Activity of Some Plant and Fungal Metabolites towards Aedes albopictus (Diptera, Culicidae)


 ,
,  and
and
Abstract
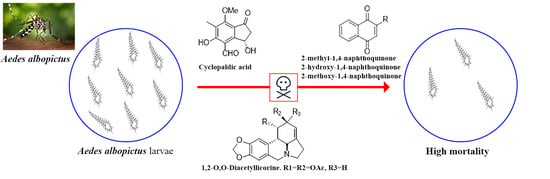
1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insects
5.2. Natural Compounds Tested
5.3. Larvicidal Tests
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface 2012, 9, 2708–2717. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Bortel, W.V. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. The changing epidemiology of yellow fever and dengue, 1900 to 2003: Full circle? Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Faye, O.; Freire, C.C.; Iamarino, A.; Faye, O.; de Oliveira, J.V.; Diallo, M.; Zanotto, P.M.A.; Sall, A.A. Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl. Trop. Dis. 2014, 8, e2636. [Google Scholar] [CrossRef]
- Dasti, J.I. Zika virus infections: An overview of current scenario. Asian Pac. J. Trop. Med. 2016, 9, 621–625. [Google Scholar] [CrossRef]
- Li, R.; Ding, J.; Ding, G.; Fan, X.; He, Y.; Wang, X.; Zhang, H.; Ji, J.; Li, H. Zika virus infections, a review. Radiol. Infect. Dis. 2017, 4, 88–93. [Google Scholar] [CrossRef]
- Townson, H.; Nathan, M.B.; Zaim, M.; Guillet, P.; Manga, L.; Bos, R.; Kindhauser, M. Exploiting the potential of vector control for disease prevention. B World Health Organ. 2005, 83, 942–947. [Google Scholar]
- Alphey, L.; Benedict, M.; Bellini, R.; Clark, G.G.; Dame, D.A.; Service, M.W.; Dobson, S.L. Sterile-insect methods for control of mosquito-borne diseases: An analysis. Vector Borne Zoonotic Dis. 2010, 10, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Rainey, S.M.; Shah, P.; Kohl, A.; Dietrich, I. Understanding the Wolbachia-mediated inhibition of arboviruses in mosquitoes: Progress and challenges. J. Gen. Virol. 2014, 95, 517–530. [Google Scholar] [CrossRef]
- McGraw, E.A.; O’Neill, S.L. Beyond insecticides: New thinking on an ancient problem. Nat. Rev. Microbiol. 2013, 11, 181–193. [Google Scholar] [CrossRef]
- Bellini, R.; Albieri, A.; Balestrino, F.; Carrieri, M.; Porretta, D.; Urbanelli, S.; Calvitti, M.; Moretti, R.; Maini, S. Dispersal and survival of Aedes albopictus (Diptera: Culicidae) males in Italian urban areas and significance for sterile insect technique application. J. Med. Entomol. 2010, 47, 1082–1091. [Google Scholar] [CrossRef]
- Bourtzis, K.; Dobson, S.L.; Xi, Z.; Rasgon, J.L.; Calvitti, M.; Moreira, L.A.; Bossin, H.C.; Moretti, R.; Baton, L.A.; Hughes, G.L.; et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. 2014, 132, S150–S163. [Google Scholar] [CrossRef]
- Mancini, M.V.; Herd, C.S.; Ant, T.H.; Murdochy, S.M.; Sinkins, S.P. Wolbachia strain wAu efficiently blocks arbovirus transmission in Aedes albopictus. PLoS. Negl. Trop. Dis. 2020, 14, e0007926. [Google Scholar] [CrossRef] [PubMed]
- Lees, R.S.; Gilles, J.R.L.; Hendrichs, J.; Vreysen, M.J.B.; Bourtzis, K. Back to the future: The sterile insect technique against mosquito disease vectors. Curr. Opin. Insect. Sci. 2015, 10, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Weeratunga, P.; Rodrigo, C.; Fernando, S.D.; Rajapakse, S. Control methods for Aedes albopictus and Aedes aegypti (Protocol)’. Cochrane Database Syst Rev. 2017, 8. [Google Scholar] [CrossRef]
- Grigoraki, L.; Pipini, D.; Labbé, P.; Chaskopoulou, A.; Weill, M.; Vontas, J. Carboxylesterase gene amplifications associated with insecticide resistance in Aedes albopictus: Geographical distribution and evolutionary origin. PLoS Negl. Trop. Dis. 2017, 11, e0005533. [Google Scholar] [CrossRef] [PubMed]
- Dusfour, I.; Vontas, J.; David, J.P.; Weetman, D.; Fonseca, D.M.; Corbel, V.; Raghavendra, K.; Coulibaly, M.B.; Martins, A.J.; Kasai, S.; et al. Management of insecticide resistance in the major Aedes vectors of arboviruses: Advances and challenges. PLoS Negl Trop Dis. 2019, 13, e0007615. [Google Scholar] [CrossRef]
- Balde, E.S.; Andolfi, A.; Bruyère, C.; Cimmino, A.; Lamoral-Theys, D.; Vurro, M.; Van Damme, M.; Altomare, C.; Mathieu, V.; Kiss, R.; et al. Investigations of fungal secondary metabolites with potential anticancer activity. J. Nat. Prod. 2010, 73, 969–971. [Google Scholar] [CrossRef]
- Schrader, K.K.; Andolfi, A.; Cantrell, C.L.; Cimmino, A.; Duke, S.O.; Osbrink, W.; Wedge, D.E.; Evidente, A. A survey of phytotoxic microbial and plant metabolites as potential natural products for pest management. Chem. Biodivers. 2010, 7, 2261–2280. [Google Scholar] [CrossRef]
- Van Goietsenoven, G.; Andolfi, A.; Lallemand, B.; Cimmino, A.; Lamoral-Theys, D.; Gras, T.; Abou-Donia, A.; Dubois, J.; Lefranc, F.; Mathieu, V.; et al. Amaryllidaceae alkaloids belonging to different structural subgroups display activity against apoptosis-resistant cancer cells. J. Nat. Prod. 2010, 73, 1223–1227. [Google Scholar] [CrossRef]
- Cimmino, A.; Evidente, A.; Mathieu, V.; Andolfi, A.; Lefranc, F.; Kornienko, A.; Kiss, R. Phenazines and cancer. Nat. Prod. Rep. 2012, 29, 487–501. [Google Scholar] [CrossRef]
- Avolio, F.; Rimando, A.M.; Cimmino, A.; Andolfi, A.; Jain, S.; Tekwani, B.L.; Evidente, A. Inuloxins A-D and derivatives as antileishmanial agents: Structure-activity relationship study. J. Antibiot. 2014, 67, 597–601. [Google Scholar] [CrossRef]
- Evidente, A.; Kornienko, A.; Cimmino, A.; Andolfi, A.; Lefranc, F.; Mathieu, V.; Kiss, R. Fungal metabolites with anticancer activity. Nat. Prod. Rep. 2014, 31, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Artem, S.; Kireev, A.R.; Jenkins, A.R.; Romero, A.E.; Steelant, W.F.A.; Van Slambrouck, S.; Kornienko, A. Biological evaluation of structurally diverse Amaryllidaceae alkaloids and their synthetic derivatives: Discovery of novel leads for anticancer drug design. Planta Med. 2009, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and pharmacological activities of Amaryllidaceae alkaloids. RSC Adv. 2015. [Google Scholar] [CrossRef]
- Evidente, A.; Iasiello, I.; Randazzo, G. An improved method for the large-scale preparation of lycorine. Chem. Ind. 1984, 348–349. [Google Scholar]
- Cook, J.W.; Loudon, J.D. The alkaloids; Academic Press: New York, NY, USA, 1952; p. 331. [Google Scholar]
- Lamoral-Theys, D.; Andolfi, A.; van Goietsenoven, G.; Cimmino, A.; Le Calve, B.; Wauthoz, N.; Mégalizzi, V.; Gras, T.; Bruyère, C.; Dubois, J.; et al. Lycorine, the main phenanthridine Amaryllidaceae alkaloid, exhibits significant antitumor activity in cancer cells that display resistance to proapoptotic stimuli: An investigation of structure-activity relationship and mechanistic insight. J. Med. Chem. 2009, 52, 6244–6256. [Google Scholar] [CrossRef]
- Abou-Donia, A.H.; Abib, A.A.; El Din, A.S.; Evidente, A.; Gaber, M.; Scopa, A. Two betaine-type alkaloids from Egyptian Pancratium Marit. Phytochem. 1992, 31, 2139–2141. [Google Scholar] [CrossRef]
- Evidente, A.; Randazzo, G.; Surico, G.; Lavermicocca, P.; Arrigoni, O. Degradation of lycorine by Pseudomonas species strain ITM 311. J. Nat. Prod. 1985, 48, 564–570. [Google Scholar] [CrossRef]
- Kornienko, A.; Evidente, A. Chemistry, biology, and medicinal potential of narciclasine and its congeners. Chem. Rev. 2008, 108, 1982–2014. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Amaryllidaceae alkaloids: Absolute configuration and biological activity. Chirality 2017, 29, 486–499. [Google Scholar] [CrossRef]
- Masi, M.; Cala, A.; Tabanca, N.; Cimmino, A.; Green, I.R.; Bloomquist, J.R.; van Otterlo, W.A.L.; Macias, F.A.; Evidente, A. Alkaloids with activity against the Zika virus vector Aedes aegypti (L.)-Crinsarnine and sarniensinol, two new crinine and mesembrine type alkaloids isolated from the South African plant Nerine sarniensis. Molecules 2016, 21, 1432. [Google Scholar] [CrossRef]
- Masi, M.; van der Westhuyzen, A.E.; Tabanca, N.; Evidente, M.; Cimmino, A.; Green, I.R.; Bernier, U.R.; Becnel, J.J.; Bloomquist, J.R.; van Otterlo, W.A.L.; et al. Sarniensine, a mesembrine-type alkaloid isolated from Nerine sarniensis, an indigenous South African Amaryllidaceae, with larvicidal and adulticidal activities against Aedes aegypti. Fitoterapia 2017, 116, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Maddau, L.; Basso, S.; Linaldeddu, B.T.; Cimmino, A.; Scanu, B.; Deidda, A.; Tuzi, A.; Evidente, A. Diplopimarane, a 20-nor-ent-pimarane produced by the oak pathogen Diplodia quercivora. J. Nat. Prod. 2014, 77, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Cala, A.; Masi, M.; Cimmino, A.; Molinillo, J.M.G.; Macias, F.A.; Evidente, A. (+)-epi-Epoformin, a phytotoxic fungal cyclohexenepoxide: Structure activity relationships. Molecules 2018, 23, 1529. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Scanu, B.; Evidente, A.; Cimmino, A. Bioactive metabolites from pathogenic and endophytic fungi of forest trees. Curr. Med. Chem. 2018, 25, 208–252. [Google Scholar] [CrossRef]
- Brillinger, G.U.; Heberle, W.; Weber, B.; Achenbach, H. Metabolic products of microorganisms. 167. Cyclopaldic acid from Aspergillus duricaulis. 1 Production, isolation and biological properties. Arch. Microbiol. 1978, 116, 245–252. [Google Scholar] [CrossRef]
- Turner, W.B.; Aldridge, D.C. Polyketides. In The fungal metabolites II; Turner, W.B., Aldridge, D.C., Eds.; Academic Press: New York, NY, USA, 1983; p. 68. [Google Scholar]
- Graniti, A.; Sparapano, L.; Evidente, A. Cyclopaldic acid, a major phytotoxic metabolite of Seiridium cupressi, the pathogen of a canker disease of cypress. Plant Pathol. 1992, 41, 563–568. [Google Scholar] [CrossRef]
- Barilli, E.; Cimmino, A.; Masi, M.; Evidente, M.; Rubiales, D.; Evidente, A. Inhibition of early development stages of rust fungi by the two fungal metabolites cyclopaldic acid and epi-epoformin. Pest. Manag. Sci. 2017, 73, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- White, G.A.; Elliot, W.B. Inhibition of electron transport and oxidative phosphorylation in plant mitochondria by gladiolic acid and structurally related aromatic ortho-dialdehydes. Can. J. Biochem. Physiol. 1980, 58, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Guilbault, G.G.; Kramer, D.N. Fluorimetric determination of lipase, acylase, alpha- and gamma-chymotrypsin and inhibitors of these enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Ribeiro, K.A.L.; Monteiro de Carvalho, C.; Molina, M.T.; Pereira Lima, E.; López-Montero, E.; Reys, J.R.M.; Farias de Oliveira, M.B.; Ventura Pinto, A.; Santana, A.E.G.; Goulart, M.O.F. Activities of naphthoquinones against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae), vector of dengue and Biomphalaria glabrata (Say, 1818), intermediate host of Schistosoma mansoni. Acta Tropica. 2009, 111, 44–50. [Google Scholar] [CrossRef]
- Cimmino, A.; Mathieu, V.; Evidente, M.; Ferderin, M.; Banuls, L.M.Y.; Masi, M.; De Carvalho, A.; Kiss, R.; Evidente, A. Glanduliferins A and B, two new glucosylated steroids from Impatiens glandulifera, with in vitro growth inhibitory activity in human cancer cells. Fitoterapia 2016, 109, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cimmino, A.; Tabanca, N.; Becnel, J.J.; Bloomquist, J.R.; Evidente, A. A survey of bacterial, fungal and plant metabolites against Aedes aegypti (Diptera: Culicidae), the vector of yellow and dengue fevers and Zika virus. Open Chem. 2017, 15, 56–166. [Google Scholar] [CrossRef]
- Carrasco, L.; Fresno, M.; Vazquez, D. Narciclasine: An antitumor alkaloid which blocks peptide bond formation by eukaryotic ribosomes. FEBS Lett. 1975, 52, 236–239. [Google Scholar] [CrossRef]
- Jimenez, A.; Sanchez, L.; Vazquez, D. Location of resistance to the alkaloid narciclasine in the 60S ribosomal subunit. FEBS Lett. 1975, 55, 53–56. [Google Scholar] [CrossRef]
- Jimenez, A.; Santos, A.; Alonso, G.; Vasquez, D. Inhibitors of protein synthesis in eukariotic cells. Comparative effects of some Amaryllidaceae alkaloids. Biochim. Biophys. Acta. 1976, 425, 342–348. [Google Scholar] [CrossRef]
- Nair, J.J.; van Staden, J.; Bastida, J. Apoptosis-inducing effects of Amaryllidaceae alkaloids. Curr. Med. Chem. 2016, 23, 161–185. [Google Scholar] [CrossRef]
- Nair, J.J.; van Staden, J. Antiprotozoal alkaloid principles of the plant family Amaryllidaceae. Bioorg. Med. Chem. Lett. 2019, 29, 126642. [Google Scholar] [CrossRef]
- Han, H.; Xie, N.; Zhong, C.; Su, A.; Hui, X.; Zhang, X.; Jin, Z.; Li, Z.; Feng, J.; He, J. Aphicidal activities of Amaryllidaceae alkaloids from bulbs of Lycoris radiata against Aphis citricola. Ind. Crop Prod. 2018, 123, 372–378. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Avolio, F.; Ali, A.; Tabanca, N.; Khan, I.A.; Evidente, A. Cyclopaldic acid, seiridin, and sphaeropsidin A as fungal phytotoxins, and larvicidal and biting deterrents against Aedes aegypti (Diptera: Culicidae): Structure activity relationships. Chem. Biodivers. 2013, 10, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Evidente, M.; Masi, M.; Ali, A.; Tabanca, N.; Khan, I.A.; Evidente, A. Papyracillic acid and its derivatives as biting deterrents against Aedes aegypti (Diptera: Culicidae): Structure–activity relationships. Med. Chem. Res. 2015, 24, 3981–3989. [Google Scholar] [CrossRef]
- Tabanca, N.; Masi, M.; Epsky, N.D.; Nocera, P.; Cimmino, A.; Kendra, P.E.; Niogret, J.; Evidente, A. Laboratory evaluation of natural and synthetic aromatic compounds as potential attractants for male Mediterranean fruit Fly, Ceratitis capitata. Molecules 2019, 24, 2409. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Freda, F.; Santoro, E.; Superchi, S.; Evidente, A.; Cristofaro, M.; Masi, M. α-Costic acid, a plant sesquiterpene with acaricidal activity against Varroa destructor parasitizing the honey bee. Nat. Prod. Res. 2019, 1–8. [Google Scholar] [CrossRef]
- Rotundo, G.; Paventi, G.; Barberio, A.; De Cristofaro, A.; Notardonato, I.; Russo, M.V.; Germinara, G.S. Biological activity of Dittrichia viscosa (L.) Greuter extracts against adult Sitophilus granarius (L.) (Coleoptera, Curculionidae) and identification of active compounds. Sci. Rep. 2019, 9, 6429. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.C.; Harshan, V.; Saxena, A.; Sukumaran, P.; Sharma, M.C.; Lakshamana Kumar, M. Larvicidal and chemosterilant activity of Annona squamosa alkaloids against Anopheles stephensi. J. Am. Mosq. Control. Assoc. 1993, 9, 84–87. [Google Scholar]
- Sun, H.; Fu, X.; Chen, X.; Shi, W.P. Toxicity and influences of the alkaloids from Cynanchum mongolicum AL. Iljinski (Asclepiadaceae) on growth and cuticle components of Spodoptera litura Fabricius (Noctuidae) larvae. Nat. Prod. Res. 2012, 26, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Liu, P.; Yang, R.; Zhang, L.; Chen, H.; Camara, I.; Liu, Y.; Shi, W. Insecticidal constituents and activity of alkaloids from Cynanchum mongolicum. Molecules 2015, 20, 17483–17492. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Efficacy of naphthoquinones as insecticides against the house fly, Musca domestica L. Ind. Crops Prod. 2013, 43, 745–750. [Google Scholar] [CrossRef]
- Cespedes, C.L.; Lina-Garcia, L.; Kubo, I.; Salazar, J.R.; Ariza-Castolo, A.; Alarcon, J.; Aqueveque, P.; Werner, E.; Seigler, D.S. Calceolaria integrifolia s.l. complex, reduces feeding and growth of Acanthoscelides obtectus, and Epilachna varivestis. A new source of bioactive compounds against Dry Bean Pests. Ind. Crops Prod. 2016, 89, 257–267. [Google Scholar] [CrossRef]
- Sut, S.; Pavela, R.; Kolarčik, V.; Cappellacci, L.; Petrelli, R.; Maggi, F.; Dall’Acqua, S.; Benelli, G. Identification of Onosma visianii roots extract and purified shikonin derivatives as potential acaricidal agents against Tetranychus urticae. Molecules 2017, 22, 1002. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.G.; Lee, H.S. Insecticidal toxicities of naphthoquinone and its structural derivatives. Appl. Biol. Chem. 2016, 59, 3–8. [Google Scholar] [CrossRef]
- Puggioli, A.; Carrieri, M.; Dindo, M.L.; Medici, A.; Lees, R.S.; Gilles, J.R.L.; Bellini, R. Development of Aedes albopictus (Diptera: Culicidae) larvae under different laboratory conditions. J. Med. Entomol. 2017, 54, 142–149. [Google Scholar] [CrossRef]
- Evidente, A.; Cicala, M.R.; Randazzo, G.; Riccio, R.; Calabrese, G.; Liso, R.; Arrigoni, O. Lycorine structure-activity relationships. Phytochemistry 1983, 22, 2193–2196. [Google Scholar] [CrossRef]
- Karakoyun, Ç.; Masi, M.; Cimmino, A.; Önür, M.A.; Somer, N.U.; Kornienko, A.; Evidente, A. A brief up-to-date overview of Amaryllidaceae alkaloids: Phytochemical studies of Narcissus tazetta subsp. tazetta L., collected in Turkey. Nat. Prod. Commun. 2019, 14, 1–6. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for laboratory and field testing of mosquito larvicides; World Health Organization: Geneva, Switzerland, 2005; p. 33. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–266. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971; p. 333. [Google Scholar]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Benelli, G.; Conti, B. Cannabis sativa and Humulus lupulus essential oils as novel control tools against the invasive mosquito Aedes albopictus and fresh water snail Physella acuta. Ind. Crops Prod. 2016, 85, 318–323. [Google Scholar] [CrossRef]
- Mukhopadhyay, P. Multivariate Statistical Analysis; World Scientific: Singapore, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC50 (ppm) (LCL-HCL) 24 h | LC90 (ppm) (LCL-HCL) 24 h | χ2 (df) | LC50 (ppm) (LCL-HCL) 48 h | LC90 (ppm) (LCL-HCL) 48 h | χ2 (df) |
|---|---|---|---|---|---|
| Cyclopaldic acid | |||||
| 113.8 | 256.0 | 11.4 (28) | 40.1 | 105.1 | 30.6 (28) |
| (91.3–157.7) | (198.5–374.6) | (34.6–46.3) | (92.7–122.4) | ||
| 102.6 * | 202.5 * | 12.9 (28) * | 28.1 * | 60.3 * | 99.0 (28) * |
| (87.5–126.3) | (168.6–258.7) | (22.1–35.6) | (49.7–79.4) | ||
| N-Methyllycorine iodide | |||||
| 177.6 | 380.8 | 11.5 (23) | 78.5 | 176.2 | 23.1 (23) |
| (126.3–357.6) | (257.4–827.1) | (67.4–94.8) | (147.4–224.4) | ||
| 176.6 * | 332.5 * | 7.3 (23) * | 64.5 * | 128.8 * | 30.5 (23) * |
| (132.7–294.1) | (240.0–587.1) | (56.3–74.9) | (111.7–154.9) | ||
| Device® SC-15 | |||||
| 18.9 | 104.8 | 4.3 a (23) | |||
| (8.0–26.3) | (88.2–132.1) | ||||
| 8.8 * | 59.3 * | 12.2 a (23) * | |||
| (1.2–14.3) | (50.8–72.6) | ||||
| Compound Concentration (ppm) | Larval Mortality (%) 2 | Mean Larval Duration (Days) ± SE | Pupal Mortality (%) 3 | Mean Pupal Duration (Days) ± SE |
|---|---|---|---|---|
| Control (distilled water) | ||||
| 7.0 (7) | 6.4 ± 3.5 a | 2.1 (4) | 3.5 ± 2.3 a | |
| Control (DMSO 1%) | ||||
| 7.0 (7) | 6.6 ± 3.0 a | 4.3 (4) | 3.8 ± 1.9 a | |
| Cyclopaldic acid | ||||
| 100 | 100.0 (3) | |||
| 50 | 100.0 (5) | |||
| 25 | 79.0 (11) | 8.6 ± 1.4 b | 38.1 (5) | 3.1 ± 2.5 b |
| 12.5 | 38.0 (28) | 8.5 ± 2.5 b | 16.1 (5) | 3.6 ± 1.0 b |
| 6.25 | 21.0 (16) | 7.6 ± 2.5 b | 15.2 (9) | 3.6 ± 1.2 ab |
| 3.125 | 13.0 (8) | 7.3 ± 2.6 b | 11.5 (7) | 3.0 ± 1.2 ab |
| 1,2-O,O’-Diacetyllycorine | ||||
| 50 | 98.0 (14) | |||
| 25 | 62.0 (7) | 7.3 ± 2.0 b | 7.9 (7) | 3.3 ± 0.9 b |
| 12.5 | 44.0 (12) | 7.1 ± 2.3 b | 7.1 (5) | 2.9 ± 0.8 b |
| 6.25 | 34.0 (7) | 7.1 ± 2.5 b | 12.3 (7) | 4.1 ± 1.3 b |
| N-methyllycorine iodide | ||||
| 100 | 100.00 (10) | |||
| 50 | 75.0 (14) | 7.2 ± 1.4 b | 48.0 (6) | 4.5 ± 1.1 cb |
| 25 | 25.0 (25) | 7.4 ± 2.0 b | 12.0 (10) | 4.8 ± 1.2 a |
| 12.5 | 21.0 (22) | 7.3 ± 1.7 b | 12.7 (7) | 3.9 ± 1.3 ab |
| 6.25 | 16.0 (17) | 7.2 ± 1.8 a | 13.1 (6) | 4.4 ± 1.2 ab |
| α-Dihydrolycorine | ||||
| 100 | 100.0 (9) | 0.00 | ||
| 50 | 100.0 (9) | 0.00 | ||
| 25 | 12.0 (15) | 6.9 ± 3.1 a | 5.7 (7) | 4.3 ± 1.7 a |
| 12.5 | 13.0 (7) | 7.2 ± 3.6 a | 5.7 (5) | 4.1 ± 1.1 a |
| 6.25 | 10.0 (5) | 6.9 ± 2.7 a | 4.4 (3) | 3.9 ± 1.3 a |
| Device® SC-15 | ||||
| 100 | 100.0 (1) | |||
| 50 | 100.0 (2) | |||
| 25 | 100.0 (2) | |||
| 12.5 | 100.0 (2) | |||
| 7 | 100.0 (2) | |||
| LC50 (ppm) (LCL-HCL) 24 h | LC90 (ppm) (LCL-HCL) 24 h | χ2 (df) | LC50 (ppm) (LCL-HCL) 48 h | LC90 (ppm) (LCL-HCL) 48 h | χ2 (df) | LC50 (ppm) (LCL-HCL) 72 h | LC90 (ppm) (LCL-HCL) 72 h | χ2 (df) |
|---|---|---|---|---|---|---|---|---|
| 2-Methyl-1,4-naphthoquinone | ||||||||
| 24.2 | 39.4 | 35.8 a (23) | 14.5 | 20.7 | 11.6 (23) | 12.1 | 15.8 | 10.2 (23) |
| (21.6–27.1) | (35.3–45.4) | (13.5–15.6) | (19.1–22.9) | (11.5–12.9) | (14.7–17.7) | |||
| 2-Hydroxy-1,4-naphthoquinone | ||||||||
| 33.7 | 41.0 | 1.8 (23) | 31.9 | 39.2 | 0.8 (23) | 30.9 | 39.6 | 13.9 (23) |
| (31.2–38.0) | (37.0–48.5) | (29.3–38.4) | (34.5–52.0) | (28.8–34.1) | (35.9–46.1) | |||
| Device® SC-15 | ||||||||
| 50.6 | 95.3 | 60.2 a (23) | 36.4 | 90.6 | 43.4 a (23) | 15.0 | 58.9 | 26.2 (23) |
| (42.7–60.1) | (81.9–116.5) | (28.8–44.4) | (77.0–112.2) | (9.7–19.5) | (51.3–70.2) | |||
| CompoundConcentration (ppm) | GLM (Time × Treatment) a | Bonferroni Test b Mean Treatment | Mean Control | Statistical Significance | |
|---|---|---|---|---|---|
| 2-Methyl-1,4-naphthoquinone | |||||
| 6.25 | F2.16 = 2.7 | p > 0.05 | 19.6 ± 0.2 | 20.0 ± 0.2 | n.s. |
| 12.5 | F2.16 = 55.6 | p < 0.01 | 12.5 ± 0.1 | 20.0 ± 0.1 | ** |
| 25 | F2.16 = 297.8 | p < 0.01 | 2.2 ± 0.1 | 20.0 ± 0.1 | ** |
| 50 | F2.16 = 36.0 | p < 0.01 | 0.4 ± 0.05 | 20.0 ± 0.05 | ** |
| 100 | 0.0 ± 0.0 | 20.0 ± 0.0 | ** | ||
| 2-Hydroxy-1,4-naphthoquinone | |||||
| 6.25 | F2.16 = 1.0 | p > 0.05 | 19.9 ± 0.05 | 20.0 ± 0.05 | n.s. |
| 12.5 | F2.16 = 2.7 | p > 0.05 | 19.9 ± 0.06 | 20.0 ± 0.06 | n.s. |
| 25 | F2.16 = 33.8 | p < 0.01 | 17.5 ± 0.2 | 20.0 ± 0.2 | ** |
| 50 | 0.0 ± 0.0 | 20.0 ± 0.0 | ** | ||
| 100 | 0.0 ± 0.0 | 20.0 ± 0.0 | ** | ||
| 2-Methoxy-1,4-naphthoquinone | |||||
| 6.25 | - | - | 19.8 ± 0.1 | 20.0 ± 0.1 | n.s. |
| 12.5 | F2.16 = 1.0 | p > 0.05 | 19.9 ± 0.09 | 20.0 ± 0.09 | n.s. |
| 25 | F2.16 = 1.0 | p > 0.05 | 19.7 ± 0.2 | 20.0 ± 0.2 | n.s. |
| 50 | F2.16 = 28.2 | p < 0.01 | 16.7 ± 0.6 | 20.0 ± 0.6 | ** |
| 100 | 0.0 ± 0.00 | 20.0 ± 0.0 | ** | ||
| Device® SC-15 | |||||
| 7 | F2.16 = 351.4 | p < 0.01 | 16.0 ± 0.3 | 20.0 ± 0.3 | ** |
| 12.5 | F2.16 = 77.5 | p < 0.01 | 14.1 ± 0.3 | 20.0 ± 0.3 | ** |
| 25 | F2.16 = 51.1 | p < 0.01 | 12.2 ± 0.3 | 20.0 ± 0.3 | ** |
| 50 | F2.16 = 10.7 | p < 0.01 | 3.5 ± 0.5 | 20.0 ± 0.5 | ** |
| 100 | F2.16 = 12.3 | p < 0.01 | 2.1 ± 0.3 | 20.0 ± 0.3 | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganassi, S.; Masi, M.; Grazioso, P.; Evidente, A.; De Cristofaro, A. Activity of Some Plant and Fungal Metabolites towards Aedes albopictus (Diptera, Culicidae). Toxins 2021, 13, 285. https://doi.org/10.3390/toxins13040285
Ganassi S, Masi M, Grazioso P, Evidente A, De Cristofaro A. Activity of Some Plant and Fungal Metabolites towards Aedes albopictus (Diptera, Culicidae). Toxins. 2021; 13(4):285. https://doi.org/10.3390/toxins13040285
Chicago/Turabian StyleGanassi, Sonia, Marco Masi, Pasqualina Grazioso, Antonio Evidente, and Antonio De Cristofaro. 2021. "Activity of Some Plant and Fungal Metabolites towards Aedes albopictus (Diptera, Culicidae)" Toxins 13, no. 4: 285. https://doi.org/10.3390/toxins13040285
APA StyleGanassi, S., Masi, M., Grazioso, P., Evidente, A., & De Cristofaro, A. (2021). Activity of Some Plant and Fungal Metabolites towards Aedes albopictus (Diptera, Culicidae). Toxins, 13(4), 285. https://doi.org/10.3390/toxins13040285