In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds

 , , , and
, , , and
Abstract
1. Introduction
2. Results and Discussion
2.1. Identification of Volatile Organic Compounds Produced by Antagonistic Yeasts
2.2. Influence of VOCs on Growth Parameters of Aspergillus Flavus
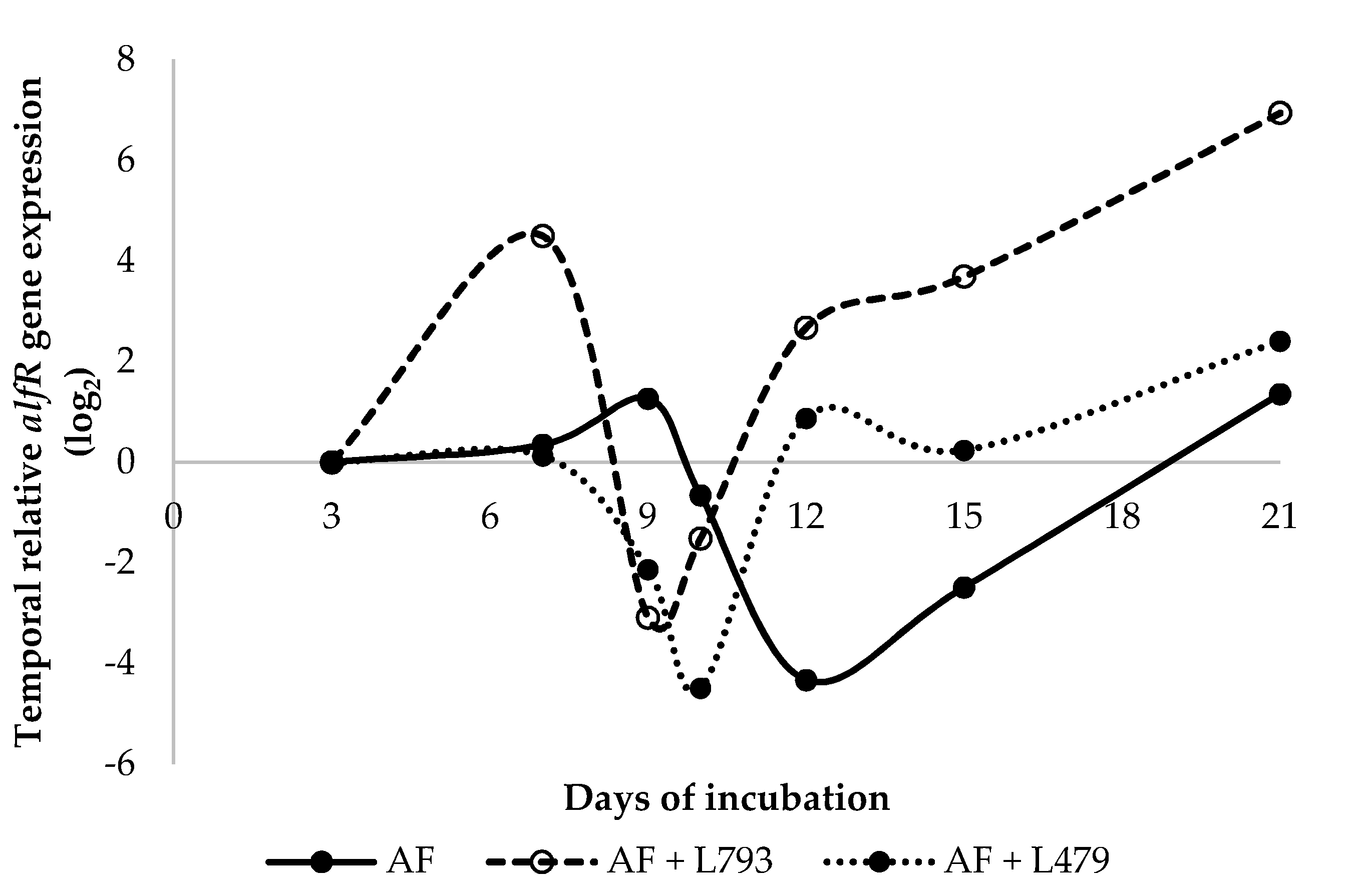
2.3. Gene Expression
2.4. Aflatoxin Amounts
3. Conclusions
4. Materials and Methods
4.1. VOC-Producing Yeasts
4.2. Aspergillus Flavus Strain
4.3. Competitiveness of VOC-Producing Yeasts against Aspergillus Flavus Assessed Using Double-Dish Systems
4.4. Analysis of Volatile Compounds
4.5. Determination of Growth Parameters of Aspergillus Flavus
4.6. Relative Quantification of the Expression of the aflR Gene
4.6.1. Sample Preparation
4.6.2. RNA Extraction
4.6.3. Two-Step Reverse-Transcription Real-Time PCR
Retrotranscription Reaction
Real-Time PCR Reactions
4.6.4. Calculation of Relative Gene Expression
4.7. Aflatoxin Analysis
4.8. Statistical Analysis of Results
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdel-Wahhab, M.A.; Ahmed, H.H.; Hagazi, M.M. Prevention of aflatoxin B1-initiated hepatotoxicity in rat by marine algae extracts. J. Appl. Toxicol. 2006, 26, 229–238. [Google Scholar] [CrossRef]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Scientific Committee for Food. Reports of the Scientific Committee for Food. In Opinion of the Scientific Committee for Food on Aflatoxins, Ochratoxin A and Patulin; 35th Series; The European Union: Luxembourg, 1994; p. 45. [Google Scholar]
- Wang, J.; Tang, L.; Glenn, T.C.; Wang, J.S. Aflatoxin B1 induced compositional changes in gut microbial communities of male F344 rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef]
- Shuaib, F.M.; Ehiri, J.; Abdullahi, A.; Williams, J.H.; Jolly, P.E. Reproductive health effects of aflatoxins: A review of the literature. Reprod. Toxicol. 2010, 29, 262–270. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). Summaries and Evaluations: Aflatoxins; IARC Press: Lyon, France, 2002; Volume 82, p. 171. [Google Scholar]
- He, X.Y.; Tang, L.; Wang, S.L.; Cai, Q.S.; Wang, J.S.; Hong, J.Y. Efficient activation of aflatoxin B1 by cytochrome P450 2A13, an enzyme predominantly expressed in human respiratory tract. Int. J. Cancer 2006, 118, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Magnussen, A.; Parsi, M.A. Aflatoxins, hepatocellular carcinoma and public health. World J. Gastroenterol. 2013, 19, 1508. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenár, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Giorni, P.A.O.L.A.; Magan, N.; Pietri, A.M.E.D.E.O.; Bertuzzi, T.; Battilani, P. Studies on Aspergillus section Flavi isolated from maize in northern Italy. Int. J. Food Microbiol. 2007, 113, 330–338. [Google Scholar] [CrossRef]
- Lai, X.; Zhang, H.; Liu, R.; Liu, C. Potential for aflatoxin B1 and B2 production by Aspergillus flavus strains isolated from rice samples. Saudi J. Biol. Sci. 2015, 22, 176–180. [Google Scholar] [CrossRef]
- Casquete, R.; Rodríguez, A.; Hernández, A.; Martín, A.; Bartolomé, T.; Córdoba, J.J.; Córdoba, M.G. Occurrence of toxigenic fungi and mycotoxins during smoked paprika production. J. Food Protect. 2017, 80, 2068–2077. [Google Scholar] [CrossRef]
- Iamanaka, B.T.; de Menezes, H.C.; Vicente, E.; Leite, R.S.; Taniwaki, M.H. Aflatoxigenic fungi and aflatoxins occurrence in sultanas and dried figs commercialized in Brazil. Food Control 2007, 18, 454–457. [Google Scholar] [CrossRef]
- Kosegarten, C.E.; Ramírez-Corona, N.; Mani-López, E.; Palou, E.; López-Malo, A. Description of Aspergillus flavus growth under the influence of different factors (water activity, incubation temperature, protein and fat concentration, pH, and cinnamon essential oil concentration) by kinetic, probability of growth, and time-to-detection models. Int. J. Food Microbiol. 2017, 240, 115–123. [Google Scholar] [PubMed]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. Royal Soc. Interface. 2012, 9, 757–767. [Google Scholar] [CrossRef]
- Bernáldez, V.; Córdoba, J.J.; Magan, N.; Peromingo, B.; Rodríguez, A. The influence of ecophysiological factors on growth, aflR gene expression and aflatoxin B1 production by a type strain of Aspergillus flavus. LWT-Food Sci. Technol. 2017, 83, 283–291. [Google Scholar] [CrossRef]
- Galván, A.I.; Rodríguez, A.; Martín, A.; Serradilla, M.J.; Martínez-Dorado, A.; Córdoba, M.G. Effect of temperature during drying and storage of dried figs on growth, gene expression and aflatoxin production. Toxins 2021, 13, 134. [Google Scholar] [CrossRef]
- Passone, M.A.; Ruffino, M.; Ponzio, V.; Resnik, S.; Etcheverry, M.G. Postharvest control of peanut Aspergillus section Flavi populations by a formulation of food-grade antioxidants. Int. J. Food Microbiol. 2009, 131, 211–217. [Google Scholar] [CrossRef]
- Dikbas, N.; Kotan, R.; Dadasoglu, F.; Sahin, F. Control of Aspergillus flavus with essential oil and methanol extract of Satureja hortensis. Int. J. Food Microbiol. 2008, 124, 179–182. [Google Scholar] [CrossRef]
- Omidbeygi, M.; Barzegar, M.; Hamidi, Z.; Naghdibadi, H. Antifungal activity of thyme, summer savory and clove essential oils against Aspergillus flavus in liquid medium and tomato paste. Food Control 2007, 18, 1518–1523. [Google Scholar] [CrossRef]
- Tian, J.; Huang, B.; Luo, X.; Zeng, H.; Ban, X.; He, J.; Wang, Y. The control of Aspergillus flavus with Cinnamomum jensenianum Hand.-Mazz essential oil and its potential use as a food preservative. Food Chem. 2012, 130, 520–527. [Google Scholar] [CrossRef]
- Ferreira, F.D.; Kemmelmeier, C.; Arrotéia, C.C.; da Costa, C.L.; Mallmann, C.A.; Janeiro, V.; Ferreira, F.M.D.; Mossini, S.A.G.; Silva, E.L.; Machinski Jr, M. Inhibitory effect of the essential oil of Curcuma longa L. and curcumin on aflatoxin production by Aspergillus flavus Link. Food Chem. 2013, 136, 789–793. [Google Scholar] [CrossRef] [PubMed]
- da Cruz Cabral, L.; Fernández-Pino, V.; Patriarca, A. Control of infection of tomato fruits by Alternaria and mycotoxin production using plant extracts. Eur. J. Plant Pathol. 2016, 145, 363–373. [Google Scholar] [CrossRef]
- Bueno, D.J.; Silva, J.O.; Oliver, G.; Gonzalez, S.N. Lactobacillus casei CRL 431 and Lactobacillus rhamnosus CRL 1224 as biological controls for Aspergillus flavus strains. J. Food Protect. 2006, 69, 2544–2548. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Z.Q.; Hu, L.B.; Cheng, L.G.; Wang, F. Antifungal compounds from Bacillus subtilis B-FS06 inhibiting the growth of Aspergillus flavus. World J. Microbiol. Biotechnol. 2008, 24, 783. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Bruns, H.A.; Abel, C.A. Biocontrol of aflatoxin in corn by inoculation with non-aflatoxigenic Aspergillus flavus isolates. Biocontrol Sci. Technol. 2006, 16, 437–449. [Google Scholar] [CrossRef]
- Dorner, J.W. Biological control of aflatoxin contamination in corn using a nontoxigenic strain of Aspergillus flavus. J. Food Protect. 2009, 72, 801–804. [Google Scholar] [CrossRef]
- Persons, K.; Raines, J.M.; Rodriguez, J.M. Antagonistic effects of Saccharomyces cerevisiae on the growth of Aspergillus flavus and Aspergillus parasiticus at varying temperatures. Mycology 2013, 4, 38–43. [Google Scholar] [CrossRef]
- Hua, S.S.T. Progress in prevention of aflatoxin contamination in food by preharvest application of a yeast strain, Pichia anomala WRL-076. Mod. Multidiscip. Appl. Microbiol. Exploit. Microbes Interact. 2006, 322–326. [Google Scholar] [CrossRef]
- Jaibangyang, S.; Nasanit, R.; Limtong, S. Biological control of aflatoxin-producing Aspergillus flavus by volatile organic compound-producing antagonistic yeasts. BioControl 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Patiño, B.; Cortés, L.; González-Jaén, M.T.; Vázquez, C. Mechanisms involved in reduction of ochratoxin A produced by Aspergillus westerdijkiae using Debaryomyces hansenii CYC 1244. Int. J. Food Microbiol. 2011, 151, 113–118. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Rosend, J.; Kuldjärv, R.; Rosenvald, S.; Paalme, T. The effects of apple variety, ripening stage, and yeast strain on the volatile composition of apple cider. Heliyon 2019, 5, e01953. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, T.; Ishihara, A.; Nakajima, H. Effects of volatile compounds in apple juices on fungal growth and patulin production of Penicillium expansum. JSM Mycotoxins 2014, 64, 1–14. [Google Scholar] [CrossRef][Green Version]
- de Arruda Moura, G.; dos Santos, C.M.E.; Sauer, E.; Wosiacki, G.; Nogueira, A. Influence of fermentation with Hanseniaspora sp. yeast on the volatile profile of fermented apple. J. Agric. Food Chem. 2012, 60, 9815–9821. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, G.A.; Wang, L.P. Controlled formation of volatile components in cider making using a combination of Saccharomyces cerevisiae and Hanseniaspora valbyensis yeast species. J. Indust. Microbiol. Biotechnol. 2006, 33, 192–196. [Google Scholar] [CrossRef]
- Moreira, N.; Pina, C.; Mendes, F.; Couto, J.A.; Hogg, T.; Vasconcelos, I. Volatile compounds contribution of Hanseniaspora guilliermondii and Hanseniaspora uvarum during red wine vinifications. Food Control 2011, 22, 662–667. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Hernández, A.; Galvan, A.I.; Córdoba, M.G.; Casquete, R.; Serradilla, M.J.; Martín, A. Selection and application of antifungal VOCs-producing yeasts as biocontrol agents of grey mould in fruits. Food Microbio. 2020, 92, 103556. [Google Scholar] [CrossRef]
- Ando, H.; Hatanaka, K.; Ohata, I.; Yamashita-Kitaguchi, Y.; Kurata, A.; Kishimoto, N. Antifungal activities of volatile substances generated by yeast isolated from Iranian commercial cheese. Food Control 2012, 26, 472–478. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism of action against postharvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile organic compounds (VOCs) produced by biocontrol yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef]
- Plata, C.; Millán, C.; Mauricio, J.C.; Ortega, J.M. Formation of ethyl acetate and isoamyl acetate by various species of wine Yeats. Food Microbiol. 2003, 20, 217–224. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Payne, G. Genetic regulation of aflatoxin biosynthesis: From gene to genome. Fungal Genet. Biol. 2009, 26, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Al-Saad, L.A.; Al-Badran, A.I.; Al-Jumayli, S.A.; Magan, N.; Rodríguez, A. Impact of bacterial biocontrol agents on aflatoxin biosynthetic genes, aflD and aflR expression, and phenotypic aflatoxin B1 production by Aspergillus flavus under different environmental and nutritional regimes. Int. J. Food Microbiol. 2016, 217, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Rodríguez, M.; Delgado, J.; Andrade, M.J.; Rodríguez, A. Gene expression as a good indicator of aflatoxin contamination in dry-cured ham. Food Microbiol. 2017, 67, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Ojalvo, D.; Rodríguez, A.; Bernáldez, V.; Córdoba, J.J.; Rodríguez, M. Influence of temperature and substrate conditions on the omt-1 gene expression of Aspergillus parasiticus in relation to its aflatoxin production. Int. J. Food Microbiol. 2013, 166, 263–269. [Google Scholar] [CrossRef]
- Rodríguez, A.; Medina, A.; Córdoba, J.J.; Magan, N. The influence of salt (NaCl) on ochratoxin A biosynthetic genes, growth and ochratoxin A production by three strains of Penicillium nordicum on a dry-cured ham-based medium. Int. J. Food Microbiol. 2014, 178, 113–119. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Abdel-Hadi, A.; Magan, N.; Geisen, R. Complex regulation of the aflatoxin biosynthesis gene cluster of Aspergillus flavus in relation to various combinations of water activity and temperature. Int. J. Food Microbiol. 2009, 135, 231–237. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Magan, N.; Geisen, R. Stress induction of mycotoxin biosynthesis genes by abiotic factors. FEMS Microbiol. Lett. 2008, 284, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Rüfer, C.E.; Abdel-Hadi, A.; Magan, N.; Geisen, R. The production of aflatoxin B1 or G1 by Aspergillus parasiticus at various combinations of temperature and water activity is related to the ratio aflS to aflR expression. Mycotoxin Res. 2010, 26, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Peromingo, B.; Andrade, M.J.; Delgado, J.; Sánchez-Montero, L.; Núñez, F. Biocontrol of aflatoxigenic Aspergillus parasiticus by native Debaryomyces hansenii in dry-cured meat products. Food Microbiol. 2019, 82, 269–276. [Google Scholar] [CrossRef]
- León, A.M.; Cataño, C.S.; Yepes, E.Q.; Villarroel, R.G.; De Antoni, G.L.; Giannuzzi, L. Inhibitory activity of lactic and acetic acid on Aspergillus flavus growth for food preservation. Food Control 2012, 24, 177–183. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Moyano, S.; Benito, M.J.; Martín, A.; Aranda, E.; Hernández, A.; Córdoba, M.G. Characterization of molds isolated from smoked paprika by PCR-RFLP and micellar electrokinetic capillary electrophoresis. Food Microbiol. 2009, 26, 776–782. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volatile Compounds | Family a | KI b | Peak Number | Retention Time | Mean Relative Abundances | p | ||
|---|---|---|---|---|---|---|---|---|
| AF c | AF + L479 d | AF + L793 e | ||||||
| 1-Propanol | ol | 554 | v4 | 5.1 | 0.28 | 0.00 * | 0.08 * | 0.032 |
| 2-Methylpentane | h | 560 | v5 | 5.3 | 2.29 | 0.73 * | 0.23 * | 0.024 |
| 3-Methylpentane | h | 581 | v6 | 5.8 | 2.58 | 0.95 * | 0.25 * | 0.050 |
| Acetic acid | ac | 595 | v9 | 6.8 | 0.00 | 43.38 + | 0.00 | 0.000 |
| Ethyl acetate | es | 612 | v10 | 6.8 | 1.89 | 2.59 + | 18.184 + | 0.023 |
| Propanoic acid | ac | 705 | v20 | 10.7 | 0.09 | 0.22 + | 0.24 + | 0.008 |
| n-Propyl acetate | es | 706 | v22 | 11.0 | 0.00 | 0.35 + | 0.70 + | 0.049 |
| Isoamyl alcohol | ol | 732 | v26 | 11.9 | 9.54 | 0.81 * | 12.77 + | 0.001 |
| 2-Methyl-1-butanol | ol | 740 | v27 | 12.1 | 4.36 | 0.43 * | 11.54 + | 0.000 |
| Isobutyric acid | ac | 775 | v29 | 12.8 | 0.84 | 1.45 + | 0.41 | 0.021 |
| 2-Methylbutanoic acid | ac | 854 | v42 | 16.8 | 0.59 | 6.91 + | 0.28 | 0.017 |
| 1,3-Dimethylbenzene | b | 875 | v44 | 18.8 | 0.72 | 0.31 * | 0.10 * | 0.047 |
| Isoamyl acetate | es | 878 | v45 | 18.9 | 0.33 | 0.32 | 9.60 + | 0.043 |
| 2-methylbutyl acetate | es | 880 | v46 | 19.0 | 0.61 | 0.30 | 3.27 + | 0.039 |
| 2-Heptanone | k | 889 | v48 | 19.5 | 0.00 | 0.03 + | 0.30 + | 0.007 |
| Furfuryl acetate | f | 997 | v57 | 24.4 | 0.00 | 0.00 | 0.26 + | 0.006 |
| Hexyl acetate | es | 1010 | v58 | 25.1 | 0.00 | 0.03 + | 0.13 + | 0.048 |
| Ethyl heptanoate | es | 1085 | v67 | 28.7 | 0.00 | 0.03 + | 0.08 + | 0.048 |
| Nonanal | al | 1104 | v68 | 29.1 | 0.20 | 0.08 * | 0.23 + | 0.046 |
| Phenethyl alcohol | b | 1110/1141 | v69 | 29.8 | 0.00 | 0.00 | 4.56 + | 0.007 |
| Phenylmethyl acetate | es | 1170 | v70 | 31.6 | 0.00 | 0.00 | 0.13 + | 0.043 |
| 2-Phenylethyl acetate | es | 1258 | v76 | 35.0 | 3.02 | 1.48 | 8.46 + | 0.045 |
| Treatment | Diameter of Mycelium (mm) | µ (mm/Day) | λ (Days) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Days of Incubation | |||||||||
| 3 | 7 | 9 | 10 | 12 | 15 | 21 | |||
| AF | 13.55 ± 0.52c 1 | 34.50 ± 1.11c | 43.72 ± 0.75b | 47.50 ± 0.74c | 57.55 ± 1.83c | 70.83 ± 0.96b | 75.20 ± 0.44b | 4.58 ± 0.03c | 0.58 ± 0.04a |
| AF + L479 | 12.00 ± 0.50b | 29.74 ± 0.97b | 37.95 ± 1.84a | 39.37 ± 0.99b | 50.26 ± 4.18b | 63.87 ± 4.38b | 73.20 ± 2.38b | 4.00 ± 0.08b | 0.87 ± 0.10b |
| AF + L793 | 8.88 ± 1.26a | 25.39 ± 1.93a | 32.36 ± 2.60a | 35.55 ± 2.85a | 42.81 ± 3.47a | 52.00 ± 5.13a | 57.00 ± 7.37a | 3.54 ± 0.08a | 1.07 ± 0.08b |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.015 | <0.001 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejero, P.; Martín, A.; Rodríguez, A.; Galván, A.I.; Ruiz-Moyano, S.; Hernández, A. In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds. Toxins 2021, 13, 663. https://doi.org/10.3390/toxins13090663
Tejero P, Martín A, Rodríguez A, Galván AI, Ruiz-Moyano S, Hernández A. In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds. Toxins. 2021; 13(9):663. https://doi.org/10.3390/toxins13090663
Chicago/Turabian StyleTejero, Paula, Alberto Martín, Alicia Rodríguez, Ana Isabel Galván, Santiago Ruiz-Moyano, and Alejandro Hernández. 2021. "In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds" Toxins 13, no. 9: 663. https://doi.org/10.3390/toxins13090663
APA StyleTejero, P., Martín, A., Rodríguez, A., Galván, A. I., Ruiz-Moyano, S., & Hernández, A. (2021). In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds. Toxins, 13(9), 663. https://doi.org/10.3390/toxins13090663