How the Neurotoxin β-N-Methylamino-l-Alanine Accumulates in Bivalves: Distribution of the Different Accumulation Fractions among Organs

 , ,
, ,  ,
, 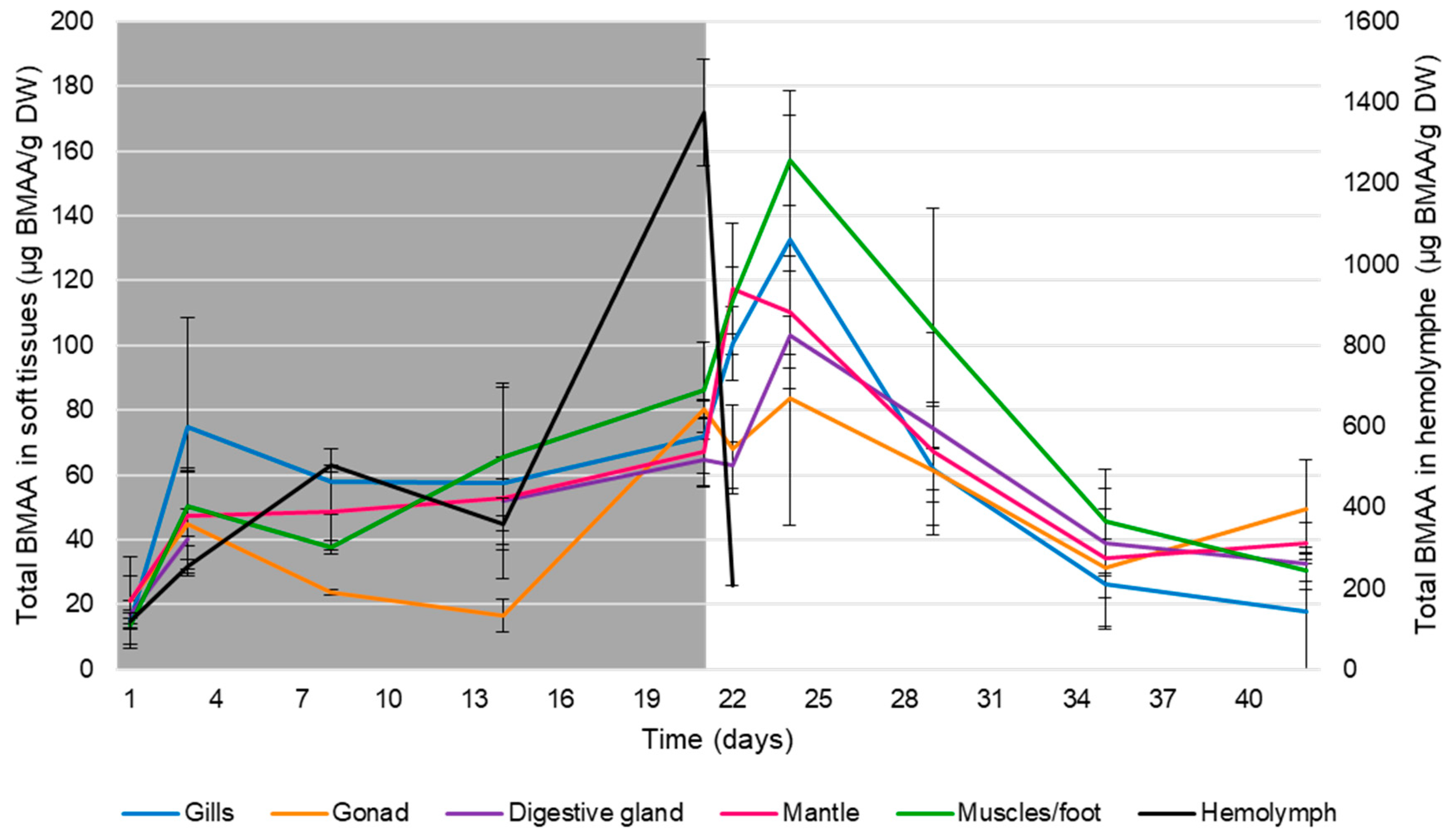{kind=link}
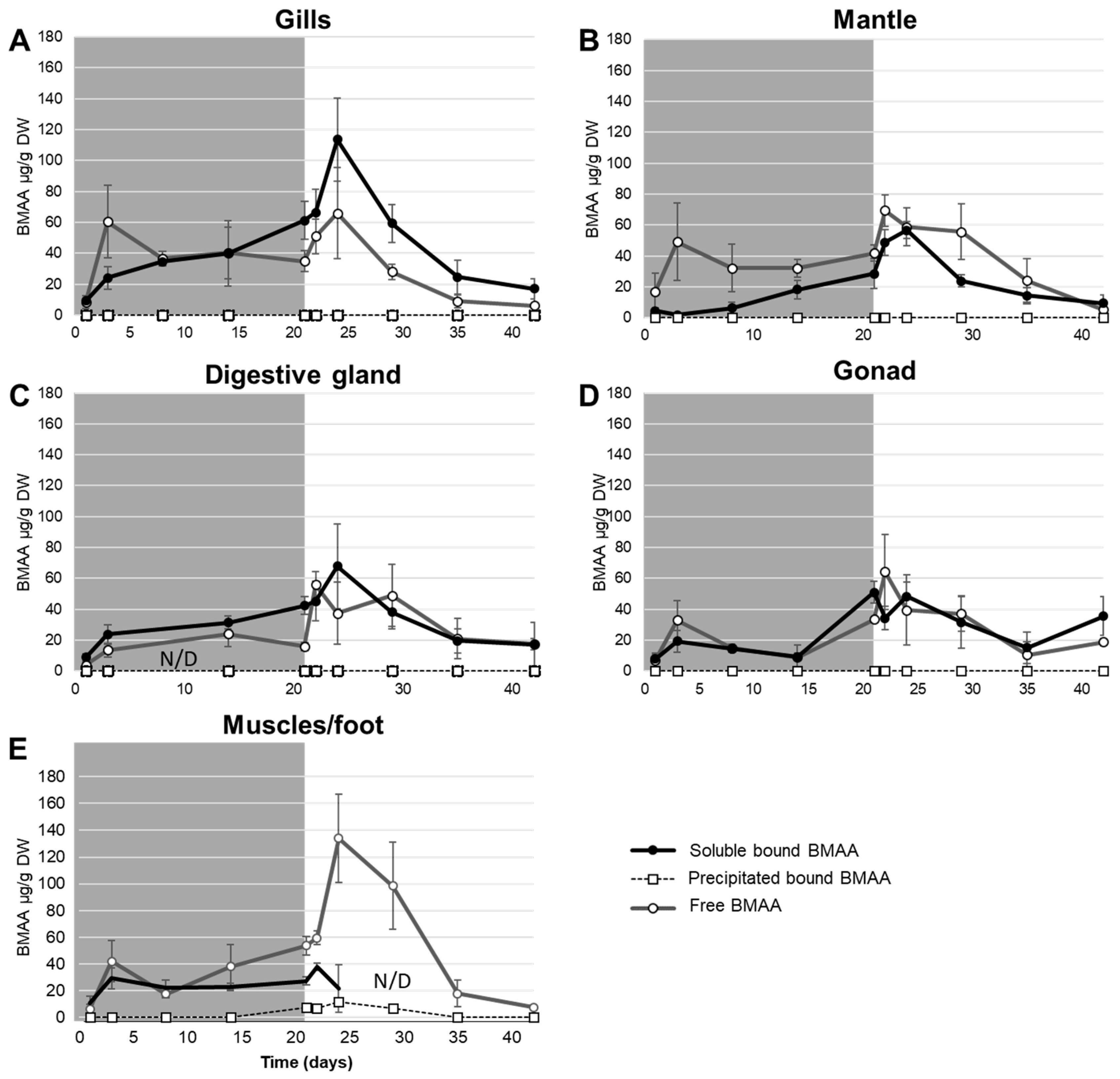{kind=link}
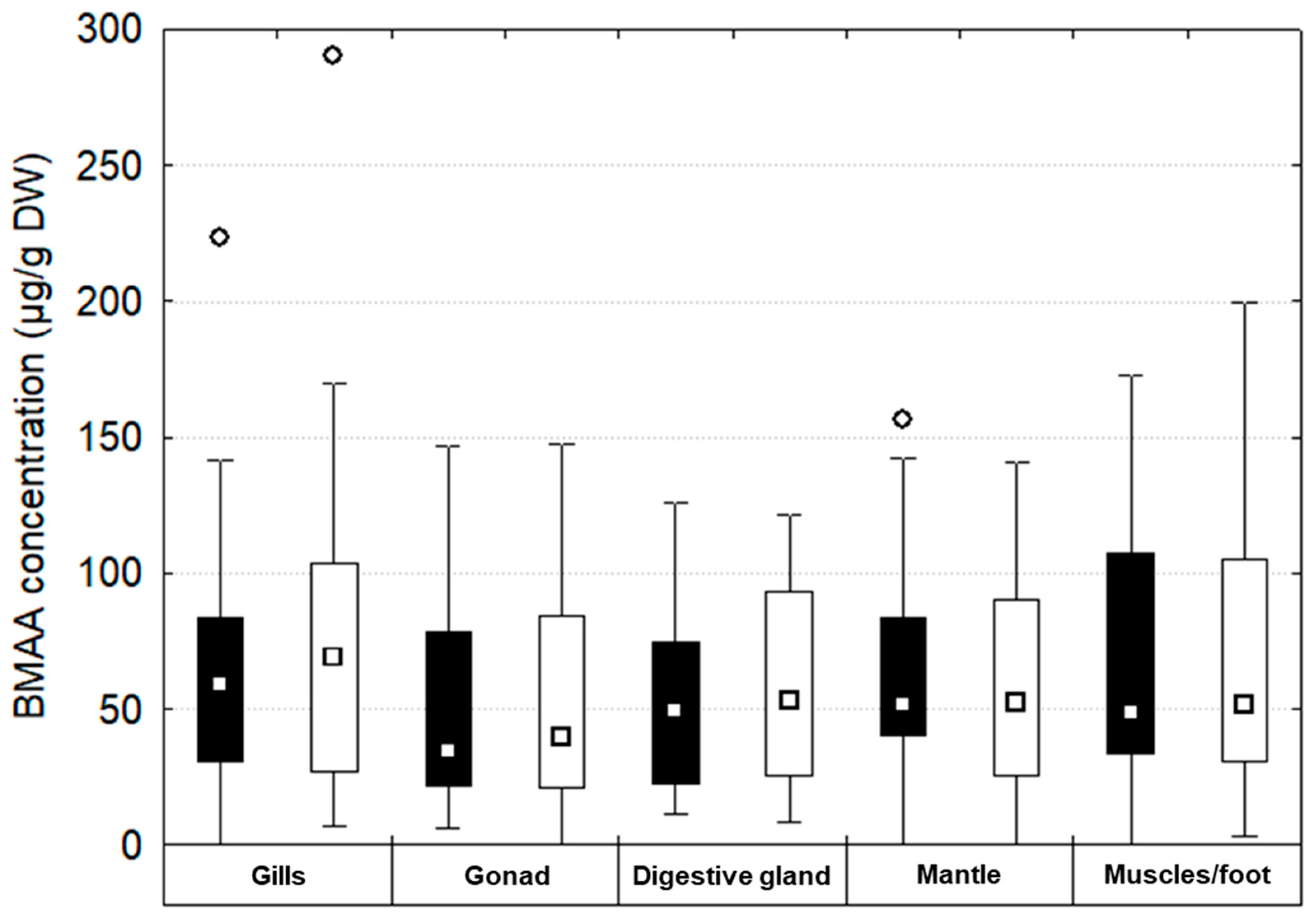{kind=link}
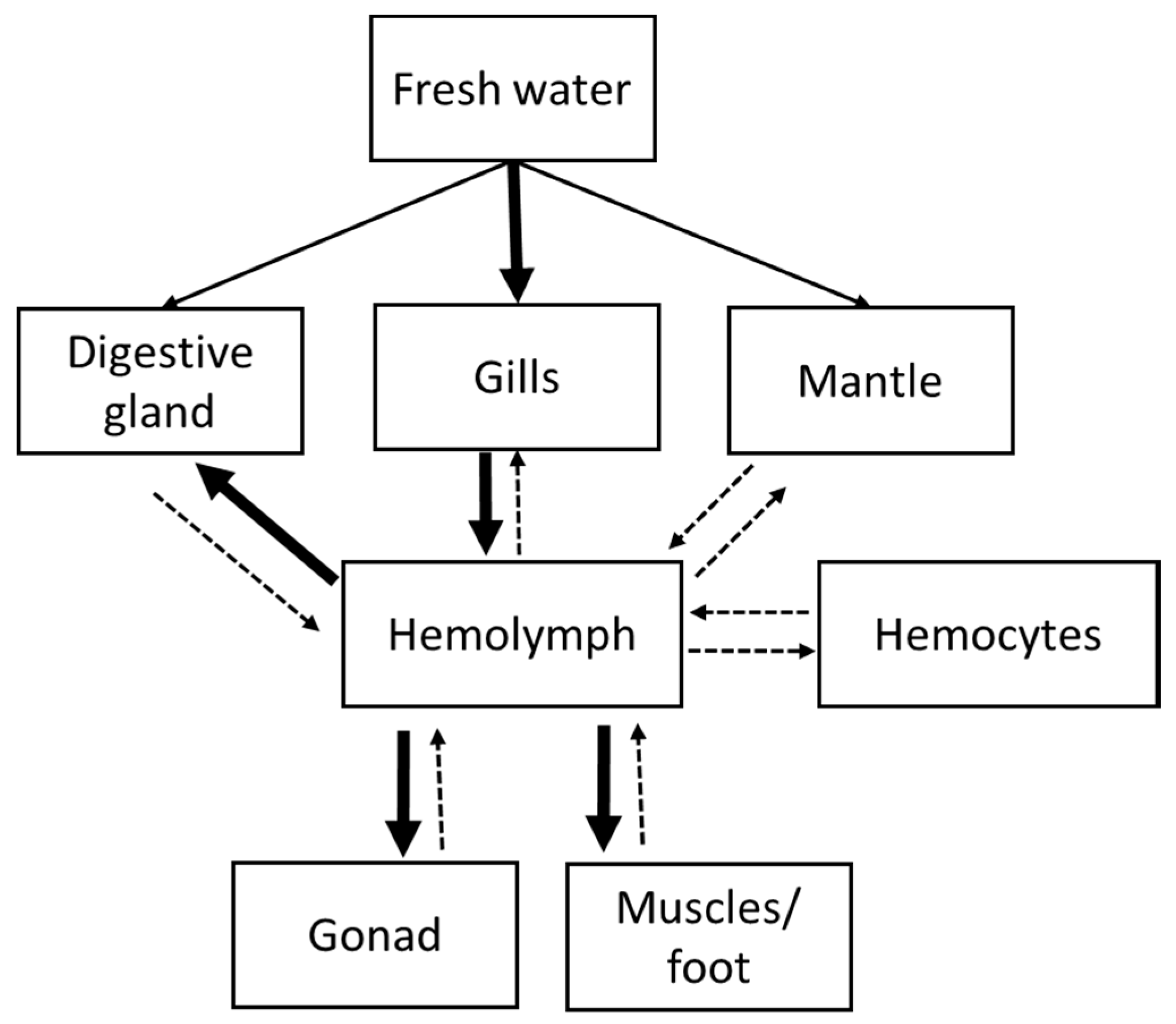{kind=link}
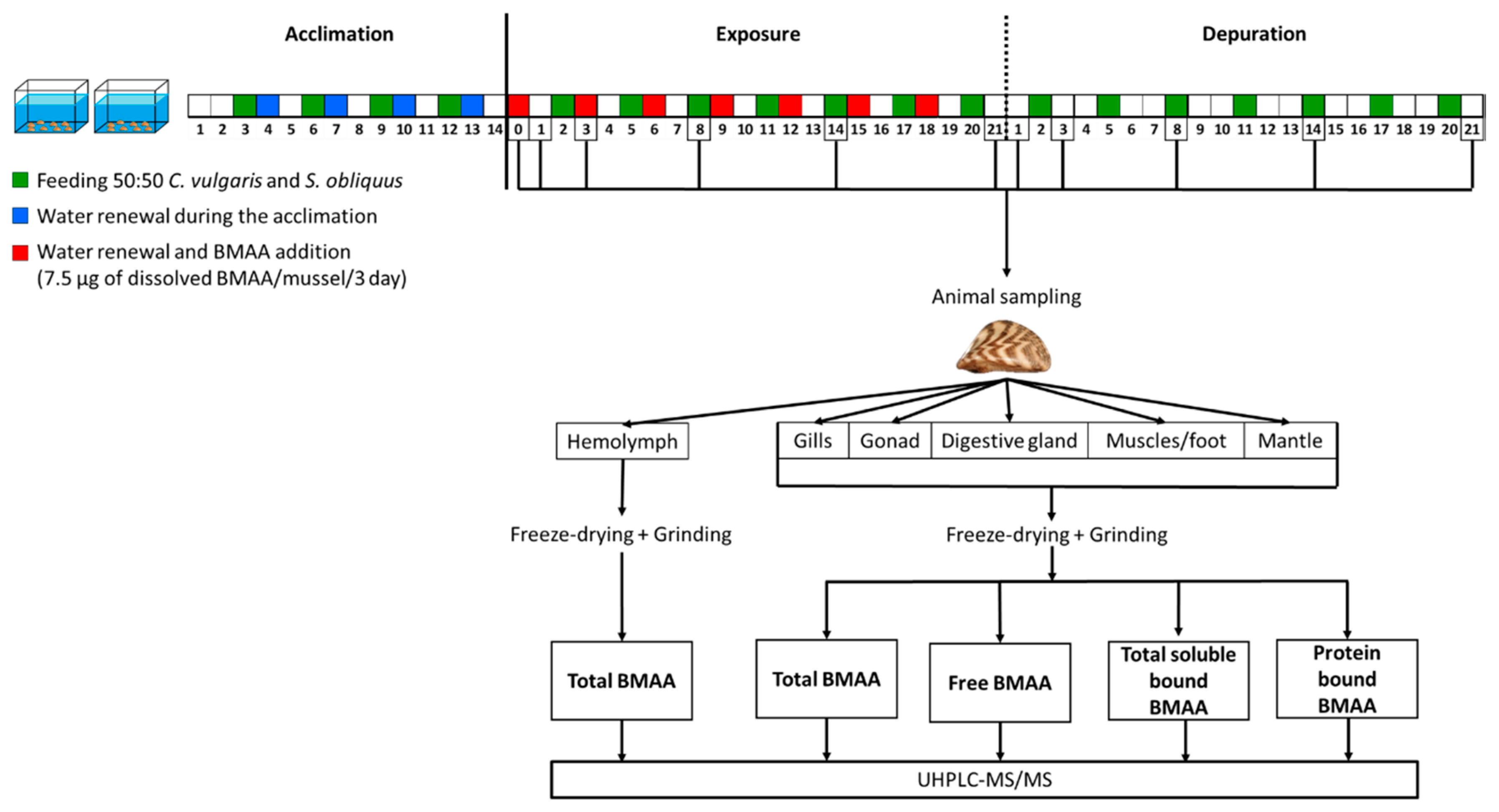{kind=link}
Abstract
1. Introduction
2. Results
3. Discussion
3.1. Organ Distribution of BMAA in D. polymorpha Over Time of Exposure and Depuration
3.2. BMAA Accumulation Fractions in Tissues of D. polymorpha
3.3. BMAA Kinetics of Accumulation and Detoxification Within Tissues
3.4. Use of Bivalves to Monitor Environmental BMAA: Pertinent Fractions and Tissues
4. Conclusions
5. Materials and Methods
5.1. Mussels Sampling and Acclimation
5.2. Exposure
5.2.1. Experimental Setting
5.2.2. Tissue Sampling
5.3. Extraction
5.3.1. Total BMAA
5.3.2. Free and Soluble Bound BMAA
5.3.3. Precipitated Bound BMAA
5.4. UHPLC-MS/MS Analysis
5.5. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Pablo, J.; Banack, S.A.; Cox, P.A.; Johnson, T.E.; Papapetropoulos, S.; Bradley, W.G.; Buck, A.; Mash, D.C. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s disease. Acta Neurol. Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.S.; Gehringer, M.M.; Welch, J.H.; Neilan, B.A. Does α-amino-β-methylaminopropionic acid (BMAA) play a role in neurodegeneration? Int. J. Environ. Res. Public Health 2011, 8, 3728–3746. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-methylamino-L-alanine (BMAA) in. ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D. Mechanisms of beta-N-methylamino-l-alanine induced neurotoxicity. Amyotroph. Lateral Scler. 2009, 10 (Suppl. 2), 56–60. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-Protein Amino Acid BMAA Is Misincorporated into Human Proteins in Place of l-Serine Causing Protein Misfolding and Aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [PubMed]
- Onselen, R.; Downing, S.; Kemp, G.; Downing, T. Investigating β-N-Methylamino-l-alanine Misincorporation in Human Cell Cultures: A Comparative Study with Known Amino Acid Analogues. Toxins 2017, 9, 400. [Google Scholar] [CrossRef]
- Delcourt, N.; Claudepierre, T.; Maignien, T.; Arnich, N.; Mattei, C. Cellular and Molecular Aspects of the β-N-Methylamino-l-alanine (BMAA) Mode of Action within the Neurodegenerative Pathway: Facts and Controversy. Toxins 2017, 10, 6. [Google Scholar] [CrossRef]
- Monteiro, M.; Costa, M.; Moreira, C.; Vasconcelos, V.M.; Baptista, M.S. Screening of BMAA-producing cyanobacteria in cultured isolates and in in situ blooms. J. Appl. Phycol. 2016. [Google Scholar] [CrossRef]
- Réveillon, D.; Abadie, E.; Séchet, V.; Brient, L.; Savar, V.; Bardouil, M.; Hess, P.; Amzil, Z. Beta-N-methylamino-l-alanine: LC-MS/MS optimization, screening of cyanobacterial strains and occurrence in shellfish from Thau, a French Mediterranean lagoon. Mar. Drugs 2014, 12, 5441–5467. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and bacteria isolated from a diatom culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef]
- Réveillon, D.; Abadie, E.; Séchet, V.; Masseret, E.; Hess, P.; Amzil, Z. β-N-methylamino-l-alanine (BMAA) and isomers: Distribution in different food web compartments of Thau lagoon, French Mediterranean Sea. Mar. Environ. Res. 2015, 110, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and Isomers in Aquatic Environments and Aquatic Food Sources for Humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E. Presence of the Neurotoxin BMAA in Aquatic Ecosystems: What Do We Really Know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Broc Glover, W.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental Neurotoxins β-N-Methylamino-l-Alanine (BMAA) and Mercury in Shark Cartilage Dietary Supplements. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spácil, Z.; Ilag, L.L.; Ronnevi, L.O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef]
- Lage, S.; Annadotter, H.; Rasmussen, U.; Rydberg, S. Biotransfer of β-N-methylamino-l-alanine (BMAA) in a eutrophicated freshwater lake. Mar. Drugs 2015, 13, 1185–1201. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed]
- Rosén, J.; Hellenäs, K.E. Determination of the neurotoxin BMAA (β-N-methylamino-l-alanine) in cycad seed and cyanobacteria by LC-MS/MS (liquid chromatography tandem mass spectrometry). Analyst 2008, 133, 1785. [Google Scholar] [CrossRef]
- Kubo, T.; Kato, N.; Hosoya, K.; Kaya, K. Effective determination method for a cyanobacterial neurotoxin, β-N-methylamino-l-alanine. Toxicon 2008, 51, 1264–1268. [Google Scholar] [CrossRef]
- Oehlmann, J.; Schulte-Oehlmann, U. Chapter 17 Molluscs as bioindicators. Trace Met. Other Contam. Environ. 2003, 6, 577–635. [Google Scholar] [CrossRef]
- Gadzała-Kopciuch, R.; Berecka, B.; Bartoszewicz, J.; Buszewski, B. Some considerations about bioindicators in environmental monitoring. Pol. J. Environ. Stud. 2004, 13, 453–462. [Google Scholar]
- Li, L.; Zheng, B.; Liu, L. Biomonitoring and Bioindicators Used for River Ecosystems: Definitions, Approaches and Trends. Procedia Environ. Sci. 2010, 2, 1510–1524. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Johengen, T.H.; Liebig, J.R. Feedback between zebra mussel selective feeding and algal composition affects mussel condition: Did the regime changer pay a price for its success? Freshw. Biol. 2009, 54, 47–63. [Google Scholar] [CrossRef]
- Winkel, E.H.T.; Davids, C. Food selection by Dreissena polymorpha Pallas (Mollusca: Bivalvia). Freshw. Biol. 1982, 12, 553–558. [Google Scholar] [CrossRef]
- Kimbrough, K.L.; Johnson, W.E.; Lauenstein, G.G.; Christensen, J.D.; Apeti, D.A. An Assessment of Two Decades of Contaminant Monitoring in the Nation’s Coastal Zone; NOAA Technical Memorandum NOS NCCOS 74; National Centers for Coastal Ocean Science: Silver Spring, MD, USA, 2008; 105p.
- Schöne, B.R.; Krause, R.A. Retrospective environmental biomonitoring—Mussel Watch expanded. Glob. Planet. Chang. 2016, 144, 228–251. [Google Scholar] [CrossRef]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The fate of the cyanobacterial toxin β-N-methylamino-l-alanine in freshwater mussels. Ecotoxicol. Environ. Saf. 2014, 101, 51–58. [Google Scholar] [CrossRef]
- Faassen, E.; Antoniou, M.; Beekman-Lukassen, W.; Blahova, L.; Chernova, E.; Christophoridis, C.; Combes, A.; Edwards, C.; Fastner, J.; Harmsen, J.; et al. A Collaborative Evaluation of LC-MS/MS Based Methods for BMAA Analysis: Soluble Bound BMAA Found to Be an Important Fraction. Mar. Drugs 2016, 14, 45. [Google Scholar] [CrossRef]
- Main, B.J.; Italiano, C.J.; Rodgers, K.J. Investigation of the interaction of β-methylamino-l-alanine with eukaryotic and prokaryotic proteins. Amino Acids 2017. [Google Scholar] [CrossRef]
- Faassen, E.J. Extraction and LC-MS/MS Analysis of Underivatised BMAA. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 439–446. [Google Scholar]
- Rosén, J.; Westerberg, E.; Schmiedt, S.; Hellenäs, K.E. BMAA detected as neither free nor protein bound amino acid in blue mussels. Toxicon 2016, 109, 45–50. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.L.; Downing, S.; Phelan, R.R.; Downing, T.G. Environmental modulation of microcystin and β-N-methylamino-l-alanine as a function of nitrogen availability. Toxicon 2014, 87, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Conroy, J.D.; Edwards, W.J.; Pontius, R.A.; Kane, D.D.; Zhang, H.; Shea, J.F.; Richey, J.N.; CULVER, D.A. Soluble nitrogen and phosphorus excretion of exotic freshwater mussels (Dreissena spp.): Potential impacts for nutrient remineralisation in western Lake Erie. Freshw. Biol. 2005, 50, 1146–1162. [Google Scholar] [CrossRef]
- Johengen, T.H.; Nalepa, T.F.; Fahnenstiel, G.L.; Goudy, G. Nutrient Changes in Saginaw Bay, Lake Huron, After the Establishment of the Zebra Mussel (Dreissena polymorpha). J. Great Lakes Res. 1995, 21, 449–464. [Google Scholar] [CrossRef]
- Effler, S.W.; Siegfried, C. Zebra Mussel (Dreissena polymorpha) Populations in the Seneca River, New York: Impact on Oxygen Resources. Environ. Sci. Technol. 1994, 28, 2216–2221. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.C. Bivalves. In Invertebrate Blood Cells; Ratcliffe, N.A., Rowley, A.F., Eds.; Academic Press: New York, NY, USA, 1981; p. 368. ISBN 0-12-582101-8. [Google Scholar]
- Mydlarz, L.D.; Jones, L.E.; Harvell, C.D. Innate immunity, environmental and disease ecology of marine and freshwater invertebrates. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 251–288. [Google Scholar] [CrossRef]
- Donaghy, L.; Lambert, C.; Choi, K.S.; Soudant, P. Hemocytes of the carpet shell clam (Ruditapes decussatus) and the Manila clam (Ruditapes philippinarum): Current knowledge and future prospects. Aquaculture 2009, 297, 10–24. [Google Scholar] [CrossRef]
- Gosling, E. Bivalve Molluscs; Gosling, E., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2007; ISBN 9780470995532. [Google Scholar]
- Marigómez, I.; Soto, M.; Cajaraville, M.P.; Angulo, E.; Giamberini, L. Cellular and subcellular distribution of metals in molluscs. Microsc. Res. Tech. 2002, 56, 358–392. [Google Scholar] [CrossRef]
- Wright, S.H. Multiple pathways for amino acid transport in Mytilus gill. J. Comp. Physiol. B 1985, 156, 259–267. [Google Scholar] [CrossRef]
- Manahan, D.T.; Wright, S.H.; Stephens, G.C.; Rice, M.A. Transport of Dissolved Amino Acids by the Mussel, Mytilus edulis: Demonstration of Net Uptake from Natural Seawater. Science 1982, 215, 1253–1255. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.A.; Stephens, G.C. Uptake and internal distribution of exogenously supplied amino acids in the Pacific oyster, Crassostrea gigas (Thunberg). Aquaculture 1987, 66, 19–31. [Google Scholar] [CrossRef]
- Weiss, J.H.; Choi, D.W. Beta-N-methylamino-L-alanine neurotoxicity: Requirement for bicarbonate as a cofactor. Science 1988, 241, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Lance, E.; Josso, C.; Dietrich, D.; Ernst, B.; Paty, C.; Senger, F.; Bormans, M.; Gérard, C. Histopathology and microcystin distribution in Lymnaea stagnalis (Gastropoda) following toxic cyanobacterial or dissolved microcystin-LR exposure. Aquat. Toxicol. 2010, 98, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.D. The Circulatory Systems of Gastropods and Bivalves. In The Mollusca; Saleuddin, A., Wilbur, K., Eds.; Elsevier: Amsterdam, The Netherlands, 1983; pp. 189–238. [Google Scholar]
- Robinson, W.E.; Ryan, D.K. Transport of cadmium and other metals in the blood of the bivalve mollusc Mercenaria mercenaria. Mar. Biol. 1988, 97, 101–109. [Google Scholar] [CrossRef]
- Ricciardi, K.L.; Poynton, H.C.; Duphily, B.J.; Blalock, B.J.; Robinson, W.E. Bioconcentration and depuration of14C-labeled 17α-ethinyl estradiol and 4-nonylphenol in individual organs of the marine bivalve Mytilus edulis L. Environ. Toxicol. Chem. 2016, 35, 863–873. [Google Scholar] [CrossRef]
- Van Onselen, R.; Venables, L.; van de Venter, M.; Downing, T.G. β-N-Methylamino-l-Alanine Toxicity in PC12: Excitotoxicity vs. Misincorporation. Neurotox. Res. 2018, 33, 15–23. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Otterstein, S.K.B.; Downing, S.; Downing, T.G.; Pflugmacher, S. Response of antioxidant and biotransformation systems of selected freshwater mussels (Dreissena polymorpha, Anadonta cygnea, Unio tumidus, and Corbicula javanicus) to the cyanobacterial neurotoxin β-N-methylamino-L-alanine. Toxicol. Environ. Chem. 2014, 96, 451–465. [Google Scholar] [CrossRef]
- Papadimitriou, T.; Kagalou, I.; Bacopoulos, V.; Leonardos, I.D. Accumulation of microcystins in water and fish tissues: An estimation of risks associated with microcystins in most of the Greek Lakes. Environ. Toxicol. 2009, 25, 418–427. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P. Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and -RR in two freshwater shrimps, Palaemon modestus and Macrobrachium nipponensis, from a large shallow, eutrophic lake of the subtropical China. Toxicon 2005, 45, 615–625. [Google Scholar] [CrossRef]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, Z.; Wang, Z. Oxidative stress responses and toxin accumulation in the freshwater snail Radix swinhoei (Gastropoda, Pulmonata) exposed to microcystin-LR. Environ. Sci. Pollut. Res. 2016, 23, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Rosén, J.; Westerberg, E.; Hellenäs, K.E.; Salomonsson, M.L. A new method for analysis of underivatized free β-methylamino-alanine: Validation and method comparison. Toxicon 2016, 121, 105–108. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepoutre, A.; Faassen, E.J.; Zweers, A.J.; Lürling, M.; Geffard, A.; Lance, E. How the Neurotoxin β-N-Methylamino-l-Alanine Accumulates in Bivalves: Distribution of the Different Accumulation Fractions among Organs. Toxins 2020, 12, 61. https://doi.org/10.3390/toxins12020061
Lepoutre A, Faassen EJ, Zweers AJ, Lürling M, Geffard A, Lance E. How the Neurotoxin β-N-Methylamino-l-Alanine Accumulates in Bivalves: Distribution of the Different Accumulation Fractions among Organs. Toxins. 2020; 12(2):61. https://doi.org/10.3390/toxins12020061
Chicago/Turabian StyleLepoutre, Alexandra, Elisabeth J. Faassen, A. J. Zweers, Miquel Lürling, Alain Geffard, and Emilie Lance. 2020. "How the Neurotoxin β-N-Methylamino-l-Alanine Accumulates in Bivalves: Distribution of the Different Accumulation Fractions among Organs" Toxins 12, no. 2: 61. https://doi.org/10.3390/toxins12020061
APA StyleLepoutre, A., Faassen, E. J., Zweers, A. J., Lürling, M., Geffard, A., & Lance, E. (2020). How the Neurotoxin β-N-Methylamino-l-Alanine Accumulates in Bivalves: Distribution of the Different Accumulation Fractions among Organs. Toxins, 12(2), 61. https://doi.org/10.3390/toxins12020061