. Environ. Contam. Toxicol. 2012, 88, 396–400. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Zheng, H.; He, C.; Zhang, H. T-2 toxin, zearalenone and fumonisin B1 in feedstuffs from China. Food Addit. Contaminants. Part B Surveill. 2013, 6, 116–122. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, Q.; Zheng, Y.; Chen, X.; Kong, D.; Huang, W.; Liu, P.; Jiang, H.; Jiang, Y. A rapid and accurate method for screening T-2 toxin in food and feed using competitive AlphaLISA. FEMS Microbiol. Lett. 2021, 368, fnab029. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food. Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, M.; Jayaraj, R.; Bhaskar, A.; Lakshmana Rao, P. Oxidative stress induction by T-2 toxin causes DNA damage and triggers apoptosis via caspase pathway in human cervical cancer cells. Toxicology 2009, 262, 153–161. [Google Scholar] [CrossRef]
- Yin, H.; Han, S.; Chen, Y.; Wang, Y.; Li, D.; Zhu, Q. T-2 Toxin Induces Oxidative Stress, Apoptosis and Cytoprotective Autophagy in Chicken Hepatocytes. Toxins 2020, 12, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, R.; Su, J.; Huang, D.; Gan, Y.; Yang, C.; Huang, G. Human hepatitis B virus and hepatocellular carcinoma. II. Experimental induction of hepatocellular carcinoma in tree shrews exposed to hepatitis B virus and aflatoxin B1. J. Cancer Res. Clin. Oncol. 1996, 122, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Yohannes, T.; Sharma, A.; Singh, S.; Goswami, T. Immunopathological effects of experimental T-2 mycotoxocosis in broiler chicken co-infected with infectious bronchitis virus (IBV). Vet. Immunol. Immunopathol. 2012, 146, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Han, M.; Zheng, M.; Zhao, J.; Chen, J.; Wen, J. Effects of dietary vitamin E on immunological stress of layers and their offspring. J. Anim. Physiol. Anim. Nutr. 2011, 95, 343–350. [Google Scholar] [CrossRef]
- Li, M.; Harkema, J.; Islam, Z.; Cuff, C.; Pestka, J. T-2 toxin impairs murine immune response to respiratory reovirus and exacerbates viral bronchiolitis. Toxicol. Appl. Pharmacol. 2006, 217, 76–85. [Google Scholar] [CrossRef]
- Savard, C.; Pinilla, V.; Provost, C.; Segura, M.; Gagnon, C.; Chorfi, Y. In vitro effect of deoxynivalenol (DON) mycotoxin on porcine reproductive and respiratory syndrome virus replication. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 65, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Savard, C.; Pinilla, V.; Provost, C.; Gagnon, C.; Chorfi, Y. In vivo effect of deoxynivalenol (DON) naturally contaminated feed on porcine reproductive and respiratory syndrome virus (PRRSV) infection. Vet. Microbiol. 2014, 174, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Savard, C.; Gagnon, C.; Chorfi, Y. Deoxynivalenol (DON) naturally contaminated feed impairs the immune response induced by porcine reproductive and respiratory syndrome virus (PRRSV) live attenuated vaccine. Vaccine 2015, 33, 3881–3886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, F.; Zhang, Z.; Hu, Z.; Hesketh, J.; Xue, H.; Chen, X.; Hao, S.; Huang, Y.; Cole Ezea, P.; Parveen, F.; et al. Ochratoxin A promotes porcine circovirus type 2 replication in vitro and in vivo. Free. Radic. Biol. Med. 2015, 80, 33–47. [Google Scholar] [CrossRef]
- Gan, F.; Hu, Z.; Huang, Y.; Xue, H.; Huang, D.; Qian, G.; Hu, J.; Chen, X.; Wang, T.; Huang, K. Overexpression of pig selenoprotein S blocks OTA-induced promotion of PCV2 replication by inhibiting oxidative stress and p38 phosphorylation in PK15 cells. Oncotarget 2016, 7, 20469–20485. [Google Scholar] [CrossRef]
- Qian, G.; Liu, D.; Hu, J.; Gan, F.; Hou, L.; Zhai, N.; Chen, X.; Huang, K. SeMet attenuates OTA-induced PCV2 replication promotion by inhibiting autophagy by activating the AKT/mTOR signaling pathway. Vet. Res. 2018, 49, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, B.; Singh, B.; Brar, R. Haematological and biochemical studies in broiler chicks fed ochratoxin and inoculated with inclusion body hepatitis virus, singly and in concurrence. Vet. Res. Commun. 1998, 22, 335–346. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, X.; Yang, W.; Nüssler, A.; Xiong, L.; Kuča, K.; Dohnal, V.; Zhang, X.; Yuan, Z. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef] [PubMed]
- Rafai, P.; Papp, Z.; Jakab, L. Biotransformation of trichothecenes alleviates the negative effects of T-2 toxin in pigs. Acta Vet. Hung. 2013, 61, 333–343. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Liu, S.; Wang, J.; Liu, X.; Zhu, Y.; Zhang, L.; Li, R. Betulinic acid attenuates T-2 toxin-induced cytotoxicity in porcine kidney cells by blocking oxidative stress and endoplasmic reticulum stress. Comp. Biochem. Physiology. Toxicol. Pharmacol. CBP 2021, 249, 109124. [Google Scholar] [CrossRef]
- Seeboth, J.; Solinhac, R.; Oswald, I.; Guzylack-Piriou, L. The fungal T-2 toxin alters the activation of primary macrophages induced by TLR-agonists resulting in a decrease of the inflammatory response in the pig. Vet. Res. 2012, 43, 35. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, L.Q.; Zheng, H.H.; Yang, Y.R.; Chen, H.Y. Construction and immunogenicity of a gE/gI/TK-deleted PRV based on porcine pseudorabies virus variant. Mol. Cell. Probes 2020, 53, 101605. [Google Scholar] [CrossRef]
- Rychlik, A.; Gonkowski, S.; Kaczmar, E.; Obremski, K.; Calka, J.; Makowska, K. The T2 Toxin Produced by spp. Impacts Porcine Duodenal Nitric Oxide Synthase (nNOS)-Positive Nervous Structures-The Preliminary Study. Int. J. Mol. Sci. 2020, 21, 5118. [Google Scholar] [CrossRef] [PubMed]
- Taroncher, M.; Pigni, M.; Diana, M.; Juan-García, A.; Ruiz, M. Does low concentration mycotoxin exposure induce toxicity in HepG2 cells through oxidative stress? Toxicol. Mech. Methods 2020, 30, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cui, Q.; Fu, Q.; Song, X.; Jia, R.; Yang, Y.; Zou, Y.; Li, L.; He, C.; Liang, X.; et al. Antiviral properties of resveratrol against pseudorabies virus are associated with the inhibition of IκB kinase activation. Sci. Rep. 2017, 7, 8782. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) |
|---|---|
| PRV-gB | F: AAGTTCAAGGCCCACATCTA |
| R: TGAAGCGGTTCGTGATGG | |
| Caspase-3 | F: GGA ATG GCA TGT CGA TCT GGT |
| R: ACT GTC CGT CTC AAT CCC AC | |
| Caspase-8 | F: TCT GCG GAC TGG ATG TGA TT |
| R: TCT GAG GTT GCT GGT CAC AC | |
| Bax | F: ATG ATC GCA GCC GTG GAC ACG |
| R: ACG AAG ATG GTC ACC GTC GC | |
| Bcl-2 | F: GAA ACC CCT AGT GCC ATC AA |
| R: GGG ACG TCA GGT CAC TGA AT | |
| Gpx-1 | F: CGTGCAACCAGTTTGGACAT |
| R: AGCATGAAGTTGGGCTCGAA | |
| Nqo1 | F: GATCATACTGGCCCACTCCG |
| R: GAGCAGTCTCGGCAGGATAC | |
| Nrf2 | F: CATAGCAGAGCCCAGTACCA |
| R: AGGGGTTGGATTGGGTTTAGT | |
| Keap1 | F: ACTTTCGTAGCCCCCATGAA |
| R: ATCCCTAGCGTGCAGGTGT | |
| β-actin | F: CTG CGG CAT CCA CGA AAC T |
| R: AGG GCC GTG ATC TCC TTC TG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, K.; Tan, L.; Yi, S.; Wu, Y.; Hu, Y.; Wang, A.; Yang, L. Low-Concentration T-2 Toxin Attenuates Pseudorabies Virus Replication in Porcine Kidney 15 Cells. Toxins 2022, 14, 121. https://doi.org/10.3390/toxins14020121
Xiong K, Tan L, Yi S, Wu Y, Hu Y, Wang A, Yang L. Low-Concentration T-2 Toxin Attenuates Pseudorabies Virus Replication in Porcine Kidney 15 Cells. Toxins. 2022; 14(2):121. https://doi.org/10.3390/toxins14020121
Chicago/Turabian StyleXiong, Kuankuan, Lei Tan, Siliang Yi, Yingxin Wu, Yi Hu, Aibing Wang, and Lingchen Yang. 2022. "Low-Concentration T-2 Toxin Attenuates Pseudorabies Virus Replication in Porcine Kidney 15 Cells" Toxins 14, no. 2: 121. https://doi.org/10.3390/toxins14020121
APA StyleXiong, K., Tan, L., Yi, S., Wu, Y., Hu, Y., Wang, A., & Yang, L. (2022). Low-Concentration T-2 Toxin Attenuates Pseudorabies Virus Replication in Porcine Kidney 15 Cells. Toxins, 14(2), 121. https://doi.org/10.3390/toxins14020121