Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy


{kind=link}
Abstract
:1. Introduction
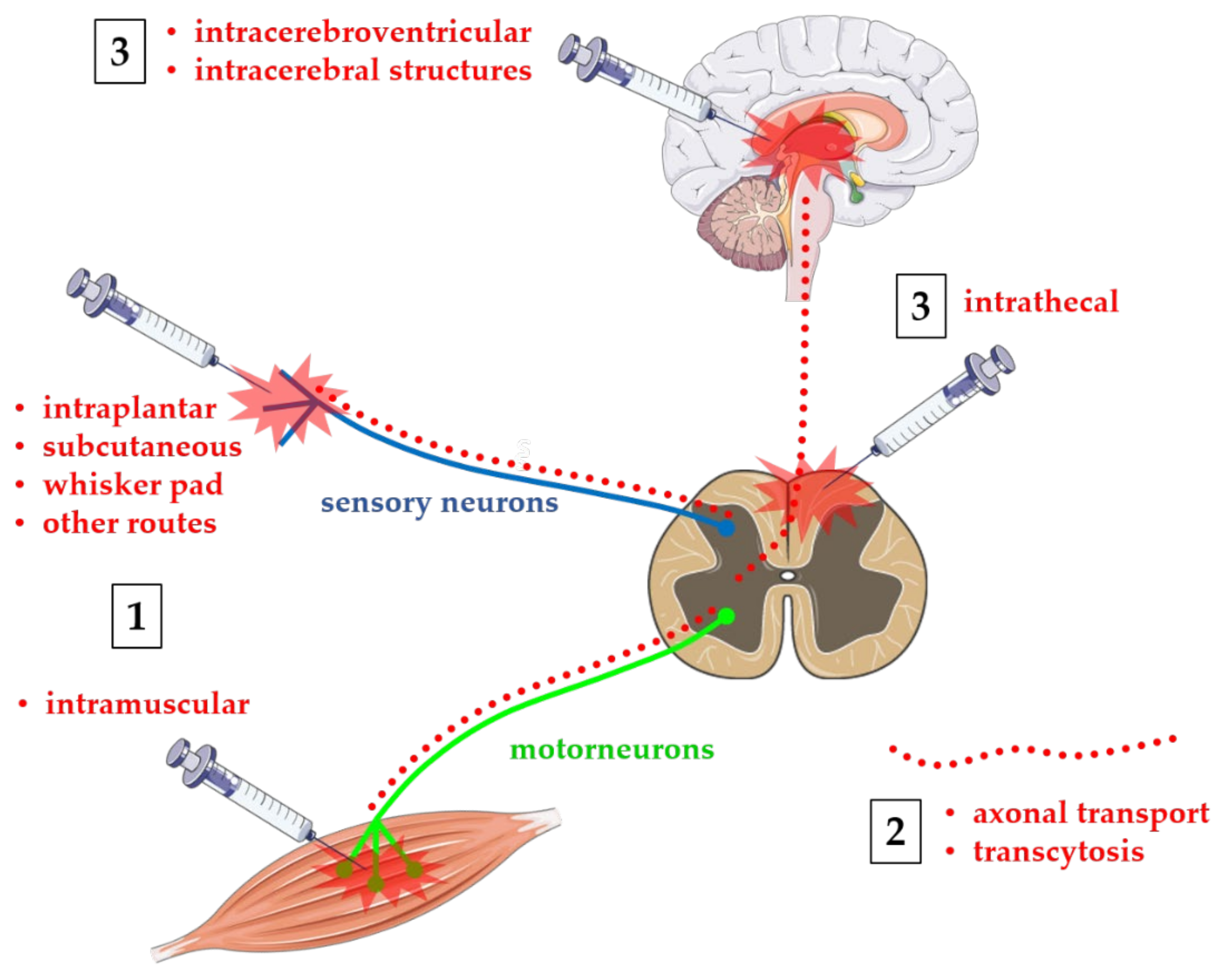
2. Indirect Central Effects following Peripheral Injection of BoNTs
3. Axonal Transport after Peripheral Injection of BoNTs
4. Central Effects of BoNTs after Direct Injection on CNS
5. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Montecucco, C.; Rasotto, M.B. On botulinum neurotoxin variability. mBio 2015, 6, e02131. [Google Scholar] [CrossRef] [Green Version]
- Tehran, D.A.; Pirazzini, M. Novel Botulinum Neurotoxins: Exploring Underneath the Iceberg Tip. Toxins 2018, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Masuyer, G.; Stenmark, P. Botulinum and Tetanus Neurotoxins. Annu. Rev. Biochem. 2019, 20, 811–837. [Google Scholar] [CrossRef]
- Steward, L.; Brin, M.F.; Brideau-Andersen, A. Novel Native and Engineered Botulinum Neurotoxins. Handb. Exp. Pharmacol. 2021, 263, 63–89. [Google Scholar]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum Neurotoxins: Biology, Pharmacology, and Toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef]
- Kumar, R.; Dhaliwal, H.P.; Kukreja, R.V.; Singh, B.R. The Botuliunum Toxin as a Therapeutic Agent: Molecular Structure and Mechanism of Action in Motor and Sensory Systems. Semin. Neurol. 2016, 36, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safarpour, Y.; Jabbari, B. Botulinum Toxin Treatment of Movement Disorders. Curr. Treat. Options Neurol. 2018, 20, 4. [Google Scholar] [CrossRef] [PubMed]
- Cocco, A.; Albanese, A. Recent developments in clinical trials of botulinum neurotoxins. Toxicon 2018, 147, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Fonfria, E.; Maignel, J.; Lezmi, S.; Martin, V.; Splevins, A.; Shubber, S.; Kalinichev, M.; Foster, K.; Picaut, P.; Krupp, J. The Expanding Therapeutic Utility of Botulinum Neurotoxins. Toxins 2018, 10, 208. [Google Scholar] [CrossRef] [Green Version]
- Chiu, S.Y.; Burns, M.R.; Malaty, I.A. An Update on Botulinum Toxin in Neurology. Neurol. Clin. 2021, 39, 209–229. [Google Scholar] [CrossRef]
- Spiegel, L.L.; Ostrem, J.L.; Bledsoe, I.O. FDA Approvals and Consensus Guidelines for Botulinum Toxins in the Treatment of Dystonia. Toxins 2020, 12, 332. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chiang, B.J.; Liao, C.H. Mechanism of Action of Botulinum Toxin A in Treatment of Functional Urological Disorders. Toxins 2020, 12, 129. [Google Scholar] [CrossRef] [Green Version]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Luvisetto, S.; Gazerani, P.; Cianchetti, C.; Pavone, F. Botulinum Toxin Type a as a Therapeutic Agent against Headache and Related Disorders. Toxins 2015, 7, 3818–3844. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.C.; Hutton, E.J.; Matharu, M. Onabotulinumtoxin A in Migraine: A Review of the Literature and Factors Associated with Efficacy. J. Clin. Med. 2021, 10, 2898. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Hulsopple, C.; Boyce, B. Utilization of Botulinum Toxin for Musculoskeletal Disorders. Curr. Sports Med. Rep. 2020, 19, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Egeo, G.; Fofi, L.; Barbanti, P. Botulinum Neurotoxin for the Treatment of Neuropathic Pain. Front. Neurol. 2020, 11, 716. [Google Scholar] [CrossRef]
- Giladi, N. The mechanism of action of Botulinum toxin type A in focal dystonia is most probably through its dual effect on efferent (motor) and afferent pathways at the injected site. J. Neurol. Sci. 1997, 152, 132–135. [Google Scholar] [CrossRef]
- Currà, A.; Trompetto, C.; Abbruzzese, G.; Berardelli, A. Central Effects of Botulinum Toxin Type A: Evidence and Supposition. Mov. Disord. 2004, 19, 560–564. [Google Scholar] [CrossRef]
- Caleo, M.; Antonucci, F.; Restani, L.; Mazzocchio, R. A reappraisal of the central effects of botuluinum neurotoxin type A: By what mechanism? J. Neurochem. 2009, 109, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Caleo, M.; Restani, L. Exploiting Botulinum Neurotoxins for the Study of Brain Physiology and Pathology. Toxins 2018, 10, 175. [Google Scholar] [CrossRef] [Green Version]
- Caleo, M.; Restani, L. Direct central nervous system effects of botulinum neuroroxin. Toxicon 2018, 147, 68–72. [Google Scholar] [CrossRef]
- Luvisetto, S. Botulinum Toxin and Neuronal Regeneration after Traumatic Injury of Central and Peripheral Nervous System. Toxins 2020, 12, 434. [Google Scholar] [CrossRef]
- Filippi, G.M.; Errico, P.; Santarelli, R.; Bagolini, B.; Manni, E. Botulinum A toxin effects on rat jaw muscle spindles. Acta Otolaryngol. 1993, 113, 400–404. [Google Scholar] [CrossRef]
- Rosales, R.L.; Arimura, K.; Takenaga, M.; Osame, M. Extrafusal and intrafusal muscle effects in experimental botulinum toxin-A injection. Muscle Nerve 1996, 19, 488–496. [Google Scholar] [CrossRef]
- Mazzocchio, R.; Caleo, M. More than at the Neuromuscular Synapse: Actions of Botulinum Neurotoxin A in the Central Nervous System. Neuroscientist 2015, 21, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Mas, M.F.; Li, S.; Francisco, G.E. Centrally mediated late motor recovery after botulinum toxin injection: Case reports and a review of current evidence. J. Rehabil. Med. 2017, 49, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Abbruzzese, G.; Berardelli, A. Neurophysiological effects of botulinum toxin type A. Neurotox. Res. 2006, 9, 109–114. [Google Scholar] [CrossRef]
- Palomar, F.J.; Mir, P. Neurophysiologicasl changes after intramuscular injection of botulinum toxin. Clin. Neurophysiol. 2012, 123, 54–60. [Google Scholar] [CrossRef]
- Toma, K.; Nakai, T. Functional MRI in human motor control studies and clinical applications. Magn. Reason. Med. Sci. 2002, 1, 109–120. [Google Scholar] [CrossRef]
- Blood, A.J. Imaging Studies in Focal Dystonias: A Systems Level Approach to Studying a Systems Level Disorder. Curr. Neuropharmacol. 2013, 11, 3–15. [Google Scholar]
- Hok, P.; Veverka, T.; Hlustik, P.; Nevrly, M.; Kanovsky, P. The Central Effects of Botulinum Toxin in Dystonia and Spasticity. Toxins 2021, 13, 155. [Google Scholar] [CrossRef] [PubMed]
- Senkarova, Z.; Hlustik, P.; Otruba, P.; Herzig, R.; Kanovsky, P. Modulation of Cortical Activity in Patients Suffering for Upper Arm Spasticity following Stroke and Treated with Botulinum Toxin A: An fMRI Study. J. Neuroimaging 2010, 20, 9–15. [Google Scholar] [CrossRef]
- Manganotti, P.; Acler, M.; Formaggio, E.; Avesani, M.; Milanese, F.; Baraldo, A.; Storti, S.F.; Gasparini, A.; Cerini, R.; Pozzi Mucelli, R.; et al. Changes in cerebral activity after decreased upper-limb hypertonus: An EMG-fMRI study. Magn. Res. Imaging 2010, 28, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Diserens, K.; Ruegg, D.; Kleiser, R.; Hyde, S.; Perret, N.; Vuadens, P.; Fornari, E.; Vingerhoets, F.; Seitz, R.J. Effectr od Repetitive Arm Cycling following Botulinum Toxin Injection for Poststroke Spasticity: Evidence from fMRI. Neurorehabil. Neural Repair 2010, 24, 753–762. [Google Scholar] [CrossRef]
- Veverka, T.; Hlustik, P.; Tomasova, Z.; Hok, P.; Otruba, P.; Kral, M.; Tudos, Z.; Zapletalova, J.; Herzig, R.; Krobot, A.; et al. BoNT-A related changes of cortical activity in patients suffering from severe hand paralysis with arm spasticity following ischemic stroke. J. Neurol. Sci. 2012, 319, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Veverka, T.; Hulstik, P.; Hok, P.; Otruba, P.; Tudos, Z.; Zapletalova, J.; Krobot, A.; Kanovsky, P. Cortical activity modulation by botulinum toxin type A in patients with post-stroke arm spasticity: Real and imagined hand movement. J. Neurol. Sci. 2014, 346, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Tomasova, Z.; Hlustik, P.; Kral, M.; Otruba, M.; Herzig, R.; Krobot, A.; Kanovsky, P. Cortical Activation Changes in Patients Suffering from Post-Stroke Arm Spasticity and Treated with Botulinum Toxin A. J. Neuroimaging 2013, 23, 337–344. [Google Scholar] [CrossRef]
- Bergfeldt, U.; Jonsson, T.; Bergfeldt, L.; Julin, P. Cortical activation changes and improved motor functions after focal spasticity therapy—An interventional study applying repeated fMRI. BMC Neurol. 2015, 15, 52. [Google Scholar] [CrossRef] [Green Version]
- Opavsky, R.; Hlustik, P.; Otruba, P.; Kanovsky, P. Sensorimotor network in cervical dystonia and the effect of botulinum toxin treatment: A functional MRI study. J. Neurol. Sci. 2011, 306, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Dresel, C.; Bayer, F.; Castrop, F.; Rimpau, C.; Zimmer, C.; Haslinger, B. Botulinum toxin modulates basl ganglia but not deficient somatosensory activation in orofacial dystonia. Mov. Disord. 2011, 26, 1496–1502. [Google Scholar] [CrossRef] [PubMed]
- Delnooz, C.C.S.; Pasman, J.W.; Beckmann, C.F.; van de Warrenburg, B.P.C. Task-Free Functional MRI in Cervical Dystonia Reveals Multi-Network Changes That Partially Normalize with Botulinum Toxin. PLoS ONE 2013, 8, e62877. [Google Scholar] [CrossRef] [Green Version]
- Delnooz, C.C.S.; Pasman, J.W.; Beckmann, C.F.; van de Warrenburg, B.P.C. Altered striatal and pallidal connectivity in cervical dystonia. Brain Struct. Funct. 2015, 220, 513–523. [Google Scholar] [CrossRef]
- Brodoehl, S.; Wagner, F.; Prell, T.; Klingner, C.; Witte, O.W.; Gunther, A. Cause or effect: Altered brain network activity in cervical dystonia is partially normalized by botulinum toxin treatment. Neuroimage Clin. 2019, 22, 101792. [Google Scholar] [CrossRef] [PubMed]
- Nevrly, M.; Hlustik, P.; Hok, P.; Otruba, P.; Tudos, Z.; Kanovsky, P. Changes in sensorimotor network activation after botulinum toxin type A injections in patients with cervical dystonia: A functional MRI study. Exp. Brain Res. 2018, 236, 2627–2637. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-L.; Weber, D.J.; Munin, M.C. Changes in Cerebellar Activation After Onabotulinumtoxin A Injections for Spasticity After Chronic Stroke: A pilot Functional Magnetic Resonance Imaging Study. Arch. Phys. Med. Rehabil. 2015, 96, 2007–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hok, P.; Hvizdosova, L.; Otruba, P.; Kaiserova, M.; Trneckova, M.; Tudos, Z.; Hlustik, P.; Kanovsky, P.; Nevrly, M. Botulinum toxin injection changes resting state cerebellar connectivity in cervical dystonia. Sci. Rep. 2021, 11, 8322. [Google Scholar] [CrossRef]
- Li, G.-F.; Ban, S.; Wang, M.; Zhang, J.; Lu, H.; Shi, Y.-H.; He, X.-W.; Wu, Y.-L.; Peng, P.; Liu, Y.-S.; et al. Brain functional changes in patients with botulism after illegal cosmetic injections of botulinum toxin: A resting-state fMRI study. PLoS ONE 2018, 11, e0207448. [Google Scholar] [CrossRef] [Green Version]
- Weise, D.; Weise, C.M.; Naumann, M. Central Effects of Botulinum Neurotoxin—Evidence from Human Studies. Toxins 2019, 11, 21. [Google Scholar] [CrossRef] [Green Version]
- Yesudhas, A.; Roshan, S.A.; Radhakrishnan, R.K.; Abirami, G.P.P.; Manickam, N.; Selvaraj, K.; Elumalai, G.; Shanmugaapriya, S.; Anusuyadevi, M.; Kandasamy, M. Intramuscular injection of BOTOX® boosts learning and memory in adult mice in association with enriched circulation of platelets and enhanced density of pyramidal neurons in the Hippocampus. Neurochem. Res. 2020, 45, 2856–2867. [Google Scholar] [CrossRef]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-distance retrograde effects of botulinum neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef]
- Restani, L.; Antonucci, F.; Gianfranceschi, L.; Rossi, C.; Rossetto, O.; Caleo, M. Evidence for anterograde transport and transcytosis of botulinum neurotoxin A (BoNT/A). J. Neurosci. 2011, 31, 15650–15659. [Google Scholar] [CrossRef] [PubMed]
- Restani, L.; Novelli, E.; Bottari, D.; Leone, P.; Barone, I.; Galli-Resta, L.; Strettori, E.; Caleo, M. Botulinum neurotoxin A impairs neurotransmission following retrograde transynaptic transport. Traffic 2012, 13, 1083–1089. [Google Scholar] [CrossRef] [Green Version]
- Restani, L.; Giribaldi, F.; Manich, M.; Bercsenyi, K.; Menendez, G.; Rossetto, O.; Caleo, M.; Schiavo, G. Botulinum Neurotoxins A and E Undergo Retrograde Axonal Transport in Primary Motor Neurons. PLoS Pathog. 2012, 8, e1003087. [Google Scholar] [CrossRef] [Green Version]
- Caleo, M.; Spinelli, M.; Colosimo, F.; Matak, I.; Rossetto, O.; Lackovic, Z.; Restani, L. Transynaptic Action of Botulinum Neurotoxin Type A at Central Cholinergic Boutons. J. Neurosci. 2018, 38, 10329–10337. [Google Scholar] [CrossRef]
- Bomba-Warczak, E.; Vevea, J.D.; Brittain, J.M.; Figueroa-Bernier, A.; Tepp, W.H.; Johnson, E.A.; Yeh, F.L.; Chapman, E.R. Interneuronal Transfer and Distal Action of Tetanus Toxin and Botulinum Neurotoxins A and D in Central Neurons. Cell Rep. 2016, 16, 1974–1987. [Google Scholar] [CrossRef] [Green Version]
- Papagioannopoulou, D.; Vardouli, L.; Dimitriadis, F.; Apostolidis, A. Retrograde transport of radiolabelled botulinum neurotoxin type A to the CNS after intradetrusor inejection in rats. BJU Int. 2016, 117, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, S.; Vacca, V.; Ricordy, R.; Uggenti, C.; Tata, A.M.; Luvisetto, S.; Pavone, F. The analgesic effect on neuropathic pain of retrogradely transported botulinum neurotoxin A involves Schwann cells and astrocytes. PLoS ONE 2012, 7, e47977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeftinija, S.D.; Jeftinija, K.V.; Stefanovic, G. Cultured astrocytes express proteins involved in vesicular glutamate release. Brain Res. 1997, 750, 41–47. [Google Scholar] [CrossRef]
- Koizumi, H.; Goto, S.; Okita, S.; Morigaki, R.; Akaike, N.; Torii, Y.; Harakawa, T.; Ginnaga, A.; Kaji, R. Spinal Central Effects of Peripherally Applied Botulinum Neurotoxin A in Comparison between Its Subtypes A1 and A2. Front. Neurol. 2014, 5, 98. [Google Scholar] [CrossRef] [Green Version]
- Bach-Rojecky, L.; Lackovic, Z. Central origin of the antinociceptive action of botulinum toxin type A. Pharmacol. Biochem. Behav. 2009, 94, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Bach-Rojecky, L.; Salkovic-Petrisic, M.; Lackovic, Z. Botulinum toxin type A reduces pain supersensitivity in experimental diabetic neuropathy: Bilateral effect after unilateral injection. Eur. J. Pharmacol. 2010, 633, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matak, I.; Bach-Rojecky, L.; Filipovic, B.; Lackovic, Z. Behavioral and immunohistochemical evidence for central antinociceptive activity of botulinum toxin A. Neuroscience 2011, 186, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipovic, B.; Matak, I.; Bach-Rojecky, L.; Lackovic, Z. Central action of peripherally applied botulinum toxin type A on pain and dural protein extravasation in rat model of trigeminal neuropathy. PLoS ONE 2012, 7, e29803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matak, I.; Riederer, P.; Lacković, Z. Botulinum toxin’s axonal transport from periphery to the spinal cord. Neurochem. Int. 2012, 61, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Lackovic, Z.; Filipovic, B.; Matak, I.; Helyes, Z. Activity of botulinum toxin type A in cranial dura: Implications for treatment of migraine and other headaches. Br. J. Pharmacol. 2016, 173, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Drinovac Vlah, V.; Filipovic, B.; Bach-Rojecky, L.; Lackovic, Z. Role of central versus peripheral opioid system in antinociceptive and anti-inflammatory effect of botulinum toxin type A in trigeminal region. Eur. J. Pain 2018, 22, 583–591. [Google Scholar] [CrossRef]
- Jang, S.-H.; Park, S.-J.; Lee, C.-J.; Ahn, D.-K.; Han, S.-K. Botulinum toxin type A enhances the inhibitory spontaneous postsynaptic currents on the substantia gelatinosa neurons of the subnucleus caudalis in immature mice. Korean J. Physiol. Pharmacol. 2018, 22, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Ni, L.-H.; Cao, S.-X.; Lian, H.; Xing-Yue Hu, X.-Y. Unilateral whisker pad injection of botulinum toxin type a enhances spatial learning in mice. Neuroreport 2018, 29, 987–992. [Google Scholar] [CrossRef]
- Marino, M.J.; Terashima, T.; Steinauer, J.J.; Eddinger, K.A.; Yaksh, T.L.; Xu, Q. Botulinum toxin B in the sensory afferent: Transmitter release, spinal activation, and pain behavior. Pain 2014, 155, 674–684. [Google Scholar] [CrossRef] [Green Version]
- Marchand-Pauvert, V.; Aymard, C.; Giboin, L.-S.; Dominici, F.; Rossi, A.; Mazzocchio, R. Beyond muscular effects: Depression of spinal recurrent inhibition after botulinum neurotoxin A. J. Physiol. 2013, 593, 1017–1029. [Google Scholar] [CrossRef]
- Aymard, C.; Giboin, L.-S.; Lackmy-Vallée, A.; Marchand-Pauvert, V. Spinal plasticity in stroke patients after botulinum neurotoxin A injection in ankle plantar flexors. Physiol. Rep. 2013, 1, e00173. [Google Scholar] [CrossRef] [PubMed]
- Schmieg, N.; Menendez, G.; Schiavo, G.; Terenzio, M. Signalling endosomes in axonal transport: Travel updates on the molecular highway. Semin. Cell Dev. Biol. 2014, 27, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Surana, S.; Tosolini, A.P.; Meyer, I.F.G.; Fellows, A.D.; Novoselov, S.S.; Schiavo, G. The travel diaries of tetanus and botulinum neurotoxins. Toxicon 2018, 147, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Martin, S.; Papadopulus, A.; Harper, C.B.; Maylyutov, T.A.; Niranjan, D.; Glass, N.R.; Cooper-White, J.J.; Sibarita, J.B.; Choquet, D.; et al. Control of atuophagosome axonal retrograde flux by presynaptic activity unveiled using botulinum neurotoxin type a. J. Neurosci. 2015, 35, 6179–6194. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, C.B.; Howard, B.D. The effects of botulinum toxin on acethylcholine metabolism in mouse brain slices and synaptosomes. J. Neurochem. 1978, 31, 1005–1013. [Google Scholar] [CrossRef]
- Hirokawa, N.; Kitamura, M. Binding of Clostridium botulinum neurotoxin to the presynaptic membrane in the central nervous system. J. Cell Biol. 1979, 81, 43–49. [Google Scholar] [CrossRef]
- Bigalke, H.; Heller, I.; Bizzini, B.; Haberman, E. Tetanus and botulinum A toxin inhibit release and uptake of various transmitter, as studied with particulate preparations from rat brain and spinal cord. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1981, 316, 244–251. [Google Scholar] [CrossRef]
- Haberman, E.; Muller, H.; Tudel, M. Tetanus toxin and botulinum A and C neurotoxins inhibit noradrenaline release from cultured mouse brain. J. Neurochem. 1988, 51, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Prieto, J.; Sihra, T.S.; Evans, D.; Ashton, A.; Dolly, J.O.; Nicholls, D.G. Botulinum toxin A blocks glutamate exocytosis from guinea-pig cerebral cortical synaptosomes. Eur. J. Biochem. 1987, 165, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Black, J.D.; Dolly, J.O. Selective location of acceptors for botulinum neurotoxin A in the central and peripheral nervous systems. Neuroscience 1987, 23, 767–779. [Google Scholar] [CrossRef]
- Rabasseda, X.; Blasi, J.; Marsal, J.; Dunant, Y.; Casanova, A.; Bizzini, B. Tetanus and botulinum toxins block the release of acetylcholine from slices of rat striatum and from the isolated electric organ of Torpedo at different concentrations. Toxicon 1988, 26, 329–336. [Google Scholar] [CrossRef]
- Williamson, L.C.; Halpern, J.L.; Montecucco, C.; Brown, J.E.; Neale, E.A. Clostridial neurotoxins and substrate proteolysis in intact neurons: Botulinum neurotoxin C acts on synaptosomal-associated protein of 25 kDa. J. Biol. Chem. 1996, 271, 7694–7699. [Google Scholar] [CrossRef] [Green Version]
- Luvisetto, S.; Rossetto, O.; Montecucco, C.; Pavone, F. Toxicity of botulinum neurotoxins in central nervous system of mice. Toxicon 2003, 41, 475–481. [Google Scholar] [CrossRef]
- Luvisetto, S.; Marinelli, S.; Rossetto, O.; Montecucco, C.; Pavone, F. Central injection of botulinum neurotoxins: Behavioural effects in mice. Behav. Pharmacol. 2004, 15, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Lackovic, Z.; Revic, V.; Riederer, P.F. Single intracerebroventricular injection of botulinum toxin type A produces slow onset and long-term memory impairment in rats. J. Neural Transm. 2009, 16, 1273–1280. [Google Scholar] [CrossRef]
- Luvisetto, S.; Marinelli, S.; Lucchetti, F.; Marchi, F.; Cobianchi, S.; Rossetto, O.; Montecucco, C.; Pavone, F. Botulinum neurotoxins and formalin-induced pain: Central vs. peripheral effects in mice. Brain Res. 2006, 1082, 124–131. [Google Scholar] [CrossRef]
- Hawlitschka, A.; Antipova, V.; Schmitt, O.; Witt, M.; Benecke, R.; Mix, E.; Wree, A. Intracerebrally applied botulinum neurotoxin in experimental neuroscience. Curr. Pharm. Biotechnol. 2013, 14, 124–130. [Google Scholar]
- Ando, S.; Kobayashi, S.; Waki, H.; Kon, K.; Fukui, F.; Tadenuma, T.; Iwamoto, M.; Takeda, Y.; Izumiyama, N.; Watanabe, K.; et al. Animal model of dementia induced by entorhinal synaptic damage and partial restoration of cognitive deficits by BDNF and carnitine. J. Neurosci. Res. 2002, 70, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Costantin, L.; Bozzi, Y.; Richichi, C.; Viegi, A.; Antonucci, F.; Funicello, M.; Gobbi, M.; Mennini, T.; Rossetto, O.; Montecucco, C.; et al. Antiepileptic effects of botulinum neurotoxin E. J. Neurosci. 2005, 25, 1943–1951. [Google Scholar] [CrossRef] [Green Version]
- Bozzi, Y.; Costantin, L.; Antonucci, F.; Caleo, M. Action of botulinum neurotoxins in the central nervous system: Antiepileptic effects. Neurotox. Res. 2006, 9, 197–203. [Google Scholar] [CrossRef]
- Manno, I.; Antonucci, F.; Caleo, M.; Bozzi, Y. BoNT/E prevents seizure-induced activation of caspase 3 in the rat hippocampus. Neuroreport 2007, 18, 577–580. [Google Scholar] [CrossRef]
- Antonucci, F.; Di Garbo, A.; Novelli, E.; Manno, I.; Sartucci, F.; Bozzi, Y.; Caleo, M. Botulinum neurotoxin E (BoNT/E) reduces CA1 neuron loss and granule cell dispersion, with no effects on chronic seizures, in a mouse model of temporal lobe epilepsy. Exp. Neurol. 2008, 210, 388–401. [Google Scholar] [CrossRef]
- Antonucci, F.; Bozzi, Y.; Caleo, M. Intrahippocampal infusion of botulinum neurotoxin E (BoNT/E) reduces spontaneous recurrent seizures in a mouse model of mesial temporal lobe epilepsy. Epilepsia 2009, 50, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Duveau, V.; Madhusudan, A.; Caleo, M.; Knuesel, I.; Fritschy, J.M. Impaired reelin processing and secretion by Cajal-Retzius cells contributes to granule cell dispersion in a mouse model of temporal lobe epilepsy. Hippocampus 2011, 21, 935–944. [Google Scholar] [CrossRef]
- Antonucci, F.; Cerri, C.; Maya Vetencourt, J.F.; Caleo, M. Acute neuroprotection by the synaptic blocker botulinum neurotoxin E in a rat model of focal cerebral ischaemia. Neuroscience 2010, 169, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Akaike, N.; Kohda, T.; Torii, Y.; Goto, Y.; Harakawa, T.; Ginnaga, A.; Kaji, R.; Kozaki, S. Botulinum neurotoxin A2 reduces incidence of seizures in mouse models of temporal lobe epilepsy. Toxicon 2013, 74, 109–115. [Google Scholar] [CrossRef]
- Huang, Z.; Lian, Y.; Chen, Y.; Li, S.; Zhang, H.; Xie, N.; Zheng, Y.; Wu, S.; Wang, Y.; Cheng, W.; et al. Intranasal Delivery of Botulinum Neurotoxin A Protects against Hippocampal Neuron Death in the Lithium-Pilocarpine Rat Model. Neurochem. Res. 2019, 44, 1262–1268. [Google Scholar] [CrossRef] [PubMed]
- Gasior, M.; Tang, R.; Rogawski, M.A. Long-lasting attenuation of amygdala-kindled seizures after convection-enhanced delivery of botulinum neurotoxins A and B into the amygdala in rats. J. Pharmacol. Exp. Ther. 2013, 346, 528–534. [Google Scholar] [CrossRef] [Green Version]
- De Leonibus, E.; Costantini, V.J.; Massaro, A.; Mandolesi, G.; Vanni, V.; Luvisetto, S.; Pavone, F.; Oliverio, A.; Mele, A. Cognitive and neural determinants of response strategy in the dual-solution plus-maze task. Learn. Mem. 2011, 18, 241–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wree, A.; Mix, E.; Hawlitschka, A.; Antipova, V.; Witt, M.; Schmitt, O.; Benecke, R. Intrastriatal botulinum toxin abolishes pathologic rotational behaviour and induces axonal varicosities in the 6-OHDA rat model of Parkinson’s disease. Neurobiol. Dis. 2011, 41, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Itakura, M.; Kohda, T.; Kubo, T.; Semi, Y.; Azuma, Y.-T.; Nakajima, H.; Kozaki, S.; Takeuchi, T. Botulinum neurotoxin A subtype 2 reduces pathological behaviors more effectively than subtype 1 in a rat Parkinson’s disease model. Biochem. Biophys. Res. Comm. 2014, 447, 311–314. [Google Scholar] [CrossRef]
- Hawlitschka, A.; Wree, A. Experimental Intrastriatal Applications of Botulinum Neurotoxin-A: A Review. Int. J. Mol. Sci. 2018, 19, 1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antipova, V.; Wree, A.; Holzmann, C.; Mann, T.; Palomero-Gallagher, N.; Zilles, K.; Schmitt, O.; Hawlitschka, A. Unilateral Botulinum Neurotoxin-A Injection into the Striatum of C57BL/6 Mice Leads to a Different Motor Behavior Compared with Rats. Toxins 2018, 10, 295. [Google Scholar] [CrossRef] [Green Version]
- Hawlitschka, A.; Holzmann, C.; Wree, A.; Antipova, V. Repeated Intrastriatal Botulinum Neurotoxin-A Injection in Hemiparkinsonian Rats Increased the Beneficial Effect on Rotational Behavior. Toxins 2018, 10, 368. [Google Scholar] [CrossRef] [Green Version]
- Antipova, V.; Holzmann, C.; Hawlitschka, A.; Wree, A. Botulinum Neurotoxin-A Injected Intrastriatally into Hemiparkinsonian Rats Improves the Initiation Time for Left and Right Forelimbs in Both Forehand and Backhand Directions. Int. J. Mol. Sci. 2019, 20, 992. [Google Scholar] [CrossRef] [Green Version]
- Tsang, A.R.; Rajakumar, N.; Jog, M.S. Intrapallidal injection of botulinum toxin A recovers gait deficits in a parkinsonian rodent model. Acta Physiol. 2019, 226, e13230. [Google Scholar] [CrossRef]
- Tsang, A.R.; Rajakumar, N.; Jog, M.S. Botulinum toxin A injection into the entopeduncular nucleus improves dynamic locomotory parameters in hemiparkinsonian rats. PLoS ONE 2019, 14, e0223450. [Google Scholar] [CrossRef]
- Antipova, V.; Holzmann, C.; Hawlitschka, A.; Witt, M.; Wree, A. Antidepressant-Like Properties of Intrastriatal Botulinum Neurotoxin-A Injection in a Unilateral 6-OHDA Rat Model of Parkinson’s Disease. Toxins 2021, 13, 505. [Google Scholar] [CrossRef]
- Ibragic, S.; Matak, I.; Dracic, A.; Smajlovic, A.; Muminovic, M.; Proft, F.; Sofic, E.; Lackovic, Z.; Riederer, P. Effects of botulinum toxin type A facial injection on monoamines and their metabolites in sensory, limbic and motor brain regions in rats. Neurosci. Lett. 2016, 617, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Mann, T.; Zilles, K.; Dikow, H.; Hellfritsch, A.; Cremer, M.; Piel, M.; Rosch, F.; Hawlitschka, A.; Schmitt, O.; Wree, A. Dopamine, Noradrenaline and Serotonin Receptor Densities in the Striatum of Hemiparkinsonian Rats following Botulinum Neurotoxin-A Injection. Neuroscience 2018, 374, 187–204. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Liu, X.; Sul, C.-J.; Zhang, Q.-L.; Wang, Z.-H.; Cao, L.-F.; Guo, X.-Y.; Huang, Y.; Luo, W.; et al. Antidepressant-Like Action of Single Facial Injection of Botulinum Neurotoxin A is Associated with Augmented 5-HT Levels and BDNF/ERK/CREB Pathways in Mouse Brain. Neurosci. Bull. 2019, 35, 661–672. [Google Scholar] [CrossRef]
- Li, Y.; Liu, T.; Luo, W. Botulinum Neurotoxin Therapy for Depression: Therapeutic Mechanisms and Future Perspective. Front. Psychiatry 2021, 12, 584416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, W.; Fan, Y.; Li, Y.; Liu, J.; Xu, Y.; Jiang, C.; Tang, Z.; Cao, C.; Liu, T.; et al. The safety and efficacy of botulinum toxin A on the treatment of depression. Brain Behav. 2021, 11, e2333. [Google Scholar] [CrossRef] [PubMed]
- Yesudhas, A.; Radhakrishnan, R.K.; Sukesh, A.; Ravichandran, S.; Manickam, N.; Kandasamy, M. BOTOX® counteracts the innate anxiety-related behaviours in correlation with increased activities of key antioxidant enzymes in the hippocampus of ageing experimental mice. Biochem. Biophys. Res. Commun. 2021, 569, 54–60. [Google Scholar] [CrossRef]
- Romo, P.G.B.; Smith, C.P.; Cox, A.; Averbeck, M.A.; Dowling, C.; Beckford, C.; Manohar, P.; Duran, S.; Cameron, A.P. Non-surgical urologic management of neurogenic bladder after spinal cord injury. World J. Urol. 2018, 36, 1555–1568. [Google Scholar] [CrossRef]
- Palazón-García, R.; Benavente-Valdepeñas, A.M. Botulinum Toxin: From Poison to Possible Treatment for Spasticity in Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 4886. [Google Scholar] [CrossRef]
- Marinelli S, Luvisetto S, Cobianchi S, Makuch W, Obara I, Mezzaroma E, Caruso M, Straface E, Przewlocka B, Pavone F Botulinum neurotoxin type A counteracts neuropathic pain and facilitates functional recovery after peripheral nerve injury in animal models. Neuroscience 2010, 171, 316–328. [CrossRef] [PubMed]
- Lee, W.H.; Shin, T.J.; Kim, H.J.; Lee, J.K.; Suh, H.W.; Lee, S.C.; Seo, K. Intrathecal administration of botulinum neurotoxin type A attenuates formalin-induced nociceptive responses in mice. Anesth. Analg. 2011, 112, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.; Oliveira, R.; Rossetto, O.; Cruz, C.D.; Cruz, F.; Avelino, A. Intrathecal administration of botulinum toxin type A improves urinary bladder function and reduces pain in rats with cystitis. Eur. J. Pain 2014, 18, 1480–1489. [Google Scholar] [CrossRef] [PubMed]
- Drinovac, V.; Bach-Rojecky, L.; Lacković, Z. Antinociceptive action of botulinum toxin type A in carrageenan-induced mirror pain. J. Neural Transm. 2016, 123, 1403–1413. [Google Scholar] [CrossRef] [Green Version]
- Coelho, A.; Oliveira, R.; Cruz, F.; Cruz, C.D. Impairment of sensory afferents by intrathecal administration of botulinum toxin A improves neurogenic detrusor overactivity in chronic spinal cord injured rats. Exp. Neurol. 2016, 285, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, R.; Sun, Y.; Li, H.; Ma, D.; Zhang, C.; Guan, Y.; Li, J.; Wang, Y. Botulinum toxin type A and gabapentin attenuate postoperative pain and NK1 receptor internalization in rats. Neurochem. Int. 2018, 116, 52–62. [Google Scholar] [CrossRef]
- Huang, P.P.; Khan, I.; Suhail, M.S.; Malkmus, S.; Yaksh, T.L. Spinal botulinum neurotoxin B: Effects on afferent transmitter release and nociceptive processing. PLoS ONE 2011, 6, e19126. [Google Scholar]
- Park, H.J.; Marino, M.J.; Rondon, E.S.; Xe, Q.; Yaksh, T.L. The effects of intraplantar and intrathecal botulinum toxin type B on tactile allodynia in mono and polyneuropathy in the mouse. Anesth. Analg. 2015, 121, 229–238. [Google Scholar] [CrossRef]
- Vacca, V.; Madaro, L.; De Angelis, F.; Proietti, D.; Cobianchi, S.; Orsini, T.; Puri, P.L.; Luvisetto, S.; Pavone, F.; Marinelli, S. Revealing the therapeutic potential of Botulinum neurotoxin type A in counteracting paralysis and neuropathic pain in spinally injured mice. Toxins 2020, 12, 491. [Google Scholar] [CrossRef]
- Ferrari, E.; Maywood, E.S.; Restani, L.; Caleo, M.; Pirazzini, M.; Rossetto, O.; Hastings, M.H.; Niranjan, D.; Schiavo, G.; Davletov, B. Re-assembled botulinum neurotoxin inhibits CNS functions without systemic toxicity. Toxins 2011, 3, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Mangione, A.S.; Obara, I.; Maiarù, M.; Geranton, S.M.; Tasorelli, C.; Ferrari, E.; Leese, C.; Davletov, B.; Hunt, S.P. Nonparalytic botulinum molecules for the control of pain. Pain 2016, 157, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Maiarù, M.; Leese, C.; Certo, M.; Echeverria-Altuna, I.; Mangione, A.S.; Arsenault, J.; Davletov, B.; Hunt, S.P. Selective neuronal silencing using synthetic botulinum molecules alleviates chronic pain in mice. Sci. Transl. Med. 2018, 10, 7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, L.; Peng, L.; Berntsson, R.P.; Liu, S.M.; Park, S.; Yu, F.; Boone, C.; Palan, S.; Beard, M.; Chabrier, P.E.; et al. Engineered botulinum neurotoxin B with improved efficacy for targeting human receptors. Nat. Commun. 2017, 8, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, R.P. Engineering of Botulinum Neurotoxins for Biomedical Applications. Toxins 2018, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Fonfria, E.; Elliott, M.; Beard, M.; Chaddock, J.A.; Krupp, J. Engineering Botulinum Toxins to Improve and Expand Targeting and SNARE Cleavage Activity. Toxins 2018, 10, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Meng, J.; Wang, J. New Engineered-Botulinum Toxins Inhibit the Release of Pain-Related mediators. Int. J. Mol. Sci. 2019, 21, 262. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.; Favre-Guilmard, C.; Liu, S.M.; Maignel, J.; Masuyer, G.; Beard, M.; Boone, C.; Carré, D.; Kalinichev, M.; Lezmi, S.; et al. Engineered botulinum neurotoxin B with improved binding to human receptors has enhanced efficacy in preclinical models. Sci. Adv. 2019, 5, 7196. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Feng, Y.; Guan, Y. A Novel Botulinum Toxin TAT-EGFP-HCS Fusion Protein Capable of Specific Delivery Through the Blood-brain Barrier to the Central Nervous System. CNS Neurol. Disord. Drug Targets 2019, 18, 37–43. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Popoff, M.R. Engineering Botulinum Neurotoxins for Enhanced Therapeutic Applications and Vaccine Development. Toxins 2020, 13, 1. [Google Scholar] [CrossRef]
- Sikorra, S.; Donald, S.; Elliott, M.; Schwede, S.; Coker, S.F.; Kupinski, A.P.; Tripathi, V.; Foster, K.; Beard, M.; Binz, T. Engineering an Effective Human SNAP-23 Cleaving Botulinum Neurotoxin A Variant. Toxins 2020, 12, 804. [Google Scholar] [CrossRef]
- Blum, T.R.; Liu, H.; Packer, M.S.; Xiong, X.; Lee, P.G.; Zhang, S.; Richter, M.; Minasov, G.; Satchell, K.J.F.; Dong, M.; et al. Phage-assisted evolution of botulinum neurotoxin proteases with reprogrammed specificity. Science 2021, 371, 803–810. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luvisetto, S. Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy. Toxins 2021, 13, 751. https://doi.org/10.3390/toxins13110751
Luvisetto S. Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy. Toxins. 2021; 13(11):751. https://doi.org/10.3390/toxins13110751
Chicago/Turabian StyleLuvisetto, Siro. 2021. "Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy" Toxins 13, no. 11: 751. https://doi.org/10.3390/toxins13110751
APA StyleLuvisetto, S. (2021). Botulinum Neurotoxins in Central Nervous System: An Overview from Animal Models to Human Therapy. Toxins, 13(11), 751. https://doi.org/10.3390/toxins13110751