Mass Occurrence of Anatoxin-a- and Dihydroanatoxin-a-Producing Tychonema sp. in Mesotrophic Reservoir Mandichosee (River Lech, Germany) as a Cause of Neurotoxicosis in Dogs

 ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Macroscopic and Microscopic Appearance of Tychonema
2.2. Animal Necroscopy and Histopathology
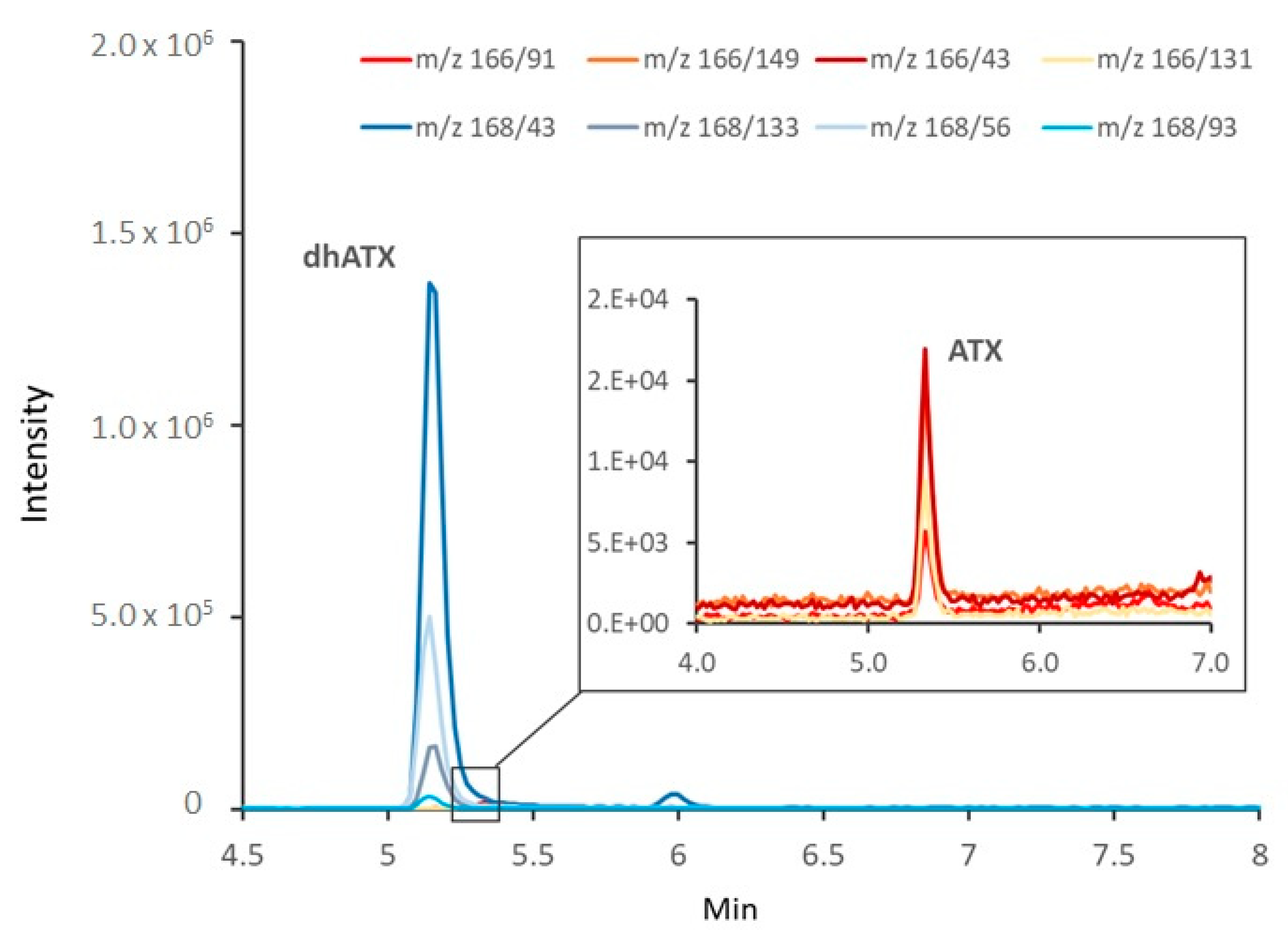
2.3. Toxin Analyses
2.4. Detection of Toxin Genes
2.5. Illumina MiSeq Sequencing
2.6. Hydro-Chemical and Hydro-Physical Characterization
3. Discussion
4. Conclusions
5. Materials and Methods
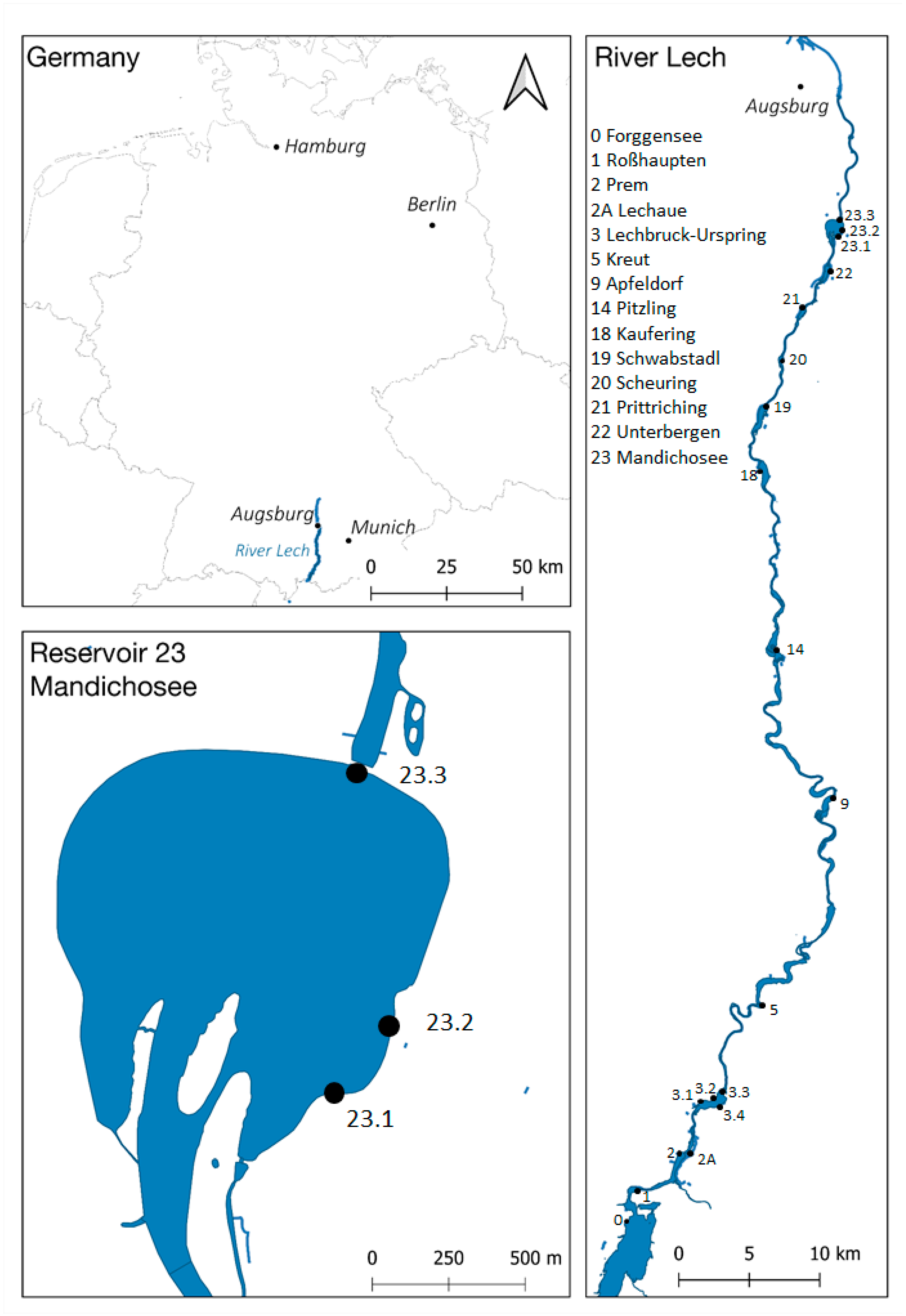
5.1. Study Site
5.2. Animal Necroscopy and Histopathology
5.3. Sampling and in Situ Measurements of Hydro-physical Parameters
5.4. Post-Processing of the Samples
5.5. Toxin Analyses
5.6. Microscopy
5.7. Molecular Approach
5.7.1. DNA Extraction and Purification
5.7.2. Polymerase Chain Reaction
5.7.3. Sequencing
5.8. Hydrochemical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carmichael, W.W. (Ed.) Freshwater blue-green algae (cyanobacteria) toxins—A review. In The Water Environment; Springer: Boston, MA, USA, 1981; pp. 1–13. [Google Scholar] [CrossRef]
- Sivonen, K. Cyanobacterial toxins. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Jakubowska, N.; Szeląg-Wasielewska, E. Toxic picoplanktonic cyanobacteria. Mar. Drugs 2015, 13, 1497–1518. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Niclas, E.; Antunes, A.; Gerwick, W.H.; Vasconcelos, V. The chemical ecology of cyanobacteria. Nat. Prod. Rep. 2012, 29, 372–391. [Google Scholar] [CrossRef] [PubMed]
- Morscheid, H.; Fromme, H.; Krause, D.; Kurmayer, R.; Morscheid, H.; Teuber, K. Toxinbildende Cyanobakterien (Blaualgen) in Bayerischen Gewässern; Bayerisches Landesamt für Umwelt: Augsburg, Germany, 2006; Volume 125, pp. 1–145. [Google Scholar]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Hall, N.S.; Wu, Y. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China. Environ. Sci. Technol. 2015, 49, 1051–1059. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria–ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Fastner, J.; Beulker, C.; Geiser, B.; Hoffmann, A.; Kröger, R.; Teske, K.; Hoppe, J.; Mundhenk, L.; Neurath, H.; Sagebiel, D.; et al. Fatal neurotoxicosis in dogs associated with tychoplanktic, anatoxin-a producing Tychonema sp. in mesotrophic Lake Tegel, Berlin. Toxins 2018, 10, 60. [Google Scholar] [CrossRef]
- Namikoshi, M.; Murakami, T.; Watanabe, M.; Oda, T.; Yamada, J.; Tsujimura, S.; Nagai, H.; Oishi, S. Simultaneous production of homoanatoxin-a, anatoxin-a, and a new non-toxic 4-hydroxyhomoanatoxin-a by the cyanobacterium Raphidiopsis mediterranea Skuja. Toxicon 2003, 42, 533–538. [Google Scholar] [CrossRef]
- Aráoz, R.; Nghiem, H.; Rippka, R.; Palibroda, N.; Tandeau de Marsac, N.; Herdman, M. Neurotoxins in axenic oscillatorian cyanobacteria: Coexistence of anatoxin-a and homoanatoxin-a determined by ligand-binding assay and GC/MS. Microbiology 2005, 151, 1263–1273. [Google Scholar] [CrossRef]
- Méjean, A.; Mann, S.; Maldiney, T.; Vassiliadis, G.; Lequin, O.; Ploux, O. Evidence that Biosynthesis of the Neurotoxic Alkaloids Anatoxin-a and Homoanatoxin-a in the Cyanobacterium Oscillatoria PCC 6506 Occurs on a Modular Polyketide Synthase Initiated by l-Proline. J. Am. Chem. Soc. 2009, 131, 7512–7513. [Google Scholar] [CrossRef] [PubMed]
- Méjean, A.; Paci, G.; Gautier, V.; Ploux, O. Biosynthesis of anatoxin-a and analogues (anatoxins) in cyanobacteria. Toxicon 2014, 91, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Heath, M.W.; Kuhajek, J.; Ryan, K.G. Fine-scale spatial variability in anatoxin-a and homoanatoxin-a concentrations in benthic cyanobacterial mats: Implication for monitoring and management. J. Appl. Microbiol. 2010, 109, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Smith, F.M.J.; Heath, M.W.; Palfroy, T.; Gaw, S.; Young, R.G.; Ryan, K.G. Within-Mat Variability in Anatoxin-a and Homoanatoxin-a Production among Benthic Phormidium (Cyanobacteria) Strains. Toxins 2012, 4, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Biessy, L.; Puddick, J. Anatoxins are consistently released into the water of streams with Microcoleus autumnalis-dominated (cyanobacteria) proliferations. Harmful Algae 2018, 80, 88–95. [Google Scholar] [CrossRef]
- Rantala-Ylinen, A.; Känä, S.; Wang, H.; Rouhiainen, L.; Wahlsten, M.; Rizzi, E.; Berg, K.; Gugger, M.; Sivonen, K. Anatoxin-a synthetase gene cluster of the cyanobacterium Anabaena sp. strain 37 and molecular methods to detect potential producers. Appl. Environ. Microbiol. 2011, 77, 7271–7278. [Google Scholar] [CrossRef]
- Faassen, E.; Harkema, L.; Begeman, L.; Lurling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef]
- Schneider, S.; Melzer, A. The Trophic Index of Macrophytes (TIM)–a new tool for indicating the trophic state of running waters. Int. Rev. Hydrobiol. 2003, 88, 49–67. [Google Scholar] [CrossRef]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota-Oscillatoriales. In Freshwater Flora of Central Europe; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Heidelberg, Germany, 2005; pp. 369–375. [Google Scholar]
- Conklin, K.Y.; Stancheva, R.; Otten, T.G.; Fadness, R.; Boyer, G.L.; Read, R.; Zhang, X.; Sheath, R.G. Molecular and morphological characterization of a novel dihydroanatoxin-a producing Microcoleus species (cyanobacteria) from the Russian River, California, USA. Harmful Algae 2020, 93, 101767. [Google Scholar] [CrossRef] [PubMed]
- Castenholtz, R.W.; Rippka, R.; Herdmann, M. Form-Tychonema. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; Wiley: Hoboken, NJ, USA, 2015; p. 561. [Google Scholar] [CrossRef]
- Shams, S.; Capelli, C.; Cerasino, L.; Ballot, A.; Dietrich, D.R.; Sivonen, K.; Salmaso, N. Anatoxin-a producing Tychonema (Cyanobacteria) in European waterbodies. Water Res. 2015, 69, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, N.; Cerasino, L.; Boscaini, A.; Capelli, C. Planktic Tychonema (Cyanobacteria) in the large lakes south of the Alps: Phylogenetic assessment and toxigenic potential. FEMS Microbiol. Ecol. 2016, 92, 155. [Google Scholar] [CrossRef]
- Capelli, C.; Cerasino, L.; Boscaini, A.; Salmaso, N. Molecular tools for the quantitative evaluation of potentially toxigenic Tychonema bourrellyi (Cyanobacteria, Oscillatoriales) in large lakes. Hydrobiologia 2018, 824, 109–119. [Google Scholar] [CrossRef]
- Komárek, J.; Komárkova, J.; Kling, H. Filamentous Cyanobacteria. In Freshwater Algae of North America; Wehr, J.D., Sheat, R.G., Eds.; Elsevier: Waltham, MA, USA, 2003; pp. 117–196. [Google Scholar]
- Butterwick, C.; Heaney, S.I.; Talling, J.F. Diversity in the influence of temperature on the growth rates of freshwater algae, and its ecological relevance. Freshw. Biol. 2005, 50, 291–300. [Google Scholar] [CrossRef]
- James, K.J.; Furey, A.; Sherlock, I.R.; Stack, M.A.; Twohig, M.; Caudwell, F.B.; Skulberg, O.M. Sensitive determination of anatoxin-a, homoanatoxin-a and their degradation products by liquid chromatography with fluorimetric detection. J. Chromatogr. 1998, 798, 147–157. [Google Scholar] [CrossRef]
- Mann, S.; Cohen, M.; Chapuis-Hugon, F.; Pichon, V.; Mazmouz, R.; Méjean, A. Synthesis, configuration assignment, and simultaneous quantification by liquid chromatography coupled to tandem mass spectrometry, of dihydroanatoxin-a and dihydrohomoanatoxin-a together with the parent toxins, in axenic cyanobacterial strains and in environmental samples. Toxicon 2012, 60, 1404–1414. [Google Scholar] [CrossRef]
- Méjean, A.; Dalle, K.; Paci, G.; Bouchonnet, S.; Mann, S.; Pichon, V.; Ploux, O. Dihydroanatoxin-a is biosynthesized from Proline in Cylindrospermum stagnale PCC 7417: Isotopic incorporation experiments and mass spectrometry analysis. J. Nat. Prod. 2016, 79, 1775–1782. [Google Scholar] [CrossRef]
- Wood, S.A.; Puddick, J.; Fleming, R.; Heussner, A.H. Detection of anatoxin-producing Phormidium in a New Zealand farm pond and an associated dog death. N. Z. J. Bot. 2017, 55, 36–46. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Bates, H.A.; Rapoport, H. Synthesis of anatoxin-a via intramolecular cyclization of iminium salts. J. Am. Chem. Soc. 1979, 101, 1259–1265. [Google Scholar] [CrossRef]
- Wonnacott, S.; Jackman, S.; Swanson, K.L.; Rapoport, H.; Albuquerque, E.X. Nicotinic pharmacology of anatoxin analogs. Side chain structure-activity relationships at neuronal nicotinic ligand binding sites. J. Pharmacol. Exp. Ther. 1991, 259, 387–391. [Google Scholar] [PubMed]
- Puddick, J.; van Ginkel, R.; Page, C.D.; Murray, J.S.; Greenhough, H.E.; Bowater, J.; Selwood, A.I.; Wood, S.A.; Prinsep, M.R.; Truman, P.; et al. Acute toxicity of dihydroanatoxin-a from Microcoleus autumnalis in comparison to anatoxin-a. Chemosphere 2020, 263, 127937. [Google Scholar] [CrossRef] [PubMed]
- Toporowska, M.; Pawlik-Skowrońska, B.; Kalinowska, R. Accumulation and effects of cyanobacterial microcystins and anatoxin-a on benthic larvae of Chironomus spp. (Diptera: Chironomidae). Eur. J. Entomol. 2014, 111, 83–90. [Google Scholar] [CrossRef]
- Kelly, L.T.; Puddick, J.; Ryan, K.G.; Champeau, O.; Wood, S.A. An ecotoxicological assessment of the acute toxicity of anatoxin congeners on New Zealand Deleatidium species (mayflies). Inland Waters 2019, 10, 101–108. [Google Scholar] [CrossRef]
- Osswald, J.; Azevedo, J.; Vasconcelos, V.; Guilhermino, L. Experimental determination of the bioconcentration factors for anatoxin-a in juvenile rainbow trout (Oncorhynchus mykiss). Proc. Int. Acad. Ecol. Environ. Sci. 2017, 1, 77. [Google Scholar]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef]
- Wood, S.A.; Hamilton, D.P.; Paul, W.J.; Safi, K.A.; Williamson, W.M. New Zealand Guidelines for Cyanobacteria in Recreational Fresh Waters: INTERIM Guidelines (Commissioned Report for External Body); Ministry for the Environment: Wellington, New Zealand, 2009; pp. 1–82.
- Chorus, I.; Schauser, I. Oligotrophication of Lake Tegel and Schlachtensee, Berlin—Analysis of System Components, Causalities and Response Thresholds Compared to Responses of Other Waterbodies; Umweltbundesamt: Dessau-Roßlau, Germany, 2011; pp. 1–157.
- James, K.J.; Crowley, J.; Hamilton, B.; Lehane, M.; Skulberg, O.; Furey, A. Anatoxins and degradation products, determined using hybrid quadrupole time-of-flight and quadrupole ion-trap mass spectrometry: Forensic investigations of cyanobacterial neurotoxin poisoning. Rapid Commun. Mass Spectrom. 2005, 19, 1167–1175. [Google Scholar] [CrossRef]
- Zwirglmaier, K.; Keiz, K.; Engel, M.; Geist, J.; Raeder, U. Seasonal and spatial patterns of microbial diversity along a trophic gradient in the interconnected lakes of the Osterseen Lake District, Bavaria. Front. Microbiol. 2015, 6, 1168. [Google Scholar] [CrossRef]
- Rantala, A.; Rajaniemi-Wacklin, P.; Lyra, C.; Lepistö, L.; Rintala, J.; Mankiewicz-Boczek, J.; Sivonen, K. Detection of microcystin-producing cyanobacteria in Finnish lakes with genus-specific microcystin synthetase gene E (mcyE) PCR and associations with environmental factors. Appl. Environ. Microbiol. 2006, 72, 6101–6110. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi-Man, O.; Davenport, E.R.; Gilad, Y. Taxonomic classification of bacterial 16S rRNA genes using short sequencing reads: Evaluation of effective study designs. PLoS ONE 2013, 8, 53608. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Joseph, D.; Kapfhammer, M.; Giritli, S.; Horn, M.; Haller, D.; Clavel, T. IMNGS: A comprehensive open resource of processed 16S rRNA microbial profiles for ecology and diversity studies. Sci. Rep 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Deutsche Einheitsverfahren zur Wasser-, Abwasser-und Schlammuntersuchung; Wasserchemische Gesellschaft in der Gesellschaft Deutscher Chemiker; Wiley-VCH: Weinheim, Germany, 2013. [CrossRef]
- Navone, R. Proposed method for nitrate in potable waters. J. Am. Water Works Assoc. 1964, 56, 781–784. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample Origin (Site) | Sample | Date (Day.Month.Year) | ATX (µg/L) | DhATX (µg/L) |
|---|---|---|---|---|
| Dog (D3) | Stomach content | 14 August 19 | n.d. | 563 |
| Dog (D1) | Stomach content | 14 August 19 | 25.2 | 1182 |
| Reservoir Mandicho (23.1) | Mat from shoreline | 21 August 19 | 24.4–320 | 1917–39,528 |
| Reservoir Mandicho (23.1) | Floating mat | 21 August 19 | 241–453 | 16,954–67,622 |
| Reservoir Mandicho (23.2) | Mat from shoreline | 21 August 19 | 28.2–214 | 2375–23,488 |
| Reservoir Mandicho (23.2) | Water and macrophytes | 14 August 19 | 9.9–40.9 | 607–3012 |
| Reservoir Mandicho (23.2) | Elodea with water | 21 August 19 | 10.3–12.9 | 320–326 |
| Reservoir Mandicho (23.2) | Macrophytes with water | 21 August 19 | n.d. | 0.7 |
| Reservoir Mandicho (23.2) | Myriophyllum with water | 21 August 19 | 3.5 | 157 |
| Reservoir Mandicho (23.1) | Macrophytes/Periphyton | 23 August 19 | 7.7–272 | 3113–29,335 |
| Reservoir Mandicho (23.1) | Macrophytes and water | 17 September 19 | 2.1 | 310 |
| Reservoir Mandicho (23.1) | Macrophytes and water | 17 September 19 | 0.1 | 0.2 |
| Reservoir Mandicho (23.2) | Water surface above mat | 21 August 19 | <LOQ | 2.7 |
| Reservoir Mandicho (23.2) | Water surface 20 cm above mat | 21 August 19 | <LOQ | 1.0 |
| Reservoir Mandicho (23.2) | Water surface 50 cm above mat | 21 August 19 | <LOQ | 1.4 |
| Reservoir Mandicho (23.2) | Water surface 100 cm above mat | 21 August 19 | <LOQ | 5.0 |
| Reservoir Mandicho (23.2) | Water | 14 August 19 | <LOQ | 3.8 |
| Reservoir Mandicho (23.1) | Water | 21 August 19 | n.d. | 0.6 |
| Reservoir Prittriching (21) | Water | 17 September 19 | n.d. | n.d. |
| Reservoir Scheuring (20) | Water | 17 September 19 | n.d. | n.d. |
| Reservoir Schwabstadl (19) | Water | 17 September 19 | n.d. | 0.7 |
| Reservoir Kaufering (18) | Water | 17 September 19 | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, F.; Fastner, J.; Bartha-Dima, B.; Breuer, W.; Falkenau, A.; Mayer, C.; Raeder, U. Mass Occurrence of Anatoxin-a- and Dihydroanatoxin-a-Producing Tychonema sp. in Mesotrophic Reservoir Mandichosee (River Lech, Germany) as a Cause of Neurotoxicosis in Dogs. Toxins 2020, 12, 726. https://doi.org/10.3390/toxins12110726
Bauer F, Fastner J, Bartha-Dima B, Breuer W, Falkenau A, Mayer C, Raeder U. Mass Occurrence of Anatoxin-a- and Dihydroanatoxin-a-Producing Tychonema sp. in Mesotrophic Reservoir Mandichosee (River Lech, Germany) as a Cause of Neurotoxicosis in Dogs. Toxins. 2020; 12(11):726. https://doi.org/10.3390/toxins12110726
Chicago/Turabian StyleBauer, Franziska, Jutta Fastner, Bernadett Bartha-Dima, Wolfram Breuer, Almuth Falkenau, Christian Mayer, and Uta Raeder. 2020. "Mass Occurrence of Anatoxin-a- and Dihydroanatoxin-a-Producing Tychonema sp. in Mesotrophic Reservoir Mandichosee (River Lech, Germany) as a Cause of Neurotoxicosis in Dogs" Toxins 12, no. 11: 726. https://doi.org/10.3390/toxins12110726
APA StyleBauer, F., Fastner, J., Bartha-Dima, B., Breuer, W., Falkenau, A., Mayer, C., & Raeder, U. (2020). Mass Occurrence of Anatoxin-a- and Dihydroanatoxin-a-Producing Tychonema sp. in Mesotrophic Reservoir Mandichosee (River Lech, Germany) as a Cause of Neurotoxicosis in Dogs. Toxins, 12(11), 726. https://doi.org/10.3390/toxins12110726