Gastric Ulceration and Immune Suppression in Weaned Piglets Associated with Feed-Borne Bacillus cereus and Aspergillus fumigatus



Abstract
1. Introduction
2. Results
2.1. B. cereus and B. subtilis Were Highly Contaminated in Swine Feed
2.2. Increasing DON Production Post Fermentation with B. cereus Isolates
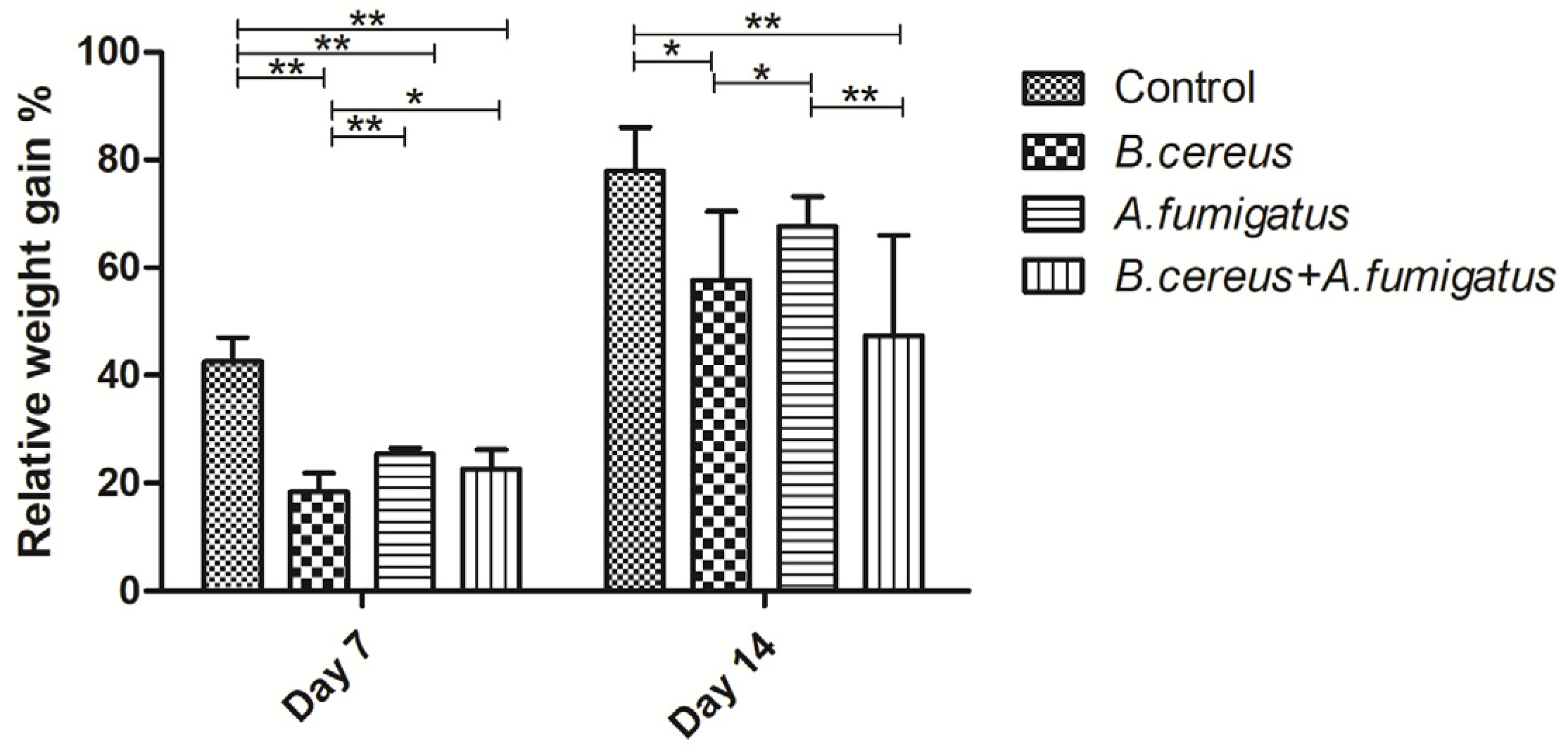
2.3. B. cereus, A. fumigatus and co-infection Exacerbated Diarrhea and Piglets’ Growth
2.4. B. cereus and A. fumigatus Reduced CSFV-, FMD- and PED-Specific Antibodies
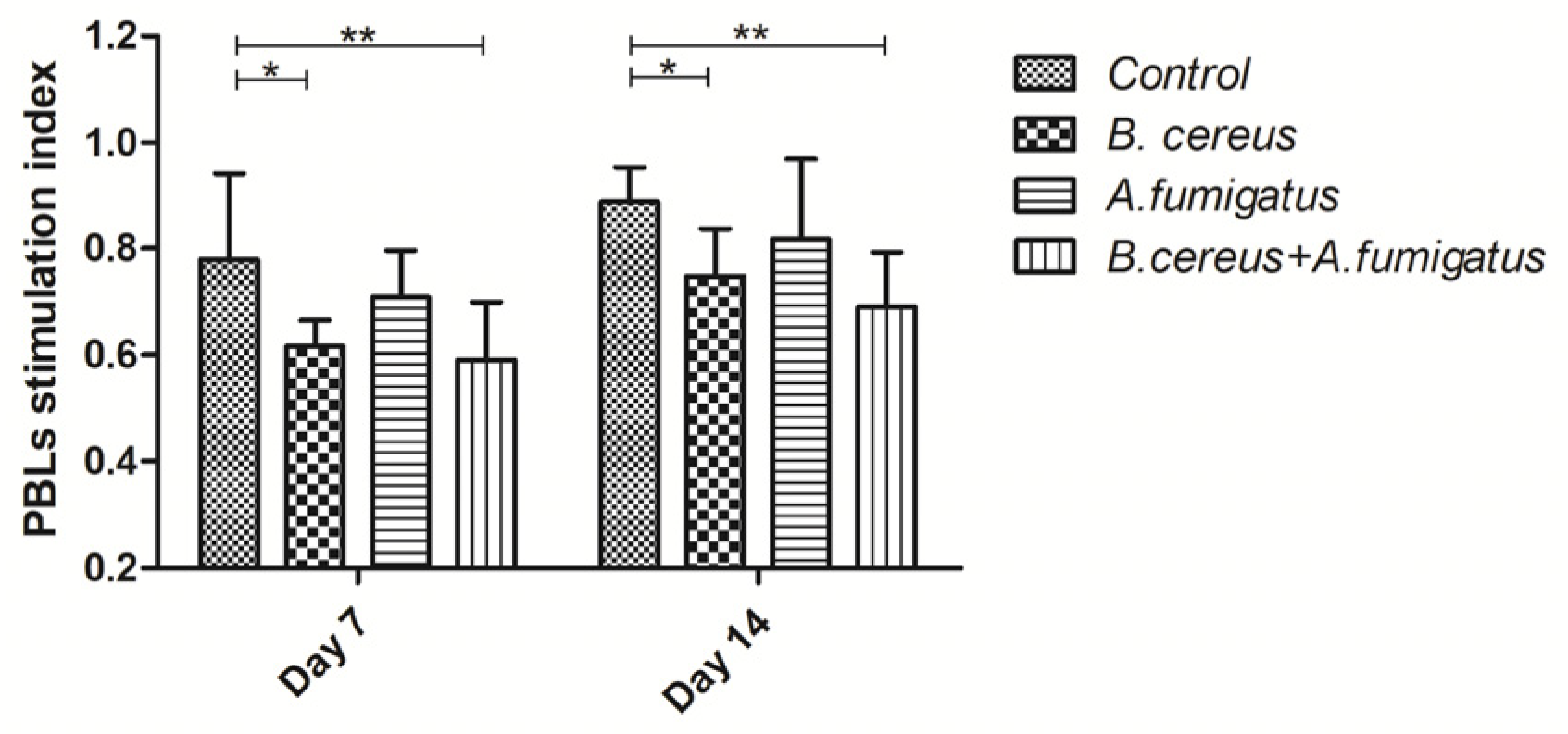
2.5. B. cereus and Co-Infection Reduced Lymphocyte Proliferation
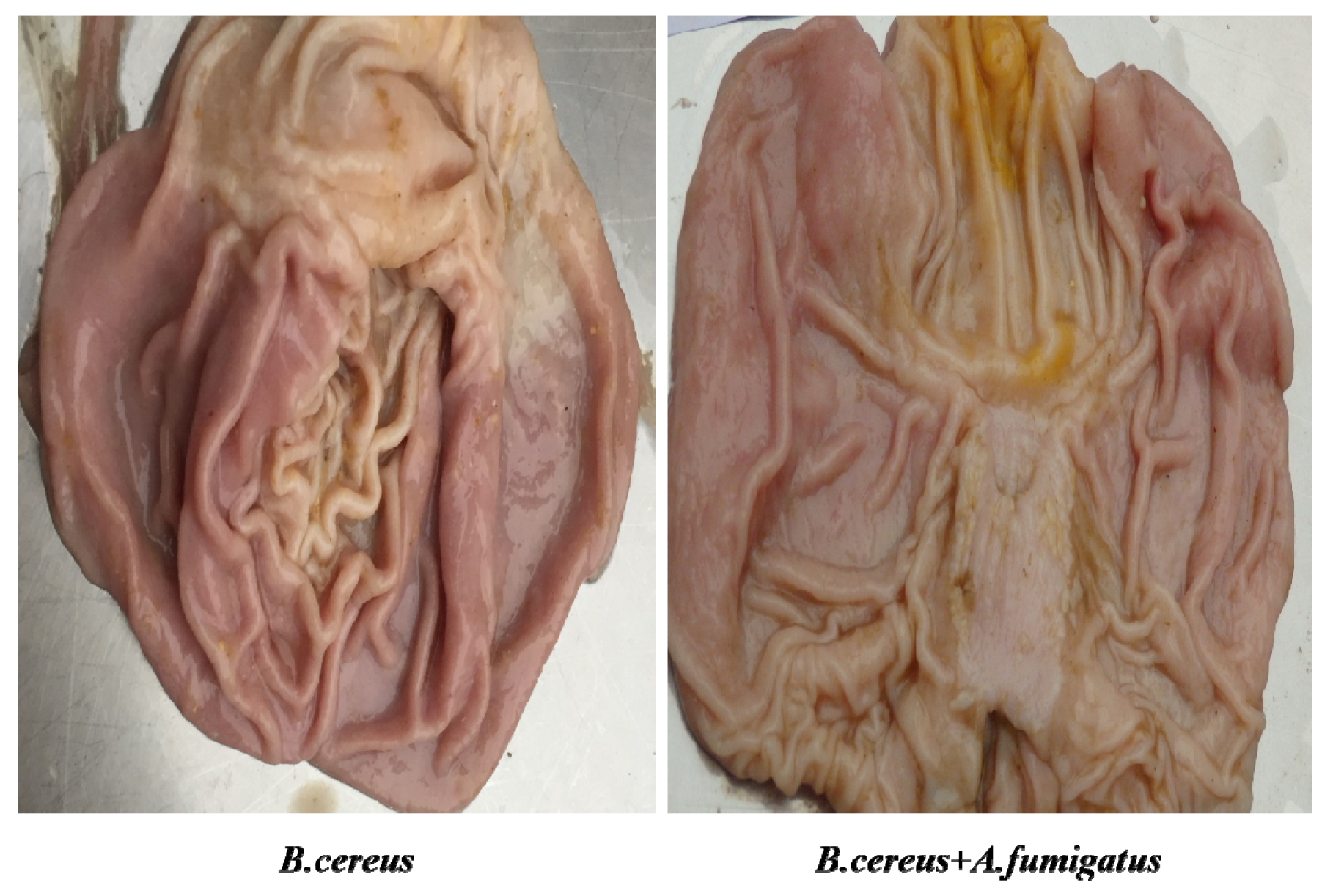
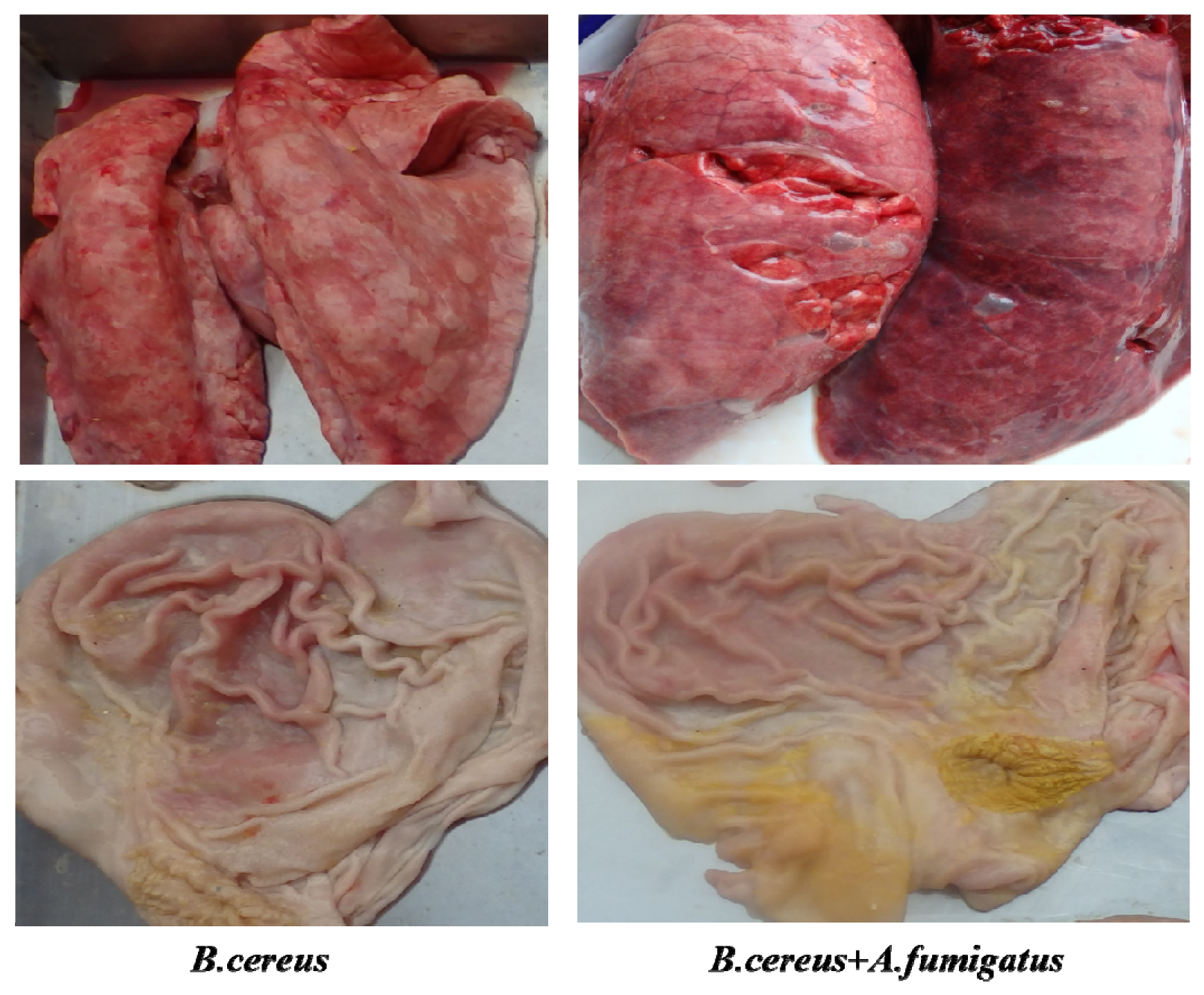
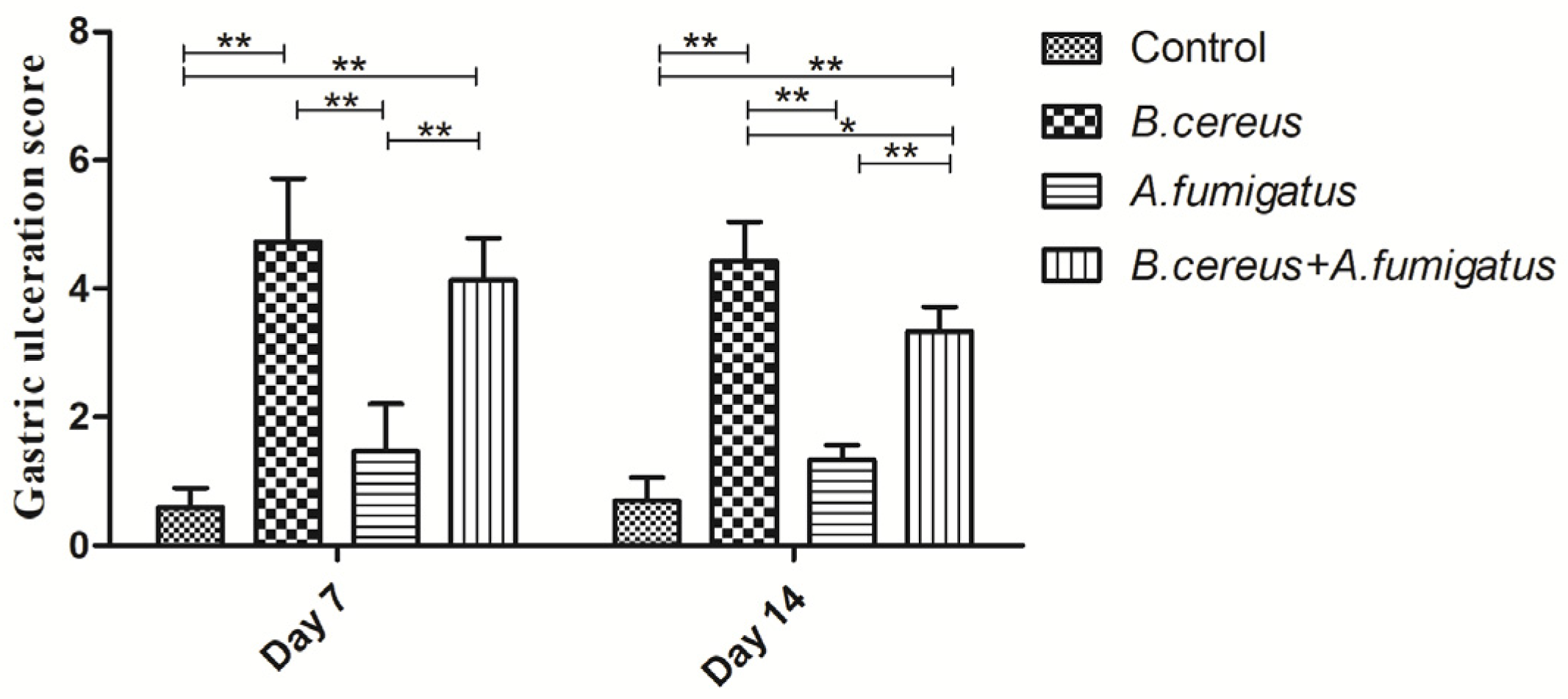
2.6. Both B. cereus and co-infection Induced Typic Gastric Ulcerations in Piglets
2.7. B. cereus and co-infection Induced DON Production in Piglets’ Gastric Ulcers
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bacillus Spp Isolation, Identification and Contamination Status
5.2. Determination of Deoxynivalenol (DON) Post Fermentation with Bacillus cereus Isolates
5.3. Animals and Ethics Statement
5.4. Detecting Antibodies against CSFV, PED and FMD
5.5. Lymphocyte Proliferation Assay
5.6. Pathological Evaluation
5.7. Quantitative DON in Stomach
5.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Amory, J.R.; Mackenzie, A.M.; Pearce, G.P. Factors in the housing environment of finisher pigs associated with the development of gastric ulcers of the article. Vet. Rec. 2006, 158, 260–264. [Google Scholar] [CrossRef]
- Friendship, R.M. Gastric ulcers. In Diseases of Swine, 9th ed.; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J., Eds.; Blackwell Professional Publishing: Ames, IA, USA, 2006; pp. 891–899. [Google Scholar]
- Gottardo, F.; Scollo, A.; Contiero, B.; Bottacini, M.; Mazzoni, C.; Edwards, S.A. Prevalence and risk factors for gastric ulceration in pigs slaughtered at 170 kg of the article. Animal 2017, 11, 2010–2018. [Google Scholar] [CrossRef]
- Tsilia, V.; Uyttendaele, M.; Kerckhof, F.M.; Rajkovic, A.; Heyndrickx, M.; van de Wiele, T. Bacillus cereus adhesion to simulated intestinal mucus is determined by its growth on mucin, rather than intestinal environmental parameters of the article. Foodborne Pathog. Dis. 2015, 12, 904–913. [Google Scholar] [CrossRef]
- Lund, T.; De Buyser, M.-L.; Granum, P.E. A new cytotoxin from bacillus cereus that may cause necrotic enteritis of the article. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef]
- Dietrich, R.; Moravek, M.; Burk, C.; Granum, P.E.; Martlbauer, E. Production and characterization of antibodies against each of the three subunits of the bacillus cereus nonhemolytic enterotoxin complex of the article. Appl. Environ. Microbiol. 2005, 71, 8214–8220. [Google Scholar] [CrossRef]
- Bottone, J.E. Bacillus cereus, a volatile human pathogen of the article. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef]
- Jessberger, N.; Dietrich, R.; Bock, S.; Didier, A.; Martlbauer, E. Bacillus cereus enterotoxins act as major virulence factors and exhibit distinct cytotoxicity to different human cell lines of the article. Toxicon 2014, 77, 49–57. [Google Scholar] [CrossRef]
- Sequoia, T.W.J. Probiotics for gastrointestinal conditions: A summary of the evidence of the article. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar]
- Kota, R.; Ambati, R.; Yalakurthi, A.; Srirama, K.; Reddy, P. Recent advances in probiotics as live biotherapeutics against gastrointestinal diseases of the article. Curr. Med. Chem. 2018, 24, 3162–3171. [Google Scholar]
- Haque, M.A.; Quan, H.; Zuo, Z.; Khan, A.; Siddique, N.; He, C. Pathogenicity of feed-borne bacillus cereus and its implication on food safety. Agrobiol. Rec. 2020, 3, 1–16, 81–85. [Google Scholar]
- Cui, Y.; Liu, X.; Dietrich, R.; Martlbauer, E.; Cao, J.; Ding, S.; Zhu, K. Characterization of bacillus cereus isolates from local dairy farms in china of the article. FEMS Microbiol. Lett. 2016, 363, fnw096. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Hu, Q.; Xu, F.; Ding, S.Y.; Zhu, K. Characterization of bacillus cereus in dairy products in china of the article. Toxins 2020, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Yang, Q.; He, F.; Lan, R.; Hao, J.; Ni, P.; Liu, Y.; Li, R. National safety survey of animal-use commercial probiotics and their spillover effects from farm to humans: An emerging threat to public health of the article. Clin. Infect. Dis. 2020, 70, 2386–2395. [Google Scholar] [CrossRef]
- Cordonnier, C.; Ribaud, P.; Herbrecht, R.; Milpied, N.; Valteau-Couanet, D.; Morgan, C.; Wade, A.; Cellulaire, S.F.d.G.d.M.e.d.T. Prognostic factors for death due to invasive aspergillosis after hematopoietic stem cell transplantation: A 1-year retrospective study of consecutive patients at french transplantation centers of the article. Aspergillosis Stem Cell Transplant. 2006, 42, 955–963. [Google Scholar] [CrossRef]
- Farnell, E.; Rousseau, K.; Thornton, D.J.; Bowyer, P.; Herrick, S.E. Expression and secretion of aspergillus fumigatus proteases are regulated in response to different protein substrates of the article. Fungal Biol. 2012, 116, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- de Oca, V.M.; Valdes, S.E.; Segundo, C.; Gomez, G.G.; Ramirez, J.; Cervantes, R.A. Aspergillosis, a natural infection in poultry: Mycological and molecular characterization and determination of gliotoxin in aspergillus fumigatus isolates of the article. Avian Dis. 2017, 61, 77–82. [Google Scholar] [CrossRef]
- Chu, J.; Zhang, Q.; Zuo, Z.; El-Ashram, S.; Guo, Y.; Zhao, P.; Huang, S.; He, C.; Khan, A. Co-infection of chlamydia psittaci with h9n2, ort and aspergillus fumigatus contributes to severe pneumonia and high mortality in spf chickens of the article. Sci. Rep. 2017, 7, 13997. [Google Scholar] [CrossRef]
- Zhang, Q.; Zuo, Z.; Guo, Y.; Zhang, T.; Han, Z.; Huang, S.; Karama, M.; Saleemi, M.K.; Khan, A.; He, C. Contaminated feed-borne bacillus cereus aggravates respiratory distress post avian influenza virus h9n2 infection by inducing pneumonia of the article. Sci. Rep. 2019, 9, 7231. [Google Scholar] [CrossRef]
- De Witte, C.; Ducatelle, R.; Haesebrouck, F. The role of infectious agents in the development of porcine gastric ulceration of the article. Vet. J. 2018, 236, 56–61. [Google Scholar] [CrossRef]
- Almeida, L.R.; Costa, P.S.; Nascimento, A.M.A.; Reis, M.d.P.; Barros, K.O.; Alvim, L.B.; Nunes, Á.C.; Queiroz, D.M.M.; Rocha, G.A.; Nicoli, J.R. Porcine stomachs with and without gastric ulcer differ in lactobacillus load and strain characteristics of the article. Can. J. Microbiol. 2018, 64, 493–499. [Google Scholar] [CrossRef]
- De Witte, C.; Flahou, B.; Ducatelle, R.; Smet, A.; De Bruyne, E.; Cnockaert, M.; Taminiau, B.; Daube, G.; Vandamme, P.; Haesebrouck, F. Detection, isolation and characterization of fusobacterium gastrosuis sp. Nov. Colonizing the stomach of pigs of the article. Syst. Appl. Microbiol. 2017, 40, 42–50. [Google Scholar] [CrossRef]
- De Witte, C.; Demeyere, K.; De Bruyckere, S.; Taminiau, B.; Daube, G.; Ducatelle, R.; Meyer, E.; Haesebrouck, F. Characterization of the non-glandular gastric region microbiota in helicobacter suis-infected versus non-infected pigs identifies a potential role for fusobacterium gastrosuis in gastric ulceration of the article. Vet. Res. 2019, 50, 39. [Google Scholar] [CrossRef] [PubMed]
- Krakowka, S.E.K.; Rings, D.M.; Argenzio, R.A. Production of gastroesophageal erosions and ulcers (geu) in gnotobiotic swine monoinfected with fermentative commensal bacteria and fed high-carbohydrate diet of the article. Vet. Pathol. 1998, 35, 274–282. [Google Scholar] [CrossRef]
- Thomson, J.R.; Friendship, R.M. Digestive system. In Diseases of Swine, 10th ed.; Zimmerman, J., Karriker, L., Ramirez, A., Schwartz, K., Stevenson, G., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2012; pp. 199–226. [Google Scholar]
- Zuo, Z.; Li, Q.; Guo, Y.; Li, X.; Huang, S.; Hegemann, J.H.; He, C. Feed-borne bacillus cereus exacerbates respiratory distress in chickens infected with chlamydia psittaci by inducing haemorrhagic pneumonia of the article. Avian Pathol. 2020, 49, 251–260. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Koehler, T.M.; Lereclus, D. The bacillus cereus group: Bacillus species with pathogenic potential of the article. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Mccormick, S. The role of don in pathogenicity of the article. In Fusarium Head Blight of Wheat & Barley Name; Alberta Ag-Info Centre: Red Deer, AB, Canada, 2003. [Google Scholar]
- Ramarao, N.; Lereclus, D. The inha1 metalloprotease allows spores of the b. Cereus group to escape macrophages of the article. Cell Microbiol. 2005, 7, 1357–1364. [Google Scholar] [CrossRef]
- Kroll, M.; Seisdedos, F.A.; Bachelerie, F.o.; Thomas, D.; Friguet, B.; Conconi, M. The secondary fungal metabolite gliotoxin targets proteolytic activities of the proteasome of the article. Chem. Biol. 1999, 6, 689–698. [Google Scholar] [CrossRef]
- Tallent, S.M.; Hait, J.M.; Ferguson, M. Comparative study of tempo bc automated mpn for the enumeration of bacillus cereus group in food of the article. J. Food Saf. 2018, 102, 1080–1090. [Google Scholar]
- Tallent, S.M.; Kotewicz, K.M.; Strain, E.; Bennett, R.W. Efficient isolation and identification of bacillus cereus group of the article. J. AOAC Int. 2012, 95, 446–451. [Google Scholar] [CrossRef]
- Olfert, E.D.; Godson, D.L. Humane endpoints for infectious disease animal models of the article. Ilar J. 2000, 41, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Baustad, B.; Nafstad, I. Gastric ulcers in swine. Effects of dietary particle size and crude fiber contents on ulceration of the article. Pathol. Vet. 1969, 6, 546–556. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feedstuffs | Samples | Positive | |||
|---|---|---|---|---|---|
| B. cereus | B. subtitis | B. licheniformis | B. thuringiensis | ||
| Started meals | 34 | 18(52.9) | 9 (26.5) | 5 (14.7) | 2 (5.9) |
| Growing meals | 36 | 23 (63.9) | 8 (22.2) | 3 (8.3) | 2 (5.6) |
| Finished meals | 35 | 22(62.9) | 6 (17.1) | 4 (11.4) | 3 (8.6) |
| Sow’s meals | 34 | 15 (44.1) | 10 (29.4) | 7 (20.6) | 2 (5.9) |
| Total | 139 | 78 (56.1) | 33 (23.7) | 19 (13.7) | 9 (6.5) |
| Feedstuffs | Samples | B. cereus (CFU/g) | |||
|---|---|---|---|---|---|
| <10 | 10–1000 | 1000–10,000 | >104 | ||
| Started meals | 10 | 1 (10.0) | 1 (10.0) | 3 (30.0) | 6 (60.0) |
| Growing meals | 11 | 0 (0.0) | 1 (9.1) | 3 (27.3) | 7 (63.6) |
| Finished meals | 12 | 0 (0.0) | 1 (8.3) | 3 (25.0) | 8 (66.7) |
| Sow meals | 10 | 1 (2.3) | 1 (10.0) | 3 (30.0) | 6 (60.0) |
| Totals | 43 | 1 (2.3) | 4 (9.3) | 12 (27.9) | 27 (62.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, Q.; Wang, Y.; Han, Z.; Qu, G.; Shen, Z.; Huang, S.; He, C. Gastric Ulceration and Immune Suppression in Weaned Piglets Associated with Feed-Borne Bacillus cereus and Aspergillus fumigatus. Toxins 2020, 12, 703. https://doi.org/10.3390/toxins12110703
Li X, Li Q, Wang Y, Han Z, Qu G, Shen Z, Huang S, He C. Gastric Ulceration and Immune Suppression in Weaned Piglets Associated with Feed-Borne Bacillus cereus and Aspergillus fumigatus. Toxins. 2020; 12(11):703. https://doi.org/10.3390/toxins12110703
Chicago/Turabian StyleLi, Xiaohui, Qiang Li, Yihui Wang, Zhenhai Han, Guanggang Qu, Zhiqiang Shen, Shujian Huang, and Cheng He. 2020. "Gastric Ulceration and Immune Suppression in Weaned Piglets Associated with Feed-Borne Bacillus cereus and Aspergillus fumigatus" Toxins 12, no. 11: 703. https://doi.org/10.3390/toxins12110703
APA StyleLi, X., Li, Q., Wang, Y., Han, Z., Qu, G., Shen, Z., Huang, S., & He, C. (2020). Gastric Ulceration and Immune Suppression in Weaned Piglets Associated with Feed-Borne Bacillus cereus and Aspergillus fumigatus. Toxins, 12(11), 703. https://doi.org/10.3390/toxins12110703