Peptides Isolated from Amphibian Skin Secretions with Emphasis on Antimicrobial Peptides


Abstract
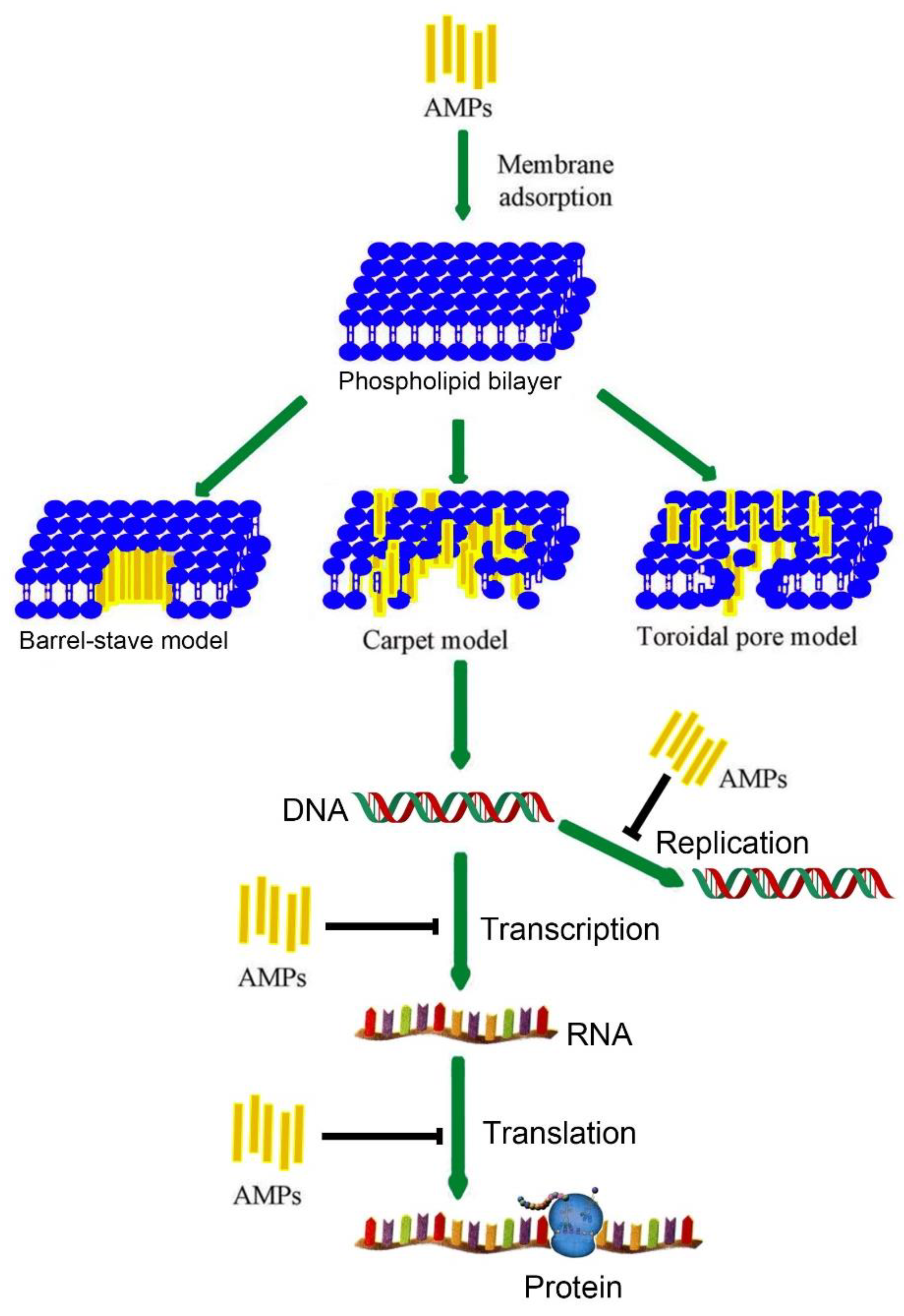
1. Introduction
2. Antimicrobial Peptides

2.1. Esculentins

{kind=link}
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Esculentin-1 | Rana esculenta | GIFSKLGRKKIKNLLISGLKNVGKEVGMDVVRTGIDIAGCKIKGEC | Hh: 19.57% Ee: 39.13% Cc: 41.30% | 46 | 0.2 (E. coli D21) 0.1 (B. megaterium BmII) 0.4 (S. aureus Cowan 1) 0.7 (P. aeruginosab ATCC15692) 0.5 (C. Albican) 0.9 (S. Cerevisiae) | [43] |
| Esculentin-1SEa | Rana sevosa | GLFSKFNKKKIKSGLIKIIKTAGKEAGLEALRTGIDVIGCKIKGEC | Hh: 36.96% Ee: 26.09% Cc: 36.96% | 46 | 1.1 (E. coli) 1.21 (M. luteus) | [44] |
| Esculentin-2a | Rana ridibunda | GILSLVKGVAKLAGKGLAKEGGKFGLELIACKIAKQC | Hh: 48.65% Ee: 8.11% Cc: 43.24% | 37 | 0.4 (E. coli D21) 0.2 (B. megaterium BmII) 0.8 (S. aureus Cowan 1) 3.5 (P. aeruginosab ATCC15692) | [44] |
| Esculentin-2SE | Rana sevosa | GFFSLIKGVAKIATKGLAKNLGKMGLDLVGCKISKEC | Hh: 43.24% Ee: 32.43% Cc: 24.32% | 37 | 0.3 (E. coli) 0.36 (M. luteus) | [44] |
| Esculentin-2-ALa | Amolops loloensis | GIFALIKTAAKFVGKNLLKQAGKAGLEHLACKANNQC | Hh: 62.16% Ee: 13.51% Cc: 24.32% | 37 | 12.5 (E. coli) 7.5 (B. dysenteriae) 25 (A. calcoaceticus) 50 (P. aeruginosa) 2.5 (S. aureus) 2.5 (B. pumilus) 7.5 (B. cereus) | [45] |
| Esculentin-2-ALb | GIFSLIKTAAKFVGKNLLKQAGKAGVEHLACKANNQC | Hh: 56.76% Ee: 5.41% Cc: 37.84% | 37 | 12.5 (E. coli) 7.5 (B. dysenteriae) 12.5 (A. calcoaceticus) 50 (P. aeruginosa) 1.25 (S. aureus) 2.5 (B. pumilus) 7.5 (B. cereus) | ||
| Esculentin-2CHa | Lithobates. chiricahuensis | GFSSIFRGVAKFASKGLGK DLAKLGVDLVA CKISKQC | Hh: 54.05% Cc: 45.95% | 37 | <6 (S. aureus) | [46] |
| Esculentin 2EM | Glandirana emeljanovi | GILDTLKQFAKGVGKDLVKGAAQGVLSTVSCKLAKTC | Hh: 27.03% Ee: 32.43% Cc: 40.54% | 37 | >75 (E. coli) <6.25 (S. aureus) | [47] |
2.2. Brevinins
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Brevinin-1 | Rana brevipoda porsa | FLPVLAGIAAKVVPALFCKITKKC | Hh: 33.33% Ee: 16.67% Cc: 50.00% | 24 | 34 (E. coli) 8 (S. aureus) | [48] |
| Brevinin-1GHd | Hylarana guentheri | FLGALFKVASKLVPAAICSISKKC | Hh: 16.67% Ee: 37.50% Cc: 45.83% | 24 | 8 (E. coli) 32 (P. aeruginosa) 2 (S. aureus) 4 (MRSA) 4 (C. albicans) | [58] |
| Brevinin-1AUa | Rana aurora aurora | FLPILAGLAAKLVPKVFCSITKKC | Hh: 12.50% Ee: 33.33% Cc: 54.17% | 24 | 13 (E. coli) 25 (P. aeruginosa) 13 (E. cloacae) 50 (K. pneumoniae) 3 (S. aureus) 6 (S. epidermidis) 3 (C. albicans) | [59] |
| Brevinin-1AUb | FLPILAGLAANILPKVFCSITKKC | Ee: 33.33% Cc: 66.67% | 24 | 25 (E. coli) 25 (P. aeruginosa) 25 (E. cloacae) >50 (K. pneumoniae) 3 (S. aureus) 6 (S. epidermidis) 3 (C. albicans) | ||
| Brevinin-1BYa | Rana boylii | FLPILASLAAKFGPKLFCLVTKKC | Hh: 16.67% Ee: 33.33% Cc: 50.00% | 24 | 17 (E. coli) 2 (S. aureus) 3 (C. albicans) | [50] |
| Brevinin-1BYb | LPILASLAAKLGPKLFCLVTKKC | Hh: 13.04% Ee: 30.43% Cc: 56.52% | 23 | 16 (E. coli) 4 (S. aureus) 16 (C. albicans) | ||
| Brevinin-1BYc | LPILASLAATLGPKLLCLITKKC | Ee: 39.13% Cc: 60.87% | 23 | NA (E. coli) 8 (S. aureus) 35 (C. albicans) | ||
| Brevinin-1SY | Rana sylvatica | FLPVVAGLAAKVLPSIICAVTKKC | Ee: 41.67% Cc: 58.33% | 24 | 45 (E. coli) 7 (S. aureus) | [51] |
| Brevinin-1LTa | Hylarana latouchii | FFGTALKIAANVLPTAICKILKKC | Hh: 25.00% Ee: 29.17% Cc: 45.83% | 24 | 80 (P. fluorescens) 10 (S. aureus) 6 (B. subtilis) 50 (C. albicans) | [60] |
| Brevinin-1Sa | Rana sphenocephala | FLPAIVGAAGQFLPKIFCAISKKC | Ee: 41.67% Cc: 58.33% | 24 | 55 (E. coli) | [61] |
| Brevinin-1Sb | FLPAIVGAAGKFLPKIFCAISKKC | Ee: 54.17% Cc: 45.83% | 24 | 17 (E. coli) | ||
| Brevinin-1Sc | FFPIVAGVAGQVLKKIYCTISKKC | Ee: 62.50% Cc: 37.50% | 24 | 14 (E. coli) | ||
| Brevinin-2 | Rana brevipoda porsa | GLLDSLKGFAATAGKGVLQSLLSTASCKLAKTC | Hh: 39.39% Ee: 6.06% Cc: 54.55% | 33 | 4 (E. coli) 8 (S. aureus) | [48] |
| Brevinin-2E | Rana esculenta | GIMDTLKNLAKTAGKGALQSLLNKASCKLSGQC | Hh: 48.48% Cc: 51.52% | 33 | 0.5 (E. coli D21) 0.2 (B. megaterium BmII) 2 (S. aureus Cowan 1) >30 (P. aeruginosab ATCC15692) NA (C. albican) NA (S. cerevisiae) | [43] |
| Brevinin-2Oa | Rana ornativentris | GLFNVFKGALKTAGKHVAGSLLNQLKCKVSGGC | Hh: 42.42% Ee: 27.27% Cc: 30.30% | 33 | 2 (E. coli) 10 (S. aureus) 40 (C. albicans) | [62] |
| Brevinin-2Ob | GIFNVFKGALKTAGKHVAGSLLNQLKCKVSGEC | Hh: 45.45% Ee: 33.33% Cc: 21.21% | 33 | 4 (E. coli) 9 (S. aureus) 40 (C. albicans) |
2.3. Ranatuerins
2.4. Ranacyclins
2.5. Temporins
2.6. Bombinins
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| BHL-bombinin | Bombina orientalis | GIGGALLSFGKSALKGLAKGLAEHF | Hh: 44.00% Ee: 24.00% Cc: 32.00% | 25 | 6.6 (E. coli) 26.2 (P. aeruginosa) 1.6 (S. aureus) 6.6 (MRSA) | [96] |
| Bombinin HL | LLGPVLGLVSNVLGGLL | Ee: 58.82% Cc: 41.18% | 17 | NA (E. coli) NA (P. aeruginosa) 156.8 (S. aureus) NA (MRSA) | ||
| Bombinin-BO1 | Bombina orientalis | GIGSAILSAGKSIIKGLAKGLAEHF | Hh: 28.00% Ee: 20.00% Cc: 52.00% | 25 | 26.3 (E. coli) 26.3 (S. aureus) 52.5 (C. albicans) | [97] |
| Bombinin H-BO1 | IIGPVLGLVGKALGGLL | Ee: 47.06% Cc: 52.94% | 17 | 26.3 (E. coli) 26.3 (S. aureus) 52.5 (C. albicans) | ||
| Bombinin H1 | Bombina variegata | IIGPVLGMVGSALGGLLKKIG | Hh: 23.81% Ee: 38.10% Cc: 38.10% | 21 | 3.8 (E. coli D 21) 2.1 (S. aureus Cowan 1) | [98] |
| Bombinin H3 | IIGPVLGMVGSALGGLLKKIG | Hh: 23.81% Ee: 38.10% Cc: 38.10% | 21 | 3.7 (E. coli D 21) 2.4 (S. aureus Cowan 1) | ||
| Bombinin H4 | LIGPVLGLVGSALGGLLKKIG | Hh: 23.81% Ee: 42.86% Cc: 33.33% | 21 | 4.8 (E. coli D 21) 3.3 (S. aureus Cowan 1) |
2.7. Dybowskins
3. Antioxidant Peptides
4. Bradykinin Peptides
5. Insulin-Releasing Peptides
6. Other Peptides
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Przybylski, R.; Firdaous, L.; Châtaigné, G.; Dhulster, P.; Nedjar, N. Production of an antimicrobial peptide derived from slaughterhouse by-product and its potential application on meat as preservative. Food Chem. 2016, 211, 306–313. [Google Scholar] [CrossRef]
- Agrawal, H.; Joshi, R.; Gupta, M. Isolation, purification and characterization of antioxidative peptide of pearl millet (Pennisetum glaucum) protein hydrolysate. Food Chem. 2016, 204, 365–372. [Google Scholar] [CrossRef]
- Shanmugam, V.P.; Kapila, S.; Sonfack, T.K.; Kapila, R. Antioxidative peptide derived from enzymatic digestion of buffalo casein. Int. Dairy J. 2015, 42, 1–5. [Google Scholar] [CrossRef]
- Tu, M.L.; Feng, L.T.; Wang, Z.Y.; Qiao, M.L.; Shahidi, F.; Lu, W.H.; Du, M. Sequence analysis and molecular docking of antithrombotic peptides from casein hydrolysate by trypsin digestion. J. Funct. Foods 2017, 32, 313–323. [Google Scholar] [CrossRef]
- Zhang, S.B. In vitro antithrombotic activities of peanut protein hydrolysates. Food Chem. 2016, 202, 1–8. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Xue, J.; Kwok, L.Y.; Yang, J.; Zhang, H.; Menghe, B. Characterization of angiotensin-converting enzyme inhibitory activity of fermented milk produced by Lactobacillus helveticus. J. Dairy Sci. 2015, 98, 5113–5124. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Xue, J.; Yang, J.; Zhang, Q.; Zhang, H.; Chen, Y. Fermentation characteristics and angiotensin I-converting enzyme-inhibitory activity of Lactobacillus helveticus isolate H9 in cow milk, soy milk, and mare milk. J. Dairy Sci. 2015, 98, 3655–3664. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Xu, B. Anti-inflammatory effects of phytochemicals from fruits, vegetables, and food legumes: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1260–1270. [Google Scholar] [CrossRef]
- Gong, X.; An, Q.; Le, L.; Geng, F.; Jiang, L.; Yan, J.; Xiang, D.; Peng, L.; Zou, L.; Zhao, G.; et al. Prospects of cereal protein-derived bioactive peptides: Sources, bioactivities diversity, and production. Crit. Rev. Food Sci. Nutr. 2022, 62, 2855–2871. [Google Scholar] [CrossRef]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial peptides (AMPs) from fish epidermis: Perspectives for investigative dermatology. J. Invest. Dermatol. 2013, 133, 1140–1149. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Zhao, Y.; Wang, H.; Zhang, H. Effect of heat treatment on the enzymatic stability of grass carp skin collagen and its ability to form fibrils in vitro. J. Sci. Food Agric. 2015, 95, 329–336. [Google Scholar] [CrossRef]
- Cao, H.; Zhao, Y.; Zhu, Y.B.; Xu, F.; Yu, J.S.; Yuan, M. Antifreeze and cryoprotective activities of ice-binding collagen peptides from pig skin. Food Chem. 2016, 194, 1245–1253. [Google Scholar] [CrossRef]
- Hong, H.; Fan, H.; Roy, B.C.; Wu, J. Amylase enhances production of low molecular weight collagen peptides from the skin of spent hen, bovine, porcine, and tilapia. Food Chem. 2021, 352, 129355. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, J.; Yang, X.; Huang, F. Hypoglycemic activity of CPU2206: A novel peptide from sika (Cervus nippon Temminck) antler. J. Food Biochem. 2019, 43, e13063. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, X.; Zhang, X.L.; Xie, Q.F. Purification and identification of anti-inflammatory peptides derived from simulated gastrointestinal digests of velvet antler protein (Cervus elaphus Linnaeus). J. Food Drug. Anal. 2016, 24, 376–384. [Google Scholar] [CrossRef]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Demori, I.; Rashed, Z.E.; Corradino, V.; Catalano, A.; Rovegno, L.; Queirolo, L.; Salvidio, S.; Biggi, E.; Zanotti-Russo, M.; Canesi, L.; et al. Peptides for Skin Protection and Healing in Amphibians. Molecules 2019, 24, 347. [Google Scholar] [CrossRef]
- He, X.; Yang, S.; Wei, L.; Liu, R.; Lai, R.; Rong, M. Antimicrobial peptide diversity in the skin of the torrent frog, Amolops jingdongensis. Amino Acids 2013, 44, 481–487. [Google Scholar] [CrossRef]
- Wang, H.; Yu, Z.; Hu, Y.; Li, F.; Liu, L.; Zheng, H.; Meng, H.; Yang, S.; Yang, X.; Liu, J. Novel antimicrobial peptides isolated from the skin secretions of Hainan odorous frog, Odorrana hainanensis. Peptides 2012, 35, 285–290. [Google Scholar] [CrossRef]
- Feng, G.; Wu, J.; Yang, H.L.; Mu, L. Discovery of Antioxidant Peptides from Amphibians: A Review. Protein Pept. Lett. 2021, 28, 1220–1229. [Google Scholar] [CrossRef]
- Guo, C.; Hu, Y.; Li, J.; Liu, Y.; Li, S.; Yan, K.; Wang, X.; Liu, J.; Wang, H. Identification of multiple peptides with antioxidant and antimicrobial activities from skin and its secretions of Hylarana taipehensis, Amolops lifanensis, and Amolops granulosus. Biochimie 2014, 105, 192–201. [Google Scholar] [CrossRef]
- Bergaoui, I.; Zairi, A.; Tangy, F.; Aouni, M.; Selmi, B.; Hani, K. In vitro antiviral activity of dermaseptin S(4) and derivatives from amphibian skin against herpes simplex virus type 2. J. Med. Virol. 2013, 85, 272–281. [Google Scholar] [CrossRef]
- Chianese, A.; Zannella, C.; Monti, A.; De Filippis, A.; Doti, N.; Franci, G.; Galdiero, M. The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23. Int. J. Mol. Sci. 2022, 23, 883. [Google Scholar] [CrossRef]
- van Zoggel, H.; Hamma-Kourbali, Y.; Galanth, C.; Ladram, A.; Nicolas, P.; Courty, J.; Amiche, M.; Delbe, J. Antitumor and angiostatic peptides from frog skin secretions. Amino Acids 2012, 42, 385–395. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, X.; Fu, C.; Zhao, R.; Jin, T.; Liu, M.; Pan, C.; Li, L.A.; Ma, J.; Yu, E.; et al. Antineoplastic Effects and Mechanisms of a New RGD Chimeric Peptide from Bullfrog Skin on the Proliferation and Apoptosis of B16F10 Cells. Protein J. 2021, 40, 709–720. [Google Scholar] [CrossRef]
- Bevins, C.L.; Zasloff, M. Peptides from frog skin. Annu. Rev. Biochem. 1990, 59, 395–414. [Google Scholar] [CrossRef]
- You, D.; Hong, J.; Rong, M.; Yu, H.; Liang, S.; Ma, Y.; Yang, H.; Wu, J.; Lin, D.; Lai, R. The first gene-encoded amphibian neurotoxin. J. Biol. Chem. 2009, 284, 22079–22086. [Google Scholar] [CrossRef]
- Xiao, J.; Jiang, D. On origin of Oviductus Ranae in Chinese Pharmacopoeia. Zhongguo Zhong Yao Za Zhi 2010, 35, 2931–2933. [Google Scholar]
- Laux-Biehlmann, A.; Mouheiche, J.; Veriepe, J.; Goumon, Y. Endogenous morphine and its metabolites in mammals: History, synthesis, localization and perspectives. Neuroscience 2013, 233, 95–117. [Google Scholar] [CrossRef]
- Bastos, P.; Trindade, F.; da Costa, J.; Ferreira, R.; Vitorino, R. Human Antimicrobial Peptides in Bodily Fluids: Current Knowledge and Therapeutic Perspectives in the Postantibiotic Era. Med. Res. Rev. 2018, 38, 101–146. [Google Scholar] [CrossRef]
- Cao, X.; Zhang, Y.; Mao, R.; Teng, D.; Wang, X.; Wang, J. Design and recombination expression of a novel plectasin-derived peptide MP1106 and its properties against Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2015, 99, 2649–2662. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial Peptides: Interaction With Model and Biological Membranes and Synergism With Chemical Antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Cilli, E.M.; Fontes, W. Combinatorial synthesis and directed evolution applied to the production of alpha-helix forming antimicrobial peptides analogues. Curr Protein Pept. Sci. 2006, 7, 473–478. [Google Scholar] [CrossRef]
- Shabir, U.; Ali, S.; Magray, A.R.; Ganai, B.A.; Firdous, P.; Hassan, T.; Nazir, R. Fish antimicrobial peptides (AMP’s) as essential and promising molecular therapeutic agents: A review. Microb. Pathog. 2018, 114, 50–56. [Google Scholar] [CrossRef]
- Han, J.; Zhao, S.; Ma, Z.; Gao, L.; Liu, H.; Muhammad, U.; Lu, Z.; Lv, F.; Bie, X. The antibacterial activity and modes of LI-F type antimicrobial peptides against Bacillus cereus in vitro. J. Appl. Microbiol. 2017, 123, 602–614. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Haney, E.F.; Hunter, H.N.; Matsuzaki, K.; Vogel, H.J. Solution NMR studies of amphibian antimicrobial peptides: Linking structure to function? Biochim. Biophys. Acta 2009, 1788, 1639–1655. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef]
- Mihajlovic, M.; Lazaridis, T. Antimicrobial peptides bind more strongly to membrane pores. Biochim. Biophys. Acta 2010, 1798, 1494–1502. [Google Scholar] [CrossRef]
- Jean-François, F.; Elezgaray, J.; Berson, P.; Vacher, P.; Dufourc, E.J. Pore formation induced by an antimicrobial peptide: Electrostatic effects. Biophys. J. 2008, 95, 5748–5756. [Google Scholar] [CrossRef]
- Konig, E.; Bininda-Emonds, O.R.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from ranid frogs: Taxonomic and phylogenetic markers and a potential source of new therapeutic agents. Biochim. Biophys. Acta 2004, 1696, 1–14. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Barra, D.; Bossa, F. Antimicrobial peptides from skin secretions of Rana esculenta. Molecular cloning of cDNAs encoding esculentin and brevinins and isolation of new active peptides. J. Biol. Chem. 1994, 269, 11956–11961. [Google Scholar] [CrossRef]
- Graham, C.; Richter, S.C.; McClean, S.; O’Kane, E.; Flatt, P.R.; Shaw, C. Histamine-releasing and antimicrobial peptides from the skin secretions of the dusky gopher frog, Rana sevosa. Peptides 2006, 27, 1313–1319. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Wang, A.; Song, Y.; Ma, D.; Yang, H.; Ma, Y.; Lai, R. Five novel antimicrobial peptides from skin secretions of the frog, Amolops loloensis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 155, 72–76. [Google Scholar] [CrossRef]
- Attoub, S.; Mechkarska, M.; Sonnevend, A.; Radosavljevic, G.; Jovanovic, I.; Lukic, M.L.; Conlon, J.M. Esculentin-2CHa: A host-defense peptide with differential cytotoxicity against bacteria, erythrocytes and tumor cells. Peptides 2013, 39, 95–102. [Google Scholar] [CrossRef]
- Malik, E.; Phoenix, D.A.; Snape, T.J.; Harris, F.; Singh, J.; Morton, L.H.G.; Dennison, S.R. Linearized esculentin-2EM shows pH dependent antibacterial activity with an alkaline optimum. Mol. Cell. Biochem. 2021, 476, 3729–3744. [Google Scholar] [CrossRef]
- Morikawa, N.; Hagiwara, K.; Nakajima, T. Brevinin-1 and -2, unique antimicrobial peptides from the skin of the frog, Rana brevipoda porsa. Biochem. Biophys. Res. Commun. 1992, 189, 184–190. [Google Scholar] [CrossRef]
- Suh, J.Y.; Lee, K.H.; Chi, S.W.; Hong, S.Y.; Choi, B.W.; Moon, H.M.; Choi, B.S. Unusually stable helical kink in the antimicrobial peptide--a derivative of gaegurin. FEBS Lett. 1996, 392, 309–312. [Google Scholar] [CrossRef]
- Conlon, J.M.; Sonnevend, A.; Patel, M.; Davidson, C.; Nielsen, P.F.; Pal, T.; Rollins-Smith, L.A. Isolation of peptides of the brevinin-1 family with potent candidacidal activity from the skin secretions of the frog Rana boylii. J. Pept. Res. 2003, 62, 207–213. [Google Scholar] [CrossRef]
- Matutte, B.; Storey, K.B.; Knoop, F.C.; Conlon, J.M. Induction of synthesis of an antimicrobial peptide in the skin of the freeze-tolerant frog, Rana sylvatica, in response to environmental stimuli. FEBS Lett. 2000, 483, 135–138. [Google Scholar] [CrossRef]
- Goraya, J.; Wang, Y.; Li, Z.; O’Flaherty, M.; Knoop, F.C.; Platz, J.E.; Conlon, J.M. Peptides with antimicrobial activity from four different families isolated from the skins of the North American frogs Rana luteiventris, Rana berlandieri and Rana pipiens. Eur. J. Biochem. 2000, 267, 894–900. [Google Scholar] [CrossRef]
- Kwon, M.Y.; Hong, S.Y.; Lee, K.H. Structure-activity analysis of brevinin 1E amide, an antimicrobial peptide from Rana esculenta. Biochim. Biophys. Acta 1998, 1387, 239–248. [Google Scholar] [CrossRef]
- Kumari, V.K.; Nagaraj, R. Structure-function studies on the amphibian peptide brevinin 1E: Translocating the cationic segment from the C-terminal end to a central position favors selective antibacterial activity. J. Pept. Res. 2001, 58, 433–441. [Google Scholar] [CrossRef]
- Ali, M.F.; Knoop, F.C.; Vaudry, H.; Conlon, J.M. Characterization of novel antimicrobial peptides from the skins of frogs of the Rana esculenta complex. Peptides 2003, 24, 955–961. [Google Scholar] [CrossRef]
- Wang, Y.; Knoop, F.C.; Remy-Jouet, I.; Delarue, C.; Vaudry, H.; Conlon, J.M. Antimicrobial peptides of the brevinin-2 family isolated from gastric tissue of the frog, Rana esculenta. Biochem. Biophys. Res. Commun. 1998, 253, 600–603. [Google Scholar] [CrossRef]
- Park, J.M.; Jung, J.E.; Lee, B.J. Antimicrobial peptides from the skin of a Korean frog, Rana rugosa. Biochem. Biophys. Res. Commun. 1994, 205, 948–954. [Google Scholar] [CrossRef]
- Jiang, Y.; Wu, Y.; Wang, T.; Chen, X.; Zhou, M.; Ma, C.; Xi, X.; Zhang, Y.; Chen, T.; Shaw, C.; et al. Brevinin-1GHd: A novel Hylarana guentheri skin secretion-derived Brevinin-1 type peptide with antimicrobial and anticancer therapeutic potential. Biosci. Rep. 2020, 40, BSR20200019. [Google Scholar] [CrossRef]
- Conlon, J.M.; Sonnevend, A.; Davidson, C.; Demandt, A.; Jouenne, T. Host-defense peptides isolated from the skin secretions of the Northern red-legged frog Rana aurora aurora. Dev. Comp. Immunol. 2005, 29, 83–90. [Google Scholar] [CrossRef]
- Kang, S.J.; Son, W.S.; Han, K.D.; Mishig-Ochir, T.; Kim, D.W.; Kim, J.I.; Lee, B.J. Solution structure of antimicrobial peptide esculentin-1c from skin secretion of Rana esculenta. Mol. Cells 2010, 30, 435–441. [Google Scholar] [CrossRef]
- Conlon, J.M.; Halverson, T.; Dulka, J.; Platz, J.E.; Knoop, F.C. Peptides with antimicrobial activity of the brevinin-1 family isolated from skin secretions of the southern leopard frog, Rana sphenocephala. J. Pept. Res. 1999, 54, 522–527. [Google Scholar] [CrossRef]
- Kim, J.B.; Iwamuro, S.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides from the skin of the Japanese mountain brown frog, Rana ornativentris. J. Pept. Res. 2001, 58, 349–356. [Google Scholar] [CrossRef]
- Goraya, J.; Knoop, F.C.; Conlon, J.M. Ranatuerins: Antimicrobial peptides isolated from the skin of the American bullfrog, Rana catesbeiana. Biochem. Biophys. Res. Commun. 1998, 250, 589–592. [Google Scholar] [CrossRef]
- Sonnevend, A.; Knoop, F.C.; Patel, M.; Pal, T.; Soto, A.M.; Conlon, J.M. Antimicrobial properties of the frog skin peptide, ranatuerin-1 and its [Lys-8]-substituted analog. Peptides 2004, 25, 29–36. [Google Scholar] [CrossRef]
- Halverson, T.; Basir, Y.J.; Knoop, F.C.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin of the North American green frog Rana clamitans. Peptides 2000, 21, 469–476. [Google Scholar] [CrossRef]
- Goraya, J.; Knoop, F.C.; Conlon, J.M. Ranatuerin 1T: An antimicrobial peptide isolated from the skin of the frog, Rana temporaria. Peptides 1999, 20, 159–163. [Google Scholar] [CrossRef]
- Subasinghage, A.P.; Conlon, J.M.; Hewage, C.M. Conformational analysis of the broad-spectrum antibacterial peptide, ranatuerin-2CSa: Identification of a full length helix-turn-helix motif. Biochim. Biophys. Acta 2008, 1784, 924–929. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, D.; Zhong, R.; Ye, Z.; Ma, C.; Zhou, M.; Xi, X.; Wang, L.; Chen, T.; Kwok, H.F. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and its Truncated Analogues. Biomolecules 2019, 9, 249. [Google Scholar] [CrossRef]
- Yang, S.J.; Xiao, X.H.; Xu, Y.G.; Li, D.D.; Chai, L.H.; Zhang, J.Y. Induction of antimicrobial peptides from Rana dybowskii under Rana grylio virus stress, and bioactivity analysis. Can. J. Microbiol. 2012, 58, 848–855. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Mignogna, G.; Andreu, D.; Shai, Y.; Barra, D.; Simmaco, M. Ranacyclins, a new family of short cyclic antimicrobial peptides: Biological function, mode of action, and parameters involved in target specificity. Biochemistry 2003, 42, 14023–14035. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from the skins of North American frogs. Biochim. Biophys. Acta 2009, 1788, 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, Y.; Chen, X.; Wang, L.; Ma, C.; Xi, X.; Zhang, Y.; Chen, T.; Shaw, C.; Zhou, M. Ranacyclin-NF, a Novel Bowman-Birk Type Protease Inhibitor from the Skin Secretion of the East Asian Frog, Pelophylax nigromaculatus. Biology 2020, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; De Biase, D.; Severini, C.; Aita, M.; Erspamer, G.F.; Barra, D.; Bossa, F. Purification and characterization of bioactive peptides from skin extracts of Rana esculenta. Biochim. Biophys. Acta 1990, 1033, 318–323. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef]
- Basir, Y.J.; Knoop, F.C.; Dulka, J.; Conlon, J.M. Multiple antimicrobial peptides and peptides related to bradykinin and neuromedin N isolated from skin secretions of the pickerel frog, Rana palustris. Biochim. Biophys. Acta 2000, 1543, 95–105. [Google Scholar] [CrossRef]
- Kim, J.B.; Halverson, T.; Basir, Y.J.; Dulka, J.; Knoop, F.C.; Abel, P.W.; Conlon, J.M. Purification and characterization of antimicrobial and vasorelaxant peptides from skin extracts and skin secretions of the North American pig frog Rana grylio. Regul. Pept. 2000, 90, 53–60. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Ghaferi, N.; Abraham, B.; Jiansheng, H.; Cosette, P.; Leprince, J.; Jouenne, T.; Vaudry, H. Antimicrobial peptides from diverse families isolated from the skin of the Asian frog, Rana grahami. Peptides 2006, 27, 2111–2117. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, Y.; Chen, T.; Fang, X.; Walker, B.; Shaw, C. Components of the peptidome and transcriptome persist in lin wa pi: The dried skin of the Heilongjiang brown frog (Rana amurensis) as used in traditional Chinese medicine. Peptides 2006, 27, 2688–2694. [Google Scholar] [CrossRef]
- Bellavita, R.; Casciaro, B.; Di Maro, S.; Brancaccio, D.; Carotenuto, A.; Falanga, A.; Cappiello, F.; Buommino, E.; Galdiero, S.; Novellino, E.; et al. First-in-Class Cyclic Temporin L Analogue: Design, Synthesis, and Antimicrobial Assessment. J. Med. Chem. 2021, 64, 11675–11694. [Google Scholar] [CrossRef]
- Wade, D.; Silberring, J.; Soliymani, R.; Heikkinen, S.; Kilpelainen, I.; Lankinen, H.; Kuusela, P. Antibacterial activities of temporin A analogs. FEBS Lett. 2000, 479, 6–9. [Google Scholar] [CrossRef]
- Bhattacharjya, S.; Straus, S.K. Design, Engineering and Discovery of Novel alpha-Helical and beta-Boomerang Antimicrobial Peptides against Drug Resistant Bacteria. Int. J. Mol. Sci. 2020, 21, 5773. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.; Mohanram, H.; Domadia, P.N.; Torres, J.; Bhattacharjya, S. Designed beta-boomerang antiendotoxic and antimicrobial peptides: Structures and activities in lipopolysaccharide. J. Biol. Chem. 2009, 284, 21991–22004. [Google Scholar] [CrossRef] [PubMed]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.M.; Marcellini, L.; Luca, V.; Barra, D.; Novellino, E.; et al. The effect of d-amino acid substitution on the selectivity of temporin L towards target cells: Identification of a potent anti-Candida peptide. Biochim. Biophys. Acta 2013, 1828, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Diao, Y.; Han, W.; Zhao, H.; Zhu, S.; Liu, X.; Feng, X.; Gu, J.; Yao, C.; Liu, S.; Sun, C.; et al. Designed synthetic analogs of the α-helical peptide temporin-La with improved antitumor efficacies via charge modification and incorporation of the integrin αvβ3 homing domain. J. Pept. Sci. 2012, 18, 476–486. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin, L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368 Pt 1, 91–100. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta 2009, 1788, 1610–1619. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Rinaldi, A.C.; Di Giulio, A.; Mignogna, G.; Bozzi, A.; Barra, D.; Simmaco, M. Structure-function relationships of temporins, small antimicrobial peptides from amphibian skin. Eur. J. Biochem. 2000, 267, 1447–1454. [Google Scholar] [CrossRef]
- Wang, H.; Yan, X.; Yu, H.; Hu, Y.; Yu, Z.; Zheng, H.; Chen, Z.; Zhang, Z.; Liu, J. Isolation, characterization and molecular cloning of new antimicrobial peptides belonging to the brevinin-1 and temporin families from the skin of Hylarana latouchii (Anura: Ranidae). Biochimie 2009, 91, 540–547. [Google Scholar] [CrossRef]
- Wei, H.; Xie, Z.; Tan, X.; Guo, R.; Song, Y.; Xie, X.; Wang, R.; Li, L.; Wang, M.; Zhang, Y. Temporin-Like Peptides Show Antimicrobial and Anti-Biofilm Activities against Streptococcus mutans with Reduced Hemolysis. Molecules 2020, 25, 5724. [Google Scholar] [CrossRef]
- Bai, B.; Hou, X.; Wang, L.; Ge, L.; Luo, Y.; Ma, C.; Zhou, M.; Duan, J.; Chen, T.; Shaw, C. Feleucins: Novel bombinin precursor-encoded nonapeptide amides from the skin secretion of Bombina variegata. Biomed. Res. Int. 2014, 2014, 671362. [Google Scholar] [CrossRef]
- Gibson, B.W.; Tang, D.Z.; Mandrell, R.; Kelly, M.; Spindel, E.R. Bombinin-like peptides with antimicrobial activity from skin secretions of the Asian toad, Bombina orientalis. J. Biol. Chem. 1991, 266, 23103–23111. [Google Scholar] [CrossRef]
- Miele, R.; Ponti, D.; Boman, H.G.; Barra, D.; Simmaco, M. Molecular cloning of a bombinin gene from Bombina orientalis: Detection of NF-kappaB and NF-IL6 binding sites in its promoter. FEBS Lett. 1998, 431, 23–28. [Google Scholar] [CrossRef]
- Lai, R.; Liu, H.; Hui Lee, W.; Zhang, Y. An anionic antimicrobial peptide from toad Bombina maxima. Biochem. Biophys. Res. Commun. 2002, 295, 796–799. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Grovale, N.; Giorgi, A.; Mignogna, G.; Simmaco, M.; Barra, D. Structure-function relationships in bombinins H, antimicrobial peptides from Bombina skin secretions. Peptides 2000, 21, 1673–1679. [Google Scholar] [CrossRef]
- Simmaco, M.; Kreil, G.; Barra, D. Bombinins, antimicrobial peptides from Bombina species. Biochim. Biophys. Acta 2009, 1788, 1551–1555. [Google Scholar] [PubMed]
- Xiang, J.; Zhou, M.; Wu, Y.; Chen, T.; Shaw, C.; Wang, L. The synergistic antimicrobial effects of novel bombinin and bombinin H peptides from the skin secretion of Bombina orientalis. Biosci. Rep. 2017, 37, BSR20170967. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhou, C.; Hou, X.; Liu, Y.; Wang, Z.; Peng, X.; Zhang, Z.; Wang, R.; Kong, D. Molecular characterization and bioactivity evaluation of two novel bombinin peptides from the skin secretion of Oriental fire-bellied toad, Bombina orientalis. Amino Acids 2018, 50, 241–253. [Google Scholar] [CrossRef]
- Mignogna, G.; Simmaco, M.; Kreil, G.; Barra, D. Antibacterial and haemolytic peptides containing D-alloisoleucine from the skin of Bombina variegata. EMBO J. 1993, 12, 4829–4832. [Google Scholar] [CrossRef]
- Kim, S.S.; Shim, M.S.; Chung, J.; Lim, D.Y.; Lee, B.J. Purification and characterization of antimicrobial peptides from the skin secretion of Rana dybowskii. Peptides 2007, 28, 1532–1539. [Google Scholar] [CrossRef]
- Jin, L.L.; Li, Q.; Song, S.S.; Feng, K.; Zhang, D.B.; Wang, Q.Y.; Chen, Y.H. Characterization of antimicrobial peptides isolated from the skin of the Chinese frog, Rana dybowskii. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 154, 174–178. [Google Scholar] [CrossRef]
- Yang, H.; Wang, X.; Liu, X.; Wu, J.; Liu, C.; Gong, W.; Zhao, Z.; Hong, J.; Lin, D.; Wang, Y.; et al. Antioxidant peptidomics reveals novel skin antioxidant system. Mol. Cell. Proteom. 2009, 8, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hong, J.; Yang, H.; Wu, J.; Ma, D.; Li, D.; Lin, D.; Lai, R. Frog skins keep redox homeostasis by antioxidant peptides with rapid radical scavenging ability. Free. Radic. Biol. Med. 2010, 48, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Fan, Y.; Yin, S.; Li, Y.; Liu, N.; Liu, Y.; Shu, L.; Fu, Z.; Wang, Y.; Zhang, Y.; et al. Novel amphibian-derived antioxidant peptide protects skin against ultraviolet irradiation damage. J. Photochem. Photobiol. B 2021, 224, 112327. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Wang, Y.; Liu, N.; Yang, M.; Hu, Y.; Li, X.; Fu, Y.; Luo, M.; Sun, J.; Yang, X. Potential skin protective effects after UVB irradiation afforded by an antioxidant peptide from Odorrana andersonii. Biomed. Pharm. 2019, 120, 109535. [Google Scholar] [CrossRef]
- Smith, H.K.; Pasmans, F.; Dhaenens, M.; Deforce, D.; Bonte, D.; Verheyen, K.; Lens, L.; Martel, A. Skin mucosome activity as an indicator of Batrachochytrium salamandrivorans susceptibility in salamanders. PLoS ONE 2018, 13, e0199295. [Google Scholar] [CrossRef]
- Cao, X.; Wang, Y.; Wu, C.; Li, X.; Fu, Z.; Yang, M.; Bian, W.; Wang, S.; Song, Y.; Tang, J.; et al. Cathelicidin-OA1, a novel antioxidant peptide identified from an amphibian, accelerates skin wound healing. Sci. Rep. 2018, 8, 943. [Google Scholar] [CrossRef]
- Wang, X.; Ren, S.; Guo, C.; Zhang, W.; Zhang, X.; Zhang, B.; Li, S.; Ren, J.; Hu, Y.; Wang, H. Identification and functional analyses of novel antioxidant peptides and antimicrobial peptides from skin secretions of four East Asian frog species. Acta Biochim. Biophys. Sin. 2017, 49, 550–559. [Google Scholar] [CrossRef]
- Bian, W.; Meng, B.; Li, X.; Wang, S.; Cao, X.; Liu, N.; Yang, M.; Tang, J.; Wang, Y.; Yang, X. OA-GL21, a novel bioactive peptide from Odorrana andersonii, accelerated the healing of skin wounds. Biosci. Rep. 2018, 38, BSR20180215. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Zou, Z.; Yang, M.; Wu, C.; Su, Y.; Tang, J.; Yang, X. OM-LV20, a novel peptide from odorous frog skin, accelerates wound healing in vitro and in vivo. Chem. Biol. Drug. Des. 2018, 91, 126–136. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, X.; Fu, Z.; Wang, S.; Li, X.; Liu, N.; Feng, Z.; Yang, M.; Tang, J.; Yang, X. Identification and characterization of a novel gene-encoded antioxidant peptide obtained from amphibian skin secretions. Nat. Prod. Res. 2020, 34, 754–758. [Google Scholar] [CrossRef]
- Placido, A.; Bueno, J.; Barbosa, E.A.; Moreira, D.C.; Dias, J.D.N.; Cabral, W.F.; Albuquerque, P.; Bessa, L.J.; Freitas, J.; Kuckelhaus, S.A.S.; et al. The Antioxidant Peptide Salamandrin-I: First Bioactive Peptide Identified from Skin Secretion of Salamandra Genus (Salamandra salamandra). Biomolecules 2020, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Gabra, B.H.; Sirois, P. Role of bradykinin B(1) receptors in diabetes-induced hyperalgesia in streptozotocin-treated mice. Eur. J. Pharmacol. 2002, 457, 115–124. [Google Scholar] [CrossRef]
- Lai, R.; Liu, H.; Hui Lee, W.; Zhang, Y. A novel bradykinin-related peptide from skin secretions of toad Bombina maxima and its precursor containing six identical copies of the final product. Biochem. Biophys. Res. Commun. 2001, 286, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.H.; Liu, S.B.; Shen, J.H.; Jin, Y.; Zhang, Y. Cloning of bradykinin precursor cDNAs from skin of Bombina maxima reveals novel bombinakinin M antagonists and a bradykinin potential peptide. Regul. Pept. 2005, 127, 207–215. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, J.; Zhong, R.; Ma, C.; Zhou, M.; Cao, Z.; Xi, X.; Shaw, C.; Chen, T.; Wang, L.; et al. Pharmacological Effects of a Novel Bradykinin-Related Peptide (RR-18) from the Skin Secretion of the Hejiang Frog (Ordorrana hejiangensis) on Smooth Muscle. Biomedicines 2020, 8, 225. [Google Scholar] [CrossRef]
- Arichi, S.; Sasaki-Hamada, S.; Kadoya, Y.; Ogata, M.; Ishibashi, H. Excitatory effect of bradykinin on intrinsic neurons of the rat heart. Neuropeptides 2019, 75, 65–74. [Google Scholar] [CrossRef]
- Marenah, L.; Flatt, P.R.; Orr, D.F.; McClean, S.; Shaw, C.; Abdel-Wahab, Y.H. Brevinin-1 and multiple insulin-releasing peptides in the skin of the frog Rana palustris. J. Endocrinol. 2004, 181, 347–354. [Google Scholar] [CrossRef]
- Manzo, G.; Scorciapino, M.A.; Srinivasan, D.; Attoub, S.; Mangoni, M.L.; Rinaldi, A.C.; Casu, M.; Flatt, P.R.; Conlon, J.M. Conformational Analysis of the Host-Defense Peptides Pseudhymenochirin-1Pb and -2Pa and Design of Analogues with Insulin-Releasing Activities and Reduced Toxicities. J. Nat. Prod. 2015, 78, 3041–3048. [Google Scholar] [CrossRef]
- Srinivasan, D.; Ojo, O.O.; Abdel-Wahab, Y.H.; Flatt, P.R.; Guilhaudis, L.; Conlon, J.M. Insulin-releasing and cytotoxic properties of the frog skin peptide, tigerinin-1R: A structure-activity study. Peptides 2014, 55, 23–31. [Google Scholar] [CrossRef]
- Abdel-Wahab, Y.H.; Power, G.J.; Ng, M.T.; Flatt, P.R.; Conlon, J.M. Insulin-releasing properties of the frog skin peptide pseudin-2 and its [Lys18]-substituted analogue. Biol. Chem. 2008, 389, 143–148. [Google Scholar] [CrossRef]
- Conlon, J.M.; Abdel-Wahab, Y.H.; Flatt, P.R.; Leprince, J.; Vaudry, H.; Jouenne, T.; Condamine, E. A glycine-leucine-rich peptide structurally related to the plasticins from skin secretions of the frog Leptodactylus laticeps (Leptodactylidae). Peptides 2009, 30, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Li, L.; Wang, H.; Li, M.; Wang, L.; Zhou, M.; Su, Q.; Chen, T.; Wu, Y. Novel peptide dermaseptin-PS1 exhibits anticancer activity via induction of intrinsic apoptosis signalling. J. Cell. Mol. Med. 2019, 23, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Rozek, T.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. The antibiotic and anticancer active aurein peptides from the Australian Bell Frogs Litoria aurea and Litoria raniformis. Part 2. Sequence determination using electrospray mass spectrometry. Rapid. Commun. Mass. Spectrom. 2000, 14, 2002–2011. [Google Scholar] [CrossRef]
- Wiedermann, C.J. Bombesin-like peptides as growth factors. Wien Klin Wochenschr 1989, 101, 435–440. [Google Scholar] [PubMed]
- Wiedermann, C.J.; Ruff, M.R.; Pert, C.B. Bombesin-like peptides: Neuropeptides with mitogenic activity. Brain. Behav. Immun. 1988, 2, 301–310. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Chen, T.; Walker, B.; Zhou, M.; Sui, D.; Conlon, J.M.; Shaw, C. Identification and molecular cloning of a novel amphibian Bowman Birk-type trypsin inhibitor from the skin of the Hejiang Odorous Frog; Odorrana hejiangensis. Peptides 2012, 33, 245–250. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, Y.; Wang, R.; Bai, B.; Zhou, M.; Chen, T.; Cai, J.; Wang, L.; Shaw, C. PD-sauvagine: A novel sauvagine/corticotropin releasing factor analogue from the skin secretion of the Mexican giant leaf frog, Pachymedusa dacnicolor. Amino Acids 2012, 43, 1147–1156. [Google Scholar] [CrossRef]
- Baroni, A.; Perfetto, B.; Canozo, N.; Braca, A.; Farina, E.; Melito, A.; De Maria, S.; Cartenì, M. Bombesin: A possible role in wound repair. Peptides 2008, 29, 1157–1166. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, M.; Lynch, L.; Chen, T.; Walker, B.; Shaw, C. Kassina senegalensis skin tachykinins: Molecular cloning of kassinin and (Thr2, Ile9)-kassinin biosynthetic precursor cDNAs and comparative bioactivity of mature tachykinins on the smooth muscle of rat urinary bladder. Biochimie 2009, 91, 613–619. [Google Scholar] [CrossRef]
- Li, J.; Liu, T.; Xu, X.; Wang, X.; Wu, M.; Yang, H.; Lai, R. Amphibian tachykinin precursor. Biochem. Biophys. Res. Commun. 2006, 350, 983–986. [Google Scholar] [CrossRef]
- Jackway, R.J.; Pukala, T.L.; Maselli, V.M.; Musgrave, I.F.; Bowie, J.H.; Liu, Y.; Surinya-Johnson, K.H.; Donnellan, S.C.; Doyle, J.R.; Llewellyn, L.E.; et al. Disulfide-containing peptides from the glandular skin secretions of froglets of the genus Crinia: Structure, activity and evolutionary trends. Regul. Pept. 2008, 151, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob Agents Chemother 1999, 43, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Gough, M.; Hancock, R.E.; Kelly, N.M. Antiendotoxin activity of cationic peptide antimicrobial agents. Infect. Immun. 1996, 64, 4922–4927. [Google Scholar] [CrossRef] [PubMed]
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Ranatuerin-1 | Rana catesbeiana | SMLSVLKNLGKVGLGFVACKINKQC | Ee: 40.00% Cc: 30.30% | 25 | 20 (E. coli) 20 (P. aeruginosa) 40 (K. pneumoniae) 20 (E. cloacae) >100 (P. mirabilis) 20 (S. aureus) 20 (S. epidermidis) 5 (Streptococcus Group B) 20 (E. faecalis) | [64] |
| Ranatuerin-1C | Rana clamitans | SMLSVLKNLGKVGLGLVACKINKQC | Ee: 28.00% Cc: 72.00% | 25 | 1.5 (E. coli) 55 (S. aureus) 58 (C. albicans) 4 (E. coli) 17 (S. aureus) 14 (C. albicans) | [65] |
| Ranalexin-1Ca | FLGGLMKAFPALICAVTKKC | Ee: 50.00% Cc: 50.00% | 20 | |||
| Ranatuerin-1T | Rana temporaria | GLLSGLKKVGKHVAKNVAVSLMDSLKCKISGDC | Hh: 36.36% Ee: 12.12% Cc: 51.52% | 33 | 40 (E. coli) | [66] |
| 120 (S. aureus) | ||||||
| 150 (C. albicans) | ||||||
| Ranatuerin-2AUa | Rana aurora aurora | GILSSFKGVAKGVAKNLAGKLLDELKCKITGC | Hh: 53.12% Ee: 9.38% Cc: 37.50% | 32 | 5 (E. coli) | [59] |
| 5 (P. aeruginosa) | ||||||
| 10 (K. pneumoniae) | ||||||
| 5 (E. cloacae) | ||||||
| >40 (P. mirabilis) | ||||||
| 20 (S. aureus) | ||||||
| 20 (S. epidermidis) | ||||||
| 5 (Streptococcus Group B) | ||||||
| >40 (E. faecalis) | ||||||
| Ranatuerin-2BYa | Rana boylii | ILSTFKGLAKGVAKDLAGNLLDKFKCKITGC | Hh: 48.39% Ee: 19.35% Cc: 32.26% | 31 | 7 (E. coli) 27 (S. aureus) NA (C. albicans) | [50] |
| Ranatuerin-2BYb | IMDSVKGLAKNLAGKLLDSLKCKITGC | Hh: 44.44% Ee: 11.11% Cc: 44.44% | 27 | 17 (E. coli) NA (S. aureus) NA (C. albicans) | ||
| Ranatuerin-2Cb | Rana clamitans | GLFLDTLKGLAGKLLQGLKCIKAGCKP | Hh: 44.44% Ee: 7.41% Cc: 48.15% | 27 | 2 (E. coli) 40 (S. aureus) 46 (C. albicans) | [65] |
| Ranatuerin-2CSa | Rana cascadae | GILSSFKGVAKGVAKDLAG KLLETLKCKITGC | Hh: 46.88% Ee: 18.75% Cc: 34.38% | 32 | 5 (E. coli) 10 (S. aureus) | [67] |
| Ranatuerin-2Pb | Rana pipiens | SFLTTVKKLVTNLAALAGTVIDTIKCKVTGGCRT | Hh: 35.29% Ee: 32.35% Cc: 32.35% | 34 | 8 (E. coli) >256 (P. aeruginosa) 8 (S. aureus) 16 (MRSA) >512 (E. faecalis) 8 (C. albicans) | [68] |
| Ranatuerin-2YJ | Rana dybowskii | GLMDIFKVAVNKLLAAGMNKPRCKAAHC | Hh: 39.29% Ee: 17.86% Cc: 42.86% | 28 | 22.5 (E. coli) 22.5 (S. aureus) | [69] |
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Ranacyclin-B-RN1 | Hylarana nigrovittata | SALVGCWTKSYPPKPCFGR | Ee: 10.53% Cc: 89.47% | 19 | 6 (S. aureus) | [16] |
| Ranacyclin-B-RN2 | SALVGCGTKSYPPKPCFGR | Ee: 10.53% Cc: 89.47% | 19 | 12.7 (S. aureus) | ||
| Ranacyclin E | Rana temporaria | SAPRGCWTKSYPPKPCK | Ee: 35.29% Cc: 64.71% | 17 | NA (E. coli D21) 9 (Y. pseudotuberculosis YP III) 80 (P. syringae pv tabaci) 3 (B. megaterium Bm11) 7 (S. lentus) 5 (M. luteus) NA (C. albicans ATCC 10231) 7.4 (C. tropicalis) 3.4 (C. guiller-mondii) 32 (P. nicotianae spores) | [70] |
| Ranacyclin T | Rana esculenta | GALRGCWTKSYPPKPCK | Ee: 29.41% Cc: 70.59% | 17 | 30 (E. coli D21) 5 (Y. pseudotuberculosis YP III) 16 (P. syringae pv tabaci) 3 (B. megaterium Bm11) 10 (S. lentus) 8 (M. luteus) 22 (C. albicans ATCC 10231) 14 (C. tropicalis) 1 (C. guiller-mondii) 16 (P. nicotianae spores) | |
| Ranacyclin-NF | Pelophylax nigromaculatus | GAPRGCWTKSYPPQPCF | Ee: 23.53% Cc: 76.47% | 17 | >512 (E. coli) >512 (P. aeruginosa) >512 (K. pneumoniae) >512 (S. aureus) >512 (MRSA) >512 (E. faecalis) | [72] |
| Ranacyclin-NF1 | GAPRGCWTKSYPPQPCF | Ee: 23.53% Cc: 76.47% | 17 | >512 (E. coli) >512 (P. aeruginosa) >512 (K. pneumoniae) >512 (S. aureus) >512 (MRSA) >512 (E. faecalis) | ||
| Ranacyclin-NF3L | GALRGCWTKSYPPQPCF | Ee: 23.53% Cc: 76.47% | 17 | >512 (E. coli) >512 (P. aeruginosa) >512 (K. pneumoniae) >512 (S. aureus) >512 (MRSA) >512 (E. faecalis) |
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Temporin-1BYa | Rana boylii | FLPIIAKVLSGLL | Ee: 61.54% Cc: 38.46% | 13 | NA (E. coli) 15 (S. aureus) NA (C. albicans) | [50] |
| Temporin-1Gb | Rana grylio | SILPTIVSFLSKFL | Ee: 42.86% Cc: 57.14% | 14 | NA (E. coli) 24 (S. aureus) NA (C. albicans) | [76] |
| Temporin-1Gc | SILPTIVSFLTKFL | Ee: 57.14% Cc: 42.86% | 14 | NA (E. coli) 25 (S. aureus) NA (C. albicans) | ||
| Temporin-1Gd | FILPLIASFLSKFL | Ee: 14.29% Cc: 85.71% | 14 | NA (E. coli) 12 (S. aureus) NA (C. albicans) | ||
| Temporin-LT1 | Hylarana latouchii | FLPGLIAGIAKML | Ee: 38.46% Cc: 61.54% | 13 | NA (P. fluorescens) 12.5 (S. aureus) 25 (B. subtilis) NA (C. albicans) | [88] |
| Temporin-LT2 | FLPIALKALGSIFPKIL | Ee: 29.41% Cc: 70.59% | 17 | NA (P. fluorescens) 12.5 (S. aureus) 50 (B. subtilis) NA (C. albicans) | ||
| Temporin-GHaR | Hylarana guentheri | FLQRIIGALGRLF | Ee: 61.54% Cc: 38.46% | 13 | 6.2 (E. coli) 12.5 (E. coli D31) 6.2 (P. aeruginosa) 25 (P. aeruginosa PAO1) 1.6 (S. aureus) 12.5 (B. subtilis) 3.1 (S. mutans) 3.1 (MRSA) 12.5 (C. albicans) | [89] |
| Temporin-GHaR6R | FLQRIRGALGRLF | Ee: 53.85% Cc: 46.15% | 13 | 12.5 (E. coli) 25 (E. coli D31) >50 (P. aeruginosa) 3.1 (P. aeruginosa PAO1) 3.1 (S. aureus) 25 (B. subtilis) 12.5 (S. mutans) 6.2 (MRSA) 50 (C. albicans) | ||
| Temporin-GHaR7R | FLQRIIRALGRLF | Ee: 53.85% Cc: 46.15% | 13 | 3.1 (E. coli) 12.5 (E. coli D31) >50 (P. aeruginosa) >50 (P. aeruginosa PAO1) 3.1 (S. aureus) >50 (B. subtilis) 6.2 (S. mutans) 3.1 (MRSA) 25 (C. albicans) | ||
| Temporin-GHaR8R | FLQRIIGRLGRLF | Ee: 61.54% Cc: 38.46% | 13 | 3.1 (E. coli) 12.5 (E. coli D31) 50 (P. aeruginosa) 6.2 (P. aeruginosa PAO1) 1.6–3.1 (S. aureus) 12.5 (B. subtilis) 6.2 (S. mutans) 3.1 (MRSA) 12.5 (C. albicans) | ||
| Temporin-GHaR9R | FLQRIIGARGRLF | Ee: 53.85% Cc: 46.15% | 13 | >50 (E. coli) >50 (E. coli D31) >50 (P. aeruginosa) >50 (P. aeruginosa PAO1) 6.2 (S. aureus) >50 (B. subtilis) >50 (S. mutans) >50 (MRSA) >50 (C. albicans) | ||
| Temporin-GHaR9W | FLQRIIGAWGRLF | Ee: 53.85% Cc: 46.15% | 13 | 12.5 (E. coli) 12.5 (E. coli D31) >50 (P. aeruginosa) 25 (P. aeruginosa PAO1) 3.1 (S. aureus) 12.5 (B. subtilis) 6.2 (S. mutans) 6.2 (MRSA) 25 (C. albicans) |
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | MIC (μM) | Ref. |
|---|---|---|---|---|---|---|
| Dybowskin-1 | Rana dybowskii | FLIGMTHGLICLISRKC | Ee: 47.06% Cc: 52.94% | 17 | >52.5 (E. coli) >52.5 (K. pneumoniae) >52.5 (P. mirabilis) >52.5 (P. aeruginosa) 13.1 (S. aureus) 26.3 (B. subtilis) 31.5 (S. epidermidis) >52.5 (S. dysenteriae) 6.6 (M. luteus) >52.5 (C. albicans) | [99] |
| Dybowskin-2 | FLIGMTQGLICLITRKC | Ee: 47.06% Cc: 52.94% | 17 | 31.4 (E. coli) 31.4 (K. pneumoniae) >52.4 (P. mirabilis) >52.4 (P. aeruginosa) 7.9 (S. aureus) 13.1 (B. subtilis) 13.1 (S. epidermidis) 26.2 (S. dysenteriae) 3.3 (M. luteus) 52.4 (C. albicans) | ||
| Dybowskin-3 | GLFDVVKGVLKGVGKNVAGSLLEQLKCKLSGGC | Hh: 27.27% Ee: 39.39% Cc: 33.33% | 33 | 4.5 (E. coli) 4.5 (K. pneumoniae) >30.2 (P. mirabilis) >30.2 (P. aeruginosa) 9 (S. aureus) 15.1 (B. subtilis) 18.1 (S. epidermidis) 18.1 (S. dysenteriae) 1.9 (M. luteus) 30.2 (C. albicans) | ||
| Dybowskin-4 | VWPLGLVICKALKIC | Ee: 33.33% Cc: 66.67% | 15 | 9.1 (E. coli) 9.1 (K. pneumoniae) >60.4 (P. mirabilis) >60.4 (P. aeruginosa) 1.9 (S. aureus) 3.8 (B. subtilis) 3.8 (S. epidermidis) 18.1 (S. dysenteriae) 0.9 (M. luteus) 15.1 (C. albicans) | ||
| Dybowskin-5 | GLFSVVTGVLKAVGKNVAKNVGGSLLEQLKCKISGGC | Hh: 24.32% Ee: 35.14% Cc: 40.54% | 37 | 16.8 (E. coli) 13.6 (K. pneumoniae) >27.2 (P. mirabilis) >27.2 (P. aeruginosa) 6.8 (S. aureus) 13.6 (B. subtilis) 13.6 (S. epidermidis) 13.6 (S. dysenteriae) 1.8 (M. luteus) 13.6 (C. albicans) | ||
| Dybowskin-6 | FLPLLLAGLPLKLCFLFKKC | Ee: 60.00% Cc: 40.00% | 20 | >44 (E. coli) >44 (K. pneumoniae) >44 (P. mirabilis) >44 (P. aeruginosa) 5.5 (S. aureus) 22 (B. subtilis) 22 (S. epidermidis) >44 (S. dysenteriae) 2.7 (M. luteus) 22 (C. albicans) | ||
| Dybowskin-1CDYa | Rana dybowskii | IIPLPLGYFAKKT | Ee: 15.38% Cc: 84.62% | 13 | 3 (E. coli) 6 (S. aureus) | [100] |
| Dybowskin-2CDYa | SAVGRHGRRFGLRKHRKH | Ee: 27.78% Cc: 72.22% | 18 | 3 (E. coli) 6 (S. aureus) |
| Peptides Name | Species | Primary Structure | Secondary Structure | Length | Ref. |
|---|---|---|---|---|---|
| Andersonin-C1 | Odorrana margaratae | TSRCIFYRRKKCS | Ee: 53.85% Cc: 46.15% | 13 | [105] |
| Andersonin-G1 | Odorrana andersonii | KEKLKLKAKAP KCYNDKLACT | Ee: 23.81% Cc: 76.19% | 21 | |
| Andersonin-H3 | Odorrana margaratae | VAIYGRDDRSDVCR QVQHNWLVCDTY | Ee: 42.31% Cc: 57.69% | 26 | |
| Antioxidin-RP1 | Rana pleuraden | AMRLTYNKPCLYGT | Ee: 28.57% Cc: 71.43% | 14 | [101,105] |
| Cathelicidin-OA1 | Odorrana andersonii | IGRDPTWSHLAASCLKCIFDDLPKTHN | Hh: 29.63% Ee: 7.41% Cc: 62.96% | 27 | [106] |
| Nigroain-B-MS1 | Hylarana maosuoensis | CVVSSGWKWNY KIRCKLTGNC | Ee: 47.62% Cc: 52.38% | 21 | [107] |
| OA-VI12 | Odorrana andersonii | VIPFLACRPLGL | Ee: 50.00% Cc: 50.00% | 12 | [104] |
| OA-GL21 | Odorrana andersonii | GLLSGHYGRVVSTQSGHYGRG | Ee: 52.38% Cc: 47.62% | 21 | [108] |
| OM-LV20 | Odorrana margaretae | LVGKLLKGAVGDVCGLLPIC | Ee: 45.00% Cc: 55.00% | 20 | [109] |
| OM-GF17 | Odorrana margaretae | GFFKWHPRCGEEHSMWT | Ee: 35.29% Cc: 64.71% | 17 | [110] |
| OS-LL11 | Odorrana schmackeri | LLPPWLCPRNK | Cc: 100.00% | 11 | [103] |
| Pleurain-A1 | Rana pleuraden | SIITMTKEAKLPQLW KQIACRLYNTC | Hh: 19.23% Ee: 30.77% Cc: 50.00% | 26 | [101] |
| Pleurain-D1 | Rana pleuraden | FLSGILKLAFKIP SVLCAVLKNC | Ee: 47.83% Cc: 52.17% | 23 | |
| Pleurain-E1 | Rana pleuraden | AKAWGIPPHVIPQI VPVRIRPLCGNV | Ee: 30.77% Cc: 69.23% | 26 | |
| Salamandrin-I | Salamandra salamandra | FAVWGCADYRGY | Ee: 58.33% Cc: 41.67% | 12 | [111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Liu, S.; Fang, J.; Zheng, S.; Wang, Z.; Jiao, Y.; Xia, P.; Wu, H.; Ma, Z.; Hao, L. Peptides Isolated from Amphibian Skin Secretions with Emphasis on Antimicrobial Peptides. Toxins 2022, 14, 722. https://doi.org/10.3390/toxins14100722
Chen X, Liu S, Fang J, Zheng S, Wang Z, Jiao Y, Xia P, Wu H, Ma Z, Hao L. Peptides Isolated from Amphibian Skin Secretions with Emphasis on Antimicrobial Peptides. Toxins. 2022; 14(10):722. https://doi.org/10.3390/toxins14100722
Chicago/Turabian StyleChen, Xi, Songcai Liu, Jiayuan Fang, Shuo Zheng, Zhaoguo Wang, Yingying Jiao, Peijun Xia, Hongyan Wu, Ze Ma, and Linlin Hao. 2022. "Peptides Isolated from Amphibian Skin Secretions with Emphasis on Antimicrobial Peptides" Toxins 14, no. 10: 722. https://doi.org/10.3390/toxins14100722
APA StyleChen, X., Liu, S., Fang, J., Zheng, S., Wang, Z., Jiao, Y., Xia, P., Wu, H., Ma, Z., & Hao, L. (2022). Peptides Isolated from Amphibian Skin Secretions with Emphasis on Antimicrobial Peptides. Toxins, 14(10), 722. https://doi.org/10.3390/toxins14100722