Emergence of New ST301 Shiga Toxin-Producing Escherichia coli Clones Harboring Extra-Intestinal Virulence Traits in Europe

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
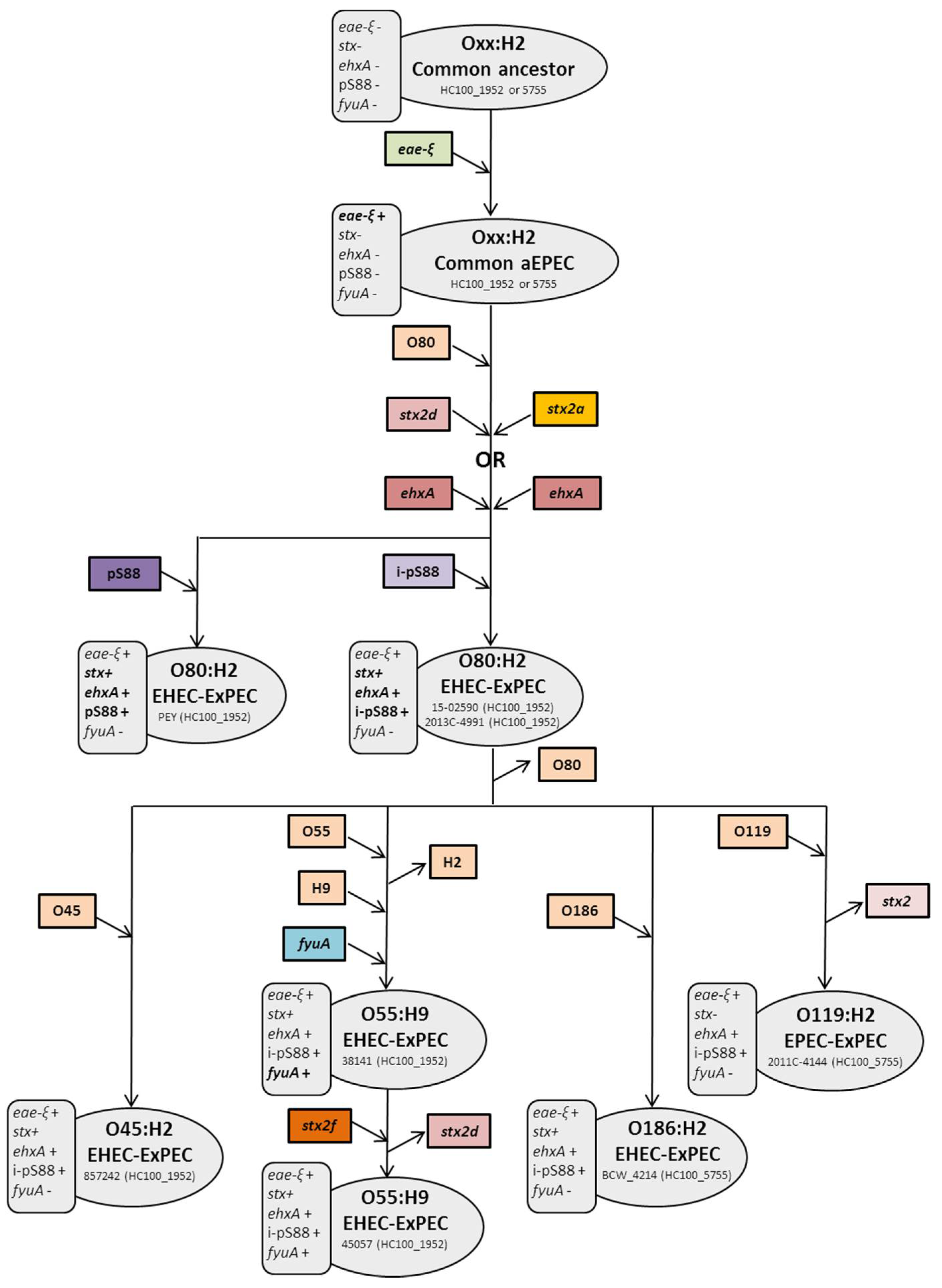
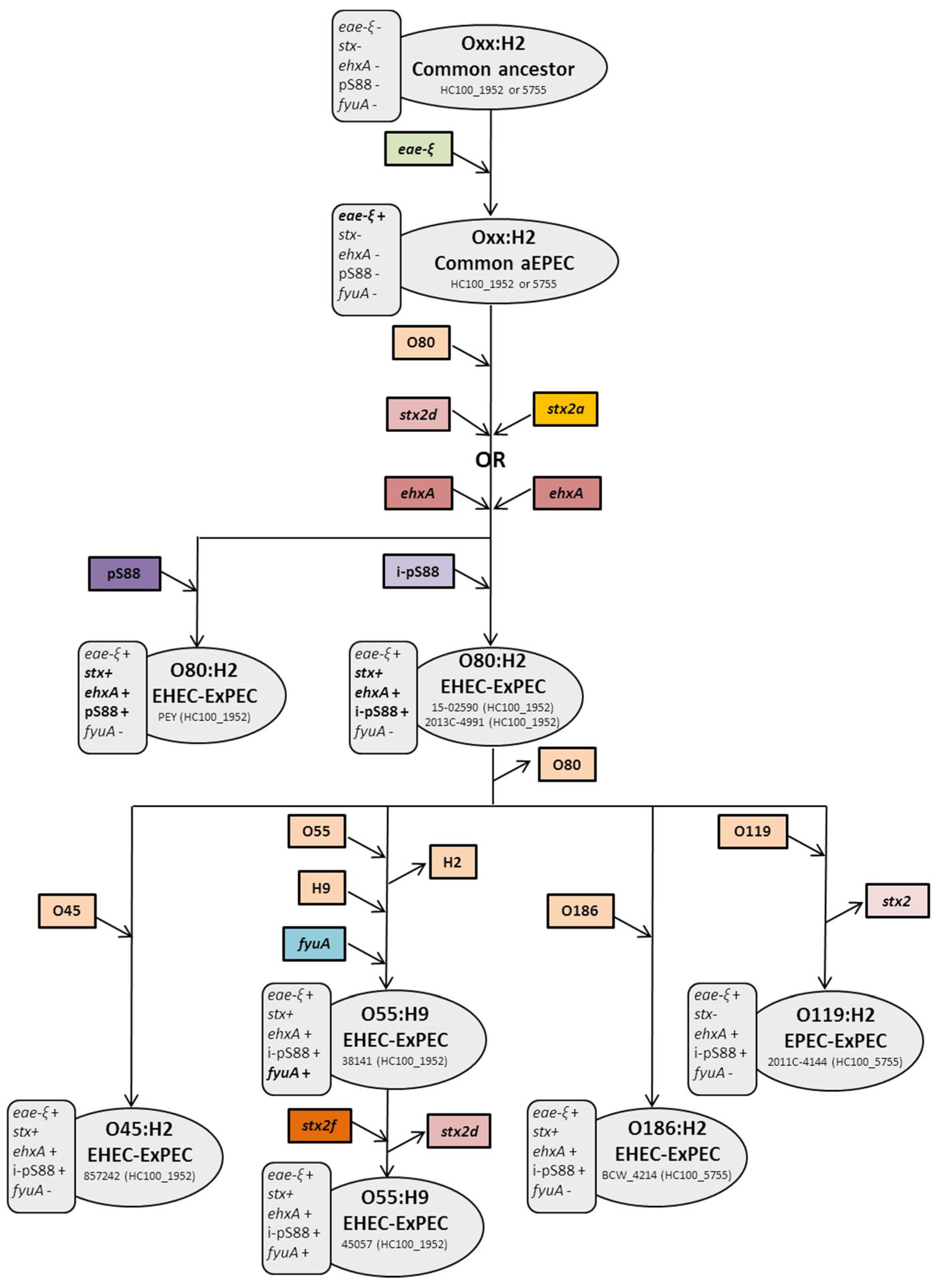
2.1. ST301 Characterization and Phylogeny
2.2. The O55:H9 Serotype and Its Relationship with the O80 Serogroup
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains, Strain Sequences from a Database, and Sampling
5.2. Whole Genome Sequencing
5.3. Sequence Analysis
5.4. pS88-like Plasmid
5.5. Phylogeny
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Garg, A.X.; Suri, R.S.; Barrowman, N.; Rehman, F.; Matsell, D.; Rosas-Arellano, M.P.; Salvadori, M.; Haynes, R.B.; Clark, W.F. Long-Term Renal Prognosis of Diarrhea-Associated Hemolytic Uremic Syndrome: A Systematic Review, Meta-Analysis, and Meta-Regression. JAMA 2003, 290, 1360–1370. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Schiller, R.; Lammers, L.; Bauwens, A.; Fruth, A.; Middendorf, B.; Schmidt, M.A.; Tarr, P.I.; Dobrindt, U.; Karch, H.; et al. Heteropathogenic Virulence and Phylogeny Reveal Phased Pathogenic Metamorphosis in Escherichia coli O2:H6. EMBO Mol. Med. 2014, 6, 347–357. [Google Scholar] [CrossRef]
- Mariani-Kurkdjian, P.; Lemaître, C.; Bidet, P.; Perez, D.; Boggini, L.; Kwon, T.; Bonacorsi, S. Haemolytic-Uraemic Syndrome with Bacteraemia Caused by a New Hybrid Escherichia coli Pathotype. New Microbes New Infect. 2014, 2, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Soysal, N.; Mariani-Kurkdjian, P.; Smail, Y.; Liguori, S.; Gouali, M.; Loukiadis, E.; Fach, P.; Bruyand, M.; Blanco, J.; Bidet, P.; et al. Enterohemorrhagic Escherichia coli Hybrid Pathotype O80:H2 as a New Therapeutic Challenge. Emerg. Infect. Dis. 2016, 22, 1604–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruyand, M.; Mariani-Kurkdjian, P.; Le Hello, S.; King, L.-A.; Van Cauteren, D.; Lefevre, S.; Gouali, M.; Jourdan-da Silva, N.; Mailles, A.; Donguy, M.-P.; et al. Paediatric Haemolytic Uraemic Syndrome Related to Shiga Toxin-Producing Escherichia coli, an Overview of 10 Years of Surveillance in France, 2007 to 2016. Eurosurveillance 2019, 24, 1800068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nüesch-Inderbinen, M.; Cernela, N.; Wüthrich, D.; Egli, A.; Stephan, R. Genetic Characterization of Shiga Toxin Producing Escherichia coli Belonging to the Emerging Hybrid Pathotype O80:H2 Isolated from Humans 2010-2017 in Switzerland. Int. J. Med. Microbiol. 2018, 308, 534–538. [Google Scholar] [CrossRef] [Green Version]
- De Rauw, K.; Thiry, D.; Caljon, B.; Saulmont, M.; Mainil, J.; Piérard, D. Characteristics of Shiga Toxin Producing- and Enteropathogenic Escherichia coli of the Emerging Serotype O80:H2 Isolated from Humans and Diarrhoeic Calves in Belgium. Clin. Microbiol. Infect. 2019, 25, 111.e5–111.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijnsma, K.L.; Schijvens, A.M.; Rossen, J.W.A.; Kooistra-Smid, A.M.D.M.; Schreuder, M.F.; van de Kar, N.C.A.J. Unusual Severe Case of Hemolytic Uremic Syndrome Due to Shiga Toxin 2d-Producing E. coli O80:H2. Pediatr. Nephrol. 2017, 32, 1263–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierz, L.; Cernela, N.; Hauser, E.; Nüesch-Inderbinen, M.; Stephan, R. Characteristics of Shigatoxin-Producing Escherichia coli Strains Isolated during 2010–2014 from Human Infections in Switzerland. Front. Microbiol. 2017, 8, 1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority; European Centre for Disease Prevention and Control The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [CrossRef]
- Cointe, A.; Birgy, A.; Mariani-Kurkdjian, P.; Liguori, S.; Courroux, C.; Blanco, J.; Delannoy, S.; Fach, P.; Loukiadis, E.; Bidet, P.; et al. Emerging Multidrug-Resistant Hybrid Pathotype Shiga Toxin-Producing Escherichia coli O80 and Related Strains of Clonal Complex 165, Europe. Emerg. Infect. Dis. 2018, 24, 2262–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The Population Genetics of Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Blanco, J.E.; Mora, A.; Dahbi, G.; Alonso, M.P.; González, E.A.; Bernárdez, M.I.; Blanco, J. Serotypes, Virulence Genes, and Intimin Types of Shiga Toxin (Verotoxin)-Producing Escherichia coli Isolates from Cattle in Spain and Identification of a New Intimin Variant Gene (eae-ξ). J. Clin. Microbiol. 2004, 42, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Thiry, D.; Saulmont, M.; Takaki, S.; De Rauw, K.; Duprez, J.-N.; Iguchi, A.; Piérard, D.; Mainil, J.G. Enteropathogenic Escherichia coli O80:H2 in Young Calves with Diarrhea, Belgium. Emerg. Infect. Dis. 2017, 23, 2093–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habets, A.; Crombé, F.; Nakamura, K.; Guérin, V.; De Rauw, K.; Piérard, D.; Saulmont, M.; Hayashi, T.; Mainil, J.G.; Thiry, D. Genetic Characterization of Shigatoxigenic and Enteropathogenic Escherichia coli O80:H2 from Diarrhoeic and Septicaemic Calves and Relatedness to Human Shigatoxigenic E. Coli O80:H2. J. Appl. Microbiol. 2020, 130, 258–264. [Google Scholar] [CrossRef]
- Cointe, A.; Birgy, A.; Pascault, A.; Louillet, F.; Dufougeray, A.; Mariani-Kurkdjian, P.; Bonacorsi, S. Be Aware of Shiga-Toxin 2f-Producing Escherichia coli: Case Report and False-Negative Results with Certain Rapid Molecular Panels. Diagn. Microbiol. Infect. Dis. 2020, 98, 115177. [Google Scholar] [CrossRef]
- Jones, G. Surveillance Du Syndrome Hémolytique et Urémique Post-Diarrhéique Chez l’enfant de Moins de 15 Ans En France En 2019; Santé Publique France: Saint Maurice, France, 2019. [Google Scholar]
- Ju, W.; Cao, G.; Rump, L.; Strain, E.; Luo, Y.; Timme, R.; Allard, M.; Zhao, S.; Brown, E.; Meng, J. Phylogenetic Analysis of Non-O157 Shiga Toxin-Producing Escherichia coli Strains by Whole-Genome Sequencing. J. Clin. Microbiol. 2012, 50, 4123–4127. [Google Scholar] [CrossRef] [Green Version]
- Lacher, D.W.; Steinsland, H.; Whittam, T.S. Allelic Subtyping of the Intimin Locus (eae) of Pathogenic Escherichia coli by Fluorescent RFLP. FEMS Microbiol. Lett. 2006, 261, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Sun, H.; Fan, R.; Fu, S.; Zhang, J.; Matussek, A.; Xiong, Y.; Bai, X. Genetic Diversity of the Intimin Gene (eae) in Non-O157 Shiga Toxin-Producing Escherichia coli Strains in China. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Cookson, A.L.; Bennett, J.; Thomson-Carter, F.; Attwood, G.T. Intimin Subtyping of Escherichia coli: Concomitant Carriage of Multiple Intimin Subtypes from Forage-Fed Cattle and Sheep. FEMS Microbiol. Lett. 2007, 272, 163–171. [Google Scholar] [CrossRef]
- Hua, Y.; Bai, X.; Zhang, J.; Jernberg, C.; Chromek, M.; Hansson, S.; Frykman, A.; Yang, X.; Xiong, Y.; Wan, C.; et al. Molecular Characteristics of eae-Positive Clinical Shiga Toxin-Producing Escherichia coli in Sweden. Emerg. Microbes. Infect. 2020, 9, 2562–2570. [Google Scholar] [CrossRef]
- Patel, P.N.; Lindsey, R.L.; Garcia-Toledo, L.; Rowe, L.A.; Batra, D.; Whitley, S.W.; Drapeau, D.; Stoneburg, D.; Martin, H.; Juieng, P.; et al. High-Quality Whole-Genome Sequences for 77 Shiga Toxin-Producing Escherichia coli Strains Generated with PacBio Sequencing. Genome Announc. 2018, 6, e00391-18. [Google Scholar] [CrossRef] [Green Version]
- Gigliucci, F.; van Hoek, A.H.A.M.; Chiani, P.; Knijn, A.; Minelli, F.; Scavia, G.; Franz, E.; Morabito, S.; Michelacci, V. Genomic Characterization of HlyF-Positive Shiga Toxin–Producing Escherichia coli, Italy and the Netherlands, 2000–2019. Emerg. Infect. Dis. J. 2021, 27, 853–861. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel Pathogenicity Assessment of Shiga Toxin Producing Escherichia coli (Stec) and the Public Health Risk Posed by Contamination of Food with Stec. EFSA J. 2020, 18, 5967.
- Galardini, M.; Clermont, O.; Baron, A.; Busby, B.; Dion, S.; Schubert, S.; Beltrao, P.; Denamur, E. Major Role of Iron Uptake Systems in the Intrinsic Extra-Intestinal Virulence of the Genus Escherichia Revealed by a Genome-Wide Association Study. PLOS Genet. 2020, 16, e1009065. [Google Scholar] [CrossRef] [PubMed]
- Peigne, C.; Bidet, P.; Mahjoub-Messai, F.; Plainvert, C.; Barbe, V.; Médigue, C.; Frapy, E.; Nassif, X.; Denamur, E.; Bingen, E.; et al. The Plasmid of Escherichia coli Strain S88 (O45:K1:H7) That Causes Neonatal Meningitis Is Closely Related to Avian Pathogenic E. Coli Plasmids and Is Associated with High-Level Bacteremia in a Neonatal Rat Meningitis Model. Infect. Immun. 2009, 77, 2272–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Fan, Y.; Achtman, M. The EnteroBase User’s Guide, with Case Studies on Salmonella Transmissions, Yersinia pestis Phylogeny, and Escherichia Core Genomic Diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [Green Version]
- Serotype Finder. Available online: Https://Cge.Cbs.Dtu.Dk/Services/SerotypeFinder (accessed on 25 March 2021).
- CGE. Available online: Http://Www.Genomicepidemiology.Org/ (accessed on 25 March 2021).
- BLAST+ 2.2.31. Available online: Https://Blast.Ncbi.Nlm.Nih.Gov/Blast.Cgi (accessed on 25 March 2021).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cointe, A.; Bizot, E.; Delannoy, S.; Fach, P.; Bidet, P.; Birgy, A.; Weill, F.-X.; Lefèvre, S.; Mariani-Kurkdjian, P.; Bonacorsi, S. Emergence of New ST301 Shiga Toxin-Producing Escherichia coli Clones Harboring Extra-Intestinal Virulence Traits in Europe. Toxins 2021, 13, 686. https://doi.org/10.3390/toxins13100686
Cointe A, Bizot E, Delannoy S, Fach P, Bidet P, Birgy A, Weill F-X, Lefèvre S, Mariani-Kurkdjian P, Bonacorsi S. Emergence of New ST301 Shiga Toxin-Producing Escherichia coli Clones Harboring Extra-Intestinal Virulence Traits in Europe. Toxins. 2021; 13(10):686. https://doi.org/10.3390/toxins13100686
Chicago/Turabian StyleCointe, Aurélie, Etienne Bizot, Sabine Delannoy, Patrick Fach, Philippe Bidet, André Birgy, François-Xavier Weill, Sophie Lefèvre, Patricia Mariani-Kurkdjian, and Stéphane Bonacorsi. 2021. "Emergence of New ST301 Shiga Toxin-Producing Escherichia coli Clones Harboring Extra-Intestinal Virulence Traits in Europe" Toxins 13, no. 10: 686. https://doi.org/10.3390/toxins13100686
APA StyleCointe, A., Bizot, E., Delannoy, S., Fach, P., Bidet, P., Birgy, A., Weill, F.-X., Lefèvre, S., Mariani-Kurkdjian, P., & Bonacorsi, S. (2021). Emergence of New ST301 Shiga Toxin-Producing Escherichia coli Clones Harboring Extra-Intestinal Virulence Traits in Europe. Toxins, 13(10), 686. https://doi.org/10.3390/toxins13100686