A Review of Nephrotoxicity of Microcystins


 and
and
Abstract
1. Introduction
2. Distribution and Metabolism of MCs in the Kidney
2.1. Distribution of MCs in the Kidney
2.2. Metabolism of MCs in the Kidney
3. Nephrotoxicity Caused by MCs on Mammals
3.1. Evidences from Epidemiological Investigation
3.2. Evidences from in Vivo Studies
3.2.1. Microcystis Extracts (MCEs)
3.2.2. Pure Microcystins (-LR)
3.3. Evidences from in Vitro Studies
4. Nephrotoxicity Caused by MCs on Fishes
4.1. Microcystis Extracts (MCEs)
4.2. Pure Microcystins
5. Potential Mechanisms of Action
5.1. Cellular Uptake of MCs
5.2. Modulation of PP2A Activities
5.3. Oxidative Stress
5.4. Cytoskeleton Disruption
5.5. DNA Damage
5.6. Apoptosis
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Boopathi, T.K.J. Impact of Environmental Factors on the Regulation of Cyanotoxin Production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef] [PubMed]
- Alosman, M.; Cao, L.; Massey, I.Y.; Yang, F. The lethal effects and determinants of microcystin-LR on heart: A mini review. Toxin Rev. 2020, 1–10. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The cyanotoxin-microcystins: Current overview. Rev. Environ. Sci. Biotechnol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Yang, F.; Massey, I.Y.; Guo, J.; Yang, S.; Pu, Y.; Zeng, W.; Tan, H. Microcystin-LR degradation utilizing a novel effective indigenous bacterial community YFMCD1 from Lake Taihu. J. Toxicol. Environ. Health A 2018, 81, 184–193. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, Y.; Sun, R.; Wei, H.; Li, Y.; Yin, L.; Pu, Y. Biodegradation of microcystin-LR and-RR by a novel microcystin-degrading bacterium isolated from Lake Taihu. Biodegradation 2014, 25, 447–457. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Rao, P.V.L. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Massey, I.Y.; Zhang, X.; Yang, F. Importance of bacterial biodegradation and detoxification processes of microcystins for environmental health. J. Toxicol. Environ. Health Part B 2018, 21, 357–369. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Zhang, X.; Xie, P. A review of reproductive toxicity of microcystins. J. Hazard. Mater. 2016, 301, 381–399. [Google Scholar] [CrossRef]
- Massey, I.Y.; Wu, P.; Wei, J.; Luo, J.; Ding, P.; Wei, H.; Yang, F. A Mini-Review on Detection Methods of Microcystins. Toxins 2020, 12, 641. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, D.J.; Kauffman, K.W.; Hall, R.A.; Huang, X.; Chu, F.S. Assessing Potential Health Risks from Microcystin Toxins in Blue-Green Algae Dietary Supplements. Environ. Health Perspect 2000, 108, 435–439. [Google Scholar] [CrossRef]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Xie, P. The dose makes the poison. Sci. Total Environ. 2018, 621, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X.; et al. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res. 2020, 174, 115638. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Yang, F. A Mini Review on Microcystins and Bacterial Degradation. Toxins 2020, 12. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Yang, F.; Wen, C.; Zheng, S.; Yang, S.; Chen, J.; Feng, X. Involvement of MAPK/ERK1/2 pathway in microcystin-induced microfilament reorganization in HL7702 hepatocytes. J. Toxicol. Environ. Health Part A 2018, 81, 1135–1141. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.A.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the Three Gorges Reservoir Region, China. Environ. Health Perspect 2011, 119, 1483–1488. [Google Scholar] [CrossRef]
- Zheng, C.; Zeng, H.; Lin, H.; Wang, J.; Feng, X.; Qiu, Z.; Chen, J.; Luo, J.; Luo, Y.; Huang, Y.; et al. Serum microcystin levels positively linked with risk of hepatocellular carcinoma: A case-control study in southwest China. Hepatology 2017, 66, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Huang, F.; Massey, I.Y.; Wen, C.; Zheng, S.; Xu, S.; Yang, F. Effects of Microcystin-LR on the Microstructure and Inflammation-Related Factors of Jejunum in Mice. Toxins 2019, 11, 482. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Huang, H.; Yang, L.; Zhang, X.; Zhang, S.; Liu, H.; Wang, Y.; Yuan, L.; Cheng, X.; Zhuang, D.; et al. Gastrointestinal toxicity induced by microcystins. World J. Clin. Cases 2018, 6, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, L.; Zhou, W.; Zhao, Q.; Wang, Y. Chronic exposure to microcystin-LR affected mitochondrial DNA maintenance and caused pathological changes of lung tissue in mice. Environ. Pollut. 2016, 210, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.M.; Cagido, V.R.; Ferraro, R.B.; Meyer-Fernandes, J.R.; Rocco, P.R.M.; Zin, W.A.; Azevedo, S.M.F.O. Effects of microcystin-LR on mouse lungs. Toxicon 2007, 50, 330–338. [Google Scholar] [CrossRef]
- Cao, L.; Massey, I.Y.; Feng, H.; Yang, F. A Review of Cardiovascular Toxicity of Microcystins. Toxins 2019, 11, 507. [Google Scholar] [CrossRef]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Fan, H.; Xie, P.; He, J. A review of neurotoxicity of microcystins. Environ. Sci. Pollut. Res. 2016, 23, 7211–7219. [Google Scholar] [CrossRef]
- Xiong, Q.; Xie, P.; Li, H.; Hao, L.; Li, G.; Qiu, T.; Liu, Y. Acute effects of microcystins exposure on the transcription of antioxidant enzyme genes in three organs (liver, kidney, and testis) of male Wistar rats. J. Biochem. Mol. Toxicol. 2010, 24, 361–367. [Google Scholar] [CrossRef]
- Yi, X.; Xu, S.; Huang, F.; Wen, C.; Zheng, S.; Feng, H.; Guo, J.; Chen, J.; Feng, X.; Yang, F. Effects of Chronic Exposure to Microcystin-LR on Kidney in Mice. Int. J. Environ. Res. Public Health 2019, 16, 5030. [Google Scholar] [CrossRef]
- Lin, H.; Liu, W.; Zeng, H.; Pu, C.; Zhang, R.; Qiu, Z.; Chen, J.; Wang, L.; Tan, Y.; Zheng, C.; et al. Determination of Environmental Exposure to Microcystin and Aflatoxin as a Risk for Renal Function Based on 5493 Rural People in Southwest China. Environ. Sci. Technol. 2016, 50, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Xie, P.; Li, L.; Guo, L.; Zhang, D.; Zhou, Q. Nephrotoxic effects from chronic toxic cyanobacterial blooms in fishes with different trophic levels in a large Chinese lake. Environ. Toxicol. Pharmacol. 2012, 33, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Sitprija, V.; Sitprija, S. Marine toxins and nephrotoxicity: Mechanism of injury. Toxicon 2019, 161, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, G.; Wu, Q.; Liu, C.; Shen, J.; Yan, W. Microcystin-LR exposure induced nephrotoxicity by triggering apoptosis in female zebrafish. Chemosphere 2019, 214, 598–605. [Google Scholar] [CrossRef]
- La-Salete, R.; Oliveira, M.M.; Palmeira, C.A.; Almeida, J.; Peixoto, F.P. Mitochondria a key role in microcystin-LR kidney intoxication. J. Appl. Toxicol. 2008, 28, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Xu, L.; Zhang, X.; Wang, Y.; Meng, X.; Miao, A.; Yang, L. Endoplasmic reticulum stress in murine liver and kidney exposed to microcystin-LR. Toxicon 2010, 56, 1334–1341. [Google Scholar] [CrossRef]
- Li, X. Effects of Chronic Exposure to Microcystin-LR on Somatic Mitochondrial DNA in Mice. Ph.D. Thesis, Nanjing University, Nanjing, China, 2015. [Google Scholar]
- Li, H.; Xie, P.; Li, G.; Hao, L.; Xiong, Q. In vivo study on the effects of microcystin extracts on the expression profiles of proto-oncogenes (c-fos, c-jun and c-myc) in liver, kidney and testis of male Wistar rats injected i.v. with toxins. Toxicon 2009, 53, 169–175. [Google Scholar] [CrossRef]
- Hao, L.; Xie, P.; Li, H.; Li, G.; Xiong, Q.; Wang, Q.; Qiu, T.; Liu, Y. Transcriptional alteration of cytoskeletal genes induced by microcystins in three organs of rats. Toxicon 2010, 55, 1378–1386. [Google Scholar] [CrossRef]
- Dias, E.; Matos, P.; Pereira, P.; Batoréu, M.C.C.; Silva, M.J.; Jordan, P. Microcystin-LR activates the ERK1/2 kinases and stimulates the proliferation of the monkey kidney-derived cell line Vero-E6. Toxicol. In Vitro 2010, 24, 1689–1695. [Google Scholar] [CrossRef]
- Fan, H.; Cai, Y.; Xie, P.; Xiao, W.; Chen, J.; Ji, W.; Zhao, S. Microcystin-LR stabilizes c-myc protein by inhibiting protein phosphatase 2A in HEK293 cells. Toxicology 2014, 319, 69–74. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, P.; Chen, J.; Liang, G. Distribution of microcystins in various organs (heart, liver, intestine, gonad, brain, kidney and lung) of Wistar rat via intravenous injection. Toxicon 2008, 52, 721–727. [Google Scholar] [CrossRef]
- Lei, H.; Xie, P.; Chen, J.; Liang, G.; Dai, M.; Zhang, X. Distribution of toxins in various tissues of crucian carp intraperitoneally injected with hepatotoxic microcystins. Environ. Toxicol. Chem. 2008, 27, 1167–1174. [Google Scholar] [CrossRef]
- Lei, H.; Xie, P.; Chen, J.; Liang, G.; Yu, T.; Jiang, Y. Tissue Distribution and Depuration of the Extracted Hepatotoxic Cyanotoxin Microcystins in Crucian Carp (Carassius carassius) Intraperitoneally Injected at a Sublethal Dose. Sci. World J. 2008, 8, 713–719. [Google Scholar] [CrossRef]
- Moutou, K.A.; Tsikogias, S.; Papadimitriou, T.; Kagalou, I. Oxidative stress in Cyprinus carpio to analyze microcystin impact in eutrophic shallow lakes: A preliminary study. J. Environ. Monit. 2012, 14, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Sedan, D.; Giannuzzi, L.; Rosso, L.; Marra, C.A.; Andrinolo, D. Biomarkers of prolonged exposure to microcystin-LR in mice. Toxicon 2013, 68, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Mitsoura, A.; Kagalou, I.; Papaioannou, N.; Berillis, P.; Mente, E.; Papadimitriou, T. The presence of microcystins in fish Cyprinus carpio tissues: A histopathological study. Int. Aquat. Res. 2013, 5, 1–16. [Google Scholar] [CrossRef]
- Bi, X.; Dai, W.; Wang, X.; Dong, S.; Zhang, S.; Zhang, D.; Wu, M. Microcystins distribution, bioaccumulation, and Microcystis genotype succession in a fish culture pond. Sci. Total Environ. 2019, 688, 380–388. [Google Scholar] [CrossRef]
- Jia, J.; Luo, W.; Lu, Y.; Giesy, J.P. Bioaccumulation of microcystins (MCs) in four fish species from Lake Taihu, China: Assessment of risks to humans. Sci. Total Environ. 2014, 487, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Asthana, R.K. Assessment of Microcystin Concentration in Carp and Catfish: A Case Study from Lakshmikund Pond, Varanasi, India. Bull. Environ. Contam. Toxicol. 2014, 92, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Zhang, J.; Luo, Y. Microcystin accumulation in bighead carp (Aristichthys nobilis) during a Microcystis-dominated bloom and risk assessment of the dietary intake in a fish pond in China. Environ. Sci. Pollut. Res. Int. 2015, 24, 8894–8902. [Google Scholar] [CrossRef]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Choudhuri, S.; Ogura, K.; Csanaky, I.L.; Lei, X.; Cheng, X.; Song, P.; Klaassen, C.D. Characterization of Organic Anion Transporting Polypeptide 1b2-null Mice: Essential Role in Hepatic Uptake/Toxicity of Phalloidin and Microcystin-LR. Toxicol. Sci. 2008, 103, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharm. 2010, 245, 9–20. [Google Scholar] [CrossRef]
- Feurstein, D.; Holst, K.; Fischer, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharm. 2009, 234, 247–255. [Google Scholar] [CrossRef]
- Zhang, Z.; Yu, S.; Wei, G.; Chen, W. Study on the whole and cells level distribution of Microcystin LR in mice. J. Toxicol. 2002, 16, 5–8. [Google Scholar]
- Li, L.; Xie, P.; Lei, H.; Zhang, X. Renal accumulation and effects of intraperitoneal injection of extracted microcystins in omnivorous crucian carp (Carassius auratus). Toxicon 2013, 70, 62–69. [Google Scholar] [CrossRef]
- Fischer, W.J.; Dietrich, D.R. Pathological and Biochemical Characterization of Microcystin-Induced Hepatopancreas and Kidney Damage in Carp (Cyprinus carpio). Toxicol. Appl. Pharm. 2000, 164, 73–81. [Google Scholar] [CrossRef]
- Greer, B.; Meneely, J.P.; Elliott, C.T. Uptake and accumulation of Microcystin-LR based on exposure through drinking water: An animal model assessing the human health risk. Sci. Rep. UK 2018, 8, 4913. [Google Scholar] [CrossRef]
- Falconer, I.R.; Burch, M.D.; Steffensen, D.A.; Choice, M.; Coverdale, O.R. Toxicity of the blue-green alga (cyanobacterium) Microcystis aeruginosa in drinking water to growing pigs, as an animal model for human injury and risk assessment. Environ. Toxicol. Water Qual. 1994, 9, 131–139. [Google Scholar] [CrossRef]
- Foss, A.J.; Aubel, M.T.; Gallagher, B.; Mettee, N.; Miller, A.; Fogelson, S.B. Diagnosing Microcystin Intoxication of Canines: Clinicopathological Indications, Pathological Characteristics, and Analytical Detection in Postmortem and Antemortem Samples. Toxins 2019, 11, 456. [Google Scholar] [CrossRef]
- Bautista, A.C.; Moore, C.E.; Lin, Y.; Cline, M.G.; Benitah, N.; Puschner, B. Hepatopathy following consumption of a commercially available blue-green algae dietary supplement in a dog. BMC Vet. Res. 2015, 11. [Google Scholar] [CrossRef]
- Rankin, K.; Alroy, K.; Kudela, R.; Oates, S.; Murray, M.; Miller, M. Treatment of Cyanobacterial (Microcystin) Toxicosis Using Oral Cholestyramine: Case Report of a Dog from Montana. Toxins 2013, 5, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Hermansky, S.J.; Stohs, S.J.; Eldeen, Z.M.; Roche, V.F.; Mereish, K.A. Evaluation of potential chemoprotectants against microcystin-LR hepatotoxicity in mice. J. Appl. Toxicol. 1991, 11, 65–73. [Google Scholar] [CrossRef]
- Kondo, F.; Ikai, Y.; Oka, H.; Okumura, M.; Ishikawa, N.; Harada, K.; Matsuura, K.; Murata, H.; Suzuki, M. Formation, characterization, and toxicity of the glutathione and cysteine conjugates of toxic heptapeptide microcystins. Chem. Res. Toxicol. 1992, 5, 591–596. [Google Scholar] [CrossRef]
- Wu, L.; Wang, Q.; Tao, M.; Chen, J.; Ma, Z.; Xie, P. Preliminary Study of the Distribution and Accumulation of GSH/Cys Metabolites of Hepatotoxic Microcystins-RR in Common Carp from a Lake with Protracted Cyanobacterial Bloom (Lake Taihu, China). Bull. Environ. Contam. Toxicol. 2013, 90, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, S.; Tao, M.; Xie, P.; Chen, J. Quantitative analysis of glutathione and cysteine S-conjugates of microcystin-LR in the liver, kidney and muscle of common carp (Cyprinus carpio) in Lake Taihu. J. Water Health 2017, 15, 300–307. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Chen, J.; Xie, P.; Zhang, D.; Li, G.; Wu, L.; Zhang, W.; Guo, X.; Li, S. Quantitatively evaluating detoxification of the hepatotoxic microcystins through the glutathione and cysteine pathway in the cyanobacteria-eating bighead carp. Aquat. Toxicol. 2012, 116–117, 61–68. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Xie, P.; Li, W.; Yu, Y.; Zhang, X.; Han, X.; Yu, X. Quantitative Study in the Process of Glutathione Detoxification of Microcystin. Asian J. Ecotoxicol. 2018, 13, 99–105. [Google Scholar]
- Li, W.; Chen, J.; Xie, P.; He, J.; Guo, X.; Tuo, X.; Zhang, W.; Wu, L. Rapid conversion and reversible conjugation of glutathione detoxification of microcystins in bighead carp (Aristichthys nobilis). Aquat. Toxicol. 2014, 147, 18–25. [Google Scholar] [CrossRef]
- Li, W.; He, J.; Chen, J.; Xie, P. Excretion pattern and dynamics of glutathione detoxification of microcystins in Sprague Dawley rat. Chemosphere 2018, 191, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.M.; Yuan, M.; Servaites, J.C.; Delgado, A.; Magalhães, V.F.; Hilborn, E.D.; Carmichael, W.W.; Azevedo, S.M.F.O. Sublethal exposure from microcystins to renal insufficiency patients in Rio de Janeiro, Brazil. Environ. Toxicol. 2006, 21, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Hilborn, E.D.; Carmichael, W.W.; Soares, R.M.; Yuan, M.; Servaites, J.C.; Barton, H.A.; Azevedo, S.M.F.O. Serologic evaluation of human microcystin exposure. Environ. Toxicol. 2007, 22, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Madhushankha, L.; Dhammika, M.A.; Charitha, P.; Tilak, A.; Lishantha, G. Cyanobacteria and cyanotoxins in well waters of the Girandurukotte, CKDu endemic area in Sri Lanka; do they drink safe water? J. Ecotechnol. Res. 2016, 1, 17–21. [Google Scholar]
- Chen, J.; Xie, P.; Li, L.; Xu, J. First Identification of the Hepatotoxic Microcystins in the Serum of a Chronically Exposed Human Population Together with Indication of Hepatocellular Damage. Toxicol. Sci. 2009, 108, 81–89. [Google Scholar] [CrossRef] [PubMed]
- R Bhattacharya, K.S.R.S. Toxicity Evaluation of Freshwater Cyanobacterium Microcystis Aeruginosa PCC 7806: II. Nephrotoxicity in Rats. Biomed. Environ. Sci. 1997, 10, 93–101. [Google Scholar]
- Pan, X.; Chang, F.; Liu, Y.; Li, D.; Xu, A.; Shen, Y.; Huang, Z. Mouse toxicity ofAnabaena flos-aquae from Lake Dianchi, China. Environ. Toxicol. 2009, 24, 10–18. [Google Scholar] [CrossRef]
- Humpage, A.; Falconer, I.; Bernard, C.; Froscio, S.; Fabbro, L. Toxicity of the cyanobacterium Limnothrix AC0243 to male Balb/c mice. Water Res. 2012, 46, 1576–1583. [Google Scholar] [CrossRef]
- Li, G.; Xie, P.; Li, H.; Hao, L.; Xiong, Q.; Qiu, T.; Liu, Y. Acute effects of microcystins on the transcription of 14 glutathione S-transferase isoforms in Wistar rat. Environ. Toxicol. 2011, 26, 187–194. [Google Scholar] [CrossRef]
- Yang, L.; Shen, F.; Lu, B.; Tong, X.; Qin, J.; Fu, Z. Study the Toxicity of Microcystin in Water of Dianchi Lake on Mice Kidney. J. Kunming Univ. 2012, 34, 52–54. [Google Scholar]
- Adamovsky, O.; Palikova, M.; Ondrackova, P.; Zikova, A.; Kopp, R.; Mares, J.; Pikula, J.; Paskerova, H.; Kohoutek, J.; Blaha, L. Biochemical and histopathological responses of Wistar rats to oral intake of microcystins and cyanobacterial biomass. Neuro Endocrinol. Lett. 2013, 34 (Suppl. 2), 11–20. [Google Scholar]
- Hooser, S.B.; Beasley, V.R.; Lovell, R.A.; Carmichael, W.W.; Haschek, W.M. Toxicity of microcystin LR, a cyclic heptapeptide hepatotoxin from Microcystis aeruginosa, to rats and mice. Vet. Pathol. 1989, 26, 246–252. [Google Scholar] [CrossRef]
- Sun, F. Comparison of the Effect of Microcytin-RR on Acute Kidney, Liver Injury and Their Possible Mechanisms in Rats and Mice. Master’s Thesis, Huazhong University of Science and Technology, Wuhan, China, 2017. [Google Scholar]
- Lowe, J.; Souza-Menezes, J.; Freire, D.S.; Mattos, L.J.; Castiglione, R.C.; Barbosa, C.M.L.; Santiago, L.; Ferrão, F.M.; Cardoso, L.H.D.; Da Silva, R.T.; et al. Single sublethal dose of microcystin-LR is responsible for different alterations in biochemical, histological and physiological renal parameters. Toxicon 2012, 59, 601–609. [Google Scholar] [CrossRef]
- Lamei, L.; Lirong, S. Acute toxicity of microcystin-LR in BALB/c mice. J. First Med. Coll. PLA 2005, 25, 565–566. [Google Scholar]
- Xu, C. Microcystin Contamination in Aquatic Environment of the Three Gorges Reservior and Antagonism Study on MC-LR Induced Toxicity. Master’s Thesis, Third Military Medical University, Chongqing, China, 2005. [Google Scholar]
- Zhong, Q. Effect of Microcystin-RR on Water Metabolism and Neurobehavior in Mice and Related Mechanism Exporation. Ph.D. Thesis, Huazhong University of Science and Technology, Wuhan, China, 2017. [Google Scholar]
- Dong, L. Study on Toxicity of Microcystin-LR on Liver, Kidney and Testicle Cells in Male Mice. Master’s Thesis, Zhengzhou University, Zhengzhou, China, 2008. [Google Scholar]
- Gaudin, J.; Huet, S.; Jarry, G.; Fessard, V. In vivo DNA damage induced by the cyanotoxin microcystin-LR: Comparison of intra-peritoneal and oral administrations by use of the comet assay. Mutat. Res. Genet. Toxicol. Environ. Mutagenes. 2008, 652, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Pichardo, S.; Jos, A.; Gómez-Amores, L.; Mate, A.; Vazquez, C.M.; Cameán, A.M. Antioxidant enzyme activity and lipid peroxidation in liver and kidney of rats exposed to microcystin-LR administered intraperitoneally. Toxicon 2005, 45, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; He, M.; Cui, D.; Xiong, W.; Zhang, Q. The protective effects of glutathione on renal oxidative damage induced by microcystin-LR on mice. Chongqing Med. 2016, 45, 320–322. [Google Scholar]
- Li, Z. Study on Mechanisms of Microcystin-LR Toxicity Based on Metabonomics. Master’s Thesis, Jiangnan University, Wuxi, China, 2009. [Google Scholar]
- Sedan, D.; Andrinolo, D.; Telese, L.; Giannuzzi, L.; de Alaniz, M.J.T.; Marra, C.A. Alteration and recovery of the antioxidant system induced by sub-chronic exposure to microcystin-LR in mice: Its relation to liver lipid composition. Toxicon 2010, 55, 333–342. [Google Scholar] [CrossRef]
- Milutinović, A.; Sedmak, B.; Horvat-Znidarsic, I.; Suput, D. Renal injuries induced by chronic intoxication with microcystins. Cell. Mol. Biol. Lett. 2002, 7, 139. [Google Scholar]
- Milutinović, A.; Živin, M.; Zorc-Pleskovič, R.; Sedmak, B.; Šuput, D. Nephrotoxic effects of chronic administration of microcystins -LR and -YR. Toxicon 2003, 42, 281–288. [Google Scholar] [CrossRef]
- Khan, S.A.; Ghosh, S.; Wickstrom, M.; Miller, L.A.; Hess, R.; Haschek, W.M.; Beasley, V.R. Comparative pathology of microcystin-lr in cultured hepatocytes, fibroblasts, and renal epithelial cells. Nat. Toxins 1995, 3, 119–128. [Google Scholar] [CrossRef]
- Chen, J.; Xu, L.; Fu, W.; Xing, M. Bax expression of rat nnephrocyte NRK induced by microcystin-LR. Acta Hydrobiol. Sin. 2004, 28, 450–451. [Google Scholar]
- Nobre, A.C.; Jorge, M.C.; Menezes, D.B.; Fonteles, M.C.; Monteiro, H.S. Effects of microcystin-LR in isolated perfused rat kidney. Braz. J. Med. Biol. Res. 1999, 32, 985–988. [Google Scholar] [CrossRef]
- Dias, E.; Andrade, M.; Alverca, E.; Pereira, P.; Batoréu, M.C.C.; Jordan, P.; Silva, M.J. Comparative study of the cytotoxic effect of microcistin-LR and purified extracts from Microcystis aeruginosa on a kidney cell line. Toxicon 2009, 53, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Alverca, E.; Andrade, M.; Dias, E.; Sam Bento, F.; Batoréu, M.C.C.; Jordan, P.; Silva, M.J.; Pereira, P. Morphological and ultrastructural effects of microcystin-LR from Microcystis aeruginosa extract on a kidney cell line. Toxicon 2009, 54, 283–294. [Google Scholar] [CrossRef]
- Menezes, C.; Alverca, E.; Dias, E.; Sam-Bento, F.; Pereira, P. Involvement of endoplasmic reticulum and autophagy in microcystin-LR toxicity in Vero-E6 and HepG2 cell lines. Toxicol. In Vitro 2013, 27, 138–148. [Google Scholar] [CrossRef]
- Li, T.; Ying, L.; Wang, H.; Li, N.; Fu, W.; Guo, Z.; Xu, L. Microcystin-LR induces ceramide to regulate PP2A and destabilize cytoskeleton in HEK293 cells. Toxicol. Sci. 2012, 128, 147–157. [Google Scholar] [CrossRef]
- Molina, R.; Moreno, I.; Pichardo, S.; Jos, A.; Moyano, R.; Monterde, J.G.; Cameán, A. Acid and alkaline phosphatase activities and pathological changes induced in Tilapia fish (Oreochromis sp.) exposed subchronically to microcystins from toxic cyanobacterial blooms under laboratory conditions. Toxicon 2005, 46, 725–735. [Google Scholar] [CrossRef]
- Li, L.; Xie, P.; Chen, J. Biochemical and ultrastructural changes of the liver and kidney of the phytoplanktivorous silver carp feeding naturally on toxic Microcystis blooms in Taihu Lake, China. Toxicon 2007, 49, 1042–1053. [Google Scholar] [CrossRef]
- Li, L.; Xie, P.; Guo, L.; Ke, Z.; Zhou, Q.; Liu, Y.; Qiu, T. Field and Laboratory Studies on Pathological and Biochemical Characterization of Microcystin-Induced Liver and Kidney Damage in the Phytoplanktivorous Bighead Carp. Sci. World J. 2008, 8, 121–137. [Google Scholar] [CrossRef]
- Atencio, L.; Moreno, I.; Jos, A.; Pichardo, S.; Moyano, R.; Blanco, A.; Cameán, A.M. Dose-dependent antioxidant responses and pathological changes in tenca (Tinca tinca) after acute oral exposure to Microcystis under laboratory conditions. Toxicon 2008, 52, 1–12. [Google Scholar] [CrossRef]
- Kotak, B.G.; Semalulu, S.; Fritz, D.L.; Prepas, E.E.; Hrudey, S.E.; Coppock, R.W. Hepatic and renal pathology of intraperitoneally administered microcystin-LR in rainbow trout (Oncorhynchus mykiss). Toxicon 1996, 34, 517–525. [Google Scholar] [CrossRef]
- Ma, J.; Feng, Y.; Xie, W.; Li, X. PP2A (PR65) in Silver Carp: cDNA Cloning and Expression Analysis. J. Biochem. Mol. Toxicol. 2015, 29, 399–409. [Google Scholar] [CrossRef]
- Prieto, A.I.; Jos, A.; Pichardo, S.; Moreno, I.; Camean, A.M. Protective role of vitamin E on the microcystin-induced oxidative stress in tilapia fish (Oreochromis niloticus). Environ. Toxicol. Chem. 2008, 27, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Puerto, M.; Gutiérrez-Praena, D.; Prieto, A.I.; Pichardo, S.; Jos, A.; Miguel-Carrasco, J.L.; Vazquez, C.M.; Cameán, A.M. Subchronic effects of cyanobacterial cells on the transcription of antioxidant enzyme genes in tilapia (Oreochromis niloticus). Ecotoxicology 2011, 20, 479–490. [Google Scholar] [CrossRef]
- Feng, Y.; Ma, J.; Xiang, R.; Li, X. Alterations in microRNA expression in the tissues of silver carp (Hypophthalmichthys molitrix) following microcystin-LR exposure. Toxicon 2017, 128, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, L.; Liu, W.; Qiao, Q.; Wu, K.; Wen, J.; Huang, C.; Tang, R.; Zhang, X. Involvement of oxidative stress and cytoskeletal disruption in microcystin-induced apoptosis in CIK cells. Aquat. Toxicol. 2015, 165, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharm. 2005, 203, 257–263. [Google Scholar] [CrossRef]
- Xia, H.; Song, T.; Wang, L.; Jiang, L.; Zhou, Q.; Wang, W.; Liu, L.; Yang, P.; Zhang, X. Effects of dietary toxic cyanobacteria and ammonia exposure on immune function of blunt snout bream (Megalabrama amblycephala). Fish Shellfish Immun. 2018, 78, 383–391. [Google Scholar] [CrossRef]
- Jayaraj, R.; Lakshmana Rao, P.V. Protein phosphorylation profile and adduct formation in liver and kidney of microcystin-LR-treated mice. Toxicon. 2006, 48, 272–277. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Lin, S.; Wang, B.; Xing, M.; Guo, Z.; Xu, L. MCLR-induced PP2A inhibition and subsequent Rac1 inactivation and hyperphosphorylation of cytoskeleton-associated proteins are involved in cytoskeleton rearrangement in SMMC-7721 human liver cancer cell line. Chemosphere 2014, 112, 141–153. [Google Scholar] [CrossRef]
- He, Y.; Huang, X.; Ding, X. Advances in Research on Toxicity and Mechanism of Action of Microcystins. Food Sci. 2020, 41, 290–298. [Google Scholar]
- Tavares, D.; Paulino, M.G.; Terezan, A.P.; Batista Fernandes, J.; Giani, A.; Fernandes, M.N. Osmoregulatory disturbance in Neotropical fish exposed to the crude extracts of the cyanobacterium, Radiocystis fernandoi. Aquat. Toxicol. 2019, 216, 105315. [Google Scholar] [CrossRef]
- Chen, L.; Xie, P. Mechanisms of Microcystin-induced Cytotoxicity and Apoptosis. Mini Rev. Med. Chem. 2016, 16, 1018–1031. [Google Scholar] [CrossRef]
- Jayaraj, R.; Anand, T.; Rao, P.V.L. Activity and gene expression profile of certain antioxidant enzymes to microcystin-LR induced oxidative stress in mice. Toxicology 2006, 220, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Mao, G. The Acute Toxic Effects of Oral Aministration of Microcystin LR in SD Rats. Food Res. Dev. 2013, 13, 13–16. [Google Scholar]
- Kim, Y.D.; Kim, W.J.; Shin, Y.K.; Lee, D.; Kim, Y.; Kim, J.K.; Rhee, J. Microcystin-LR bioconcentration induces antioxidant responses in the digestive gland of two marine bivalves Crassostrea gigas and Mytilus edulis. Aquat. Toxicol. 2017, 188, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, X.; Zhou, W.; Qiao, Q.; Liang, H.; Li, G.; Wang, J.; Cai, F. The interactive effects of cytoskeleton disruption and mitochondria dysfunction lead to reproductive toxicity induced by microcystin-LR. PLoS ONE. 2013, 8, e53949. [Google Scholar] [CrossRef]
- Zhou, M.; Tu, W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef]
- Ding, W.; Shen, H.; Ong, C. Microcystic Cyanobacteria Extract Induces Cytoskeletal Disruption and Intracellular Glutathione Alteration in Hepatocytes. Environ. Health Perspect 2000, 108, 605–609. [Google Scholar] [CrossRef]
- Dias, E.; Louro, H.; Pinto, M.; Santos, T.; Antunes, S.; Pereira, P.; Silva, M.J. Genotoxicity of Microcystin-LR in In Vitro and In Vivo Experimental Models. Biomed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Pathmalal, M.M.; Tennekoon, K.H.; Piyathilaka, M.A.P.C.; De Silva, B.G.D.N.; Samarakoon, S.R.; Chanthirika, S. Microcystin-LR-induced cytotoxicity and apoptosis in human embryonic kidney and human kidney adenocarcinoma cell lines. Microbiology 2015, 161, 819–828. [Google Scholar] [CrossRef]



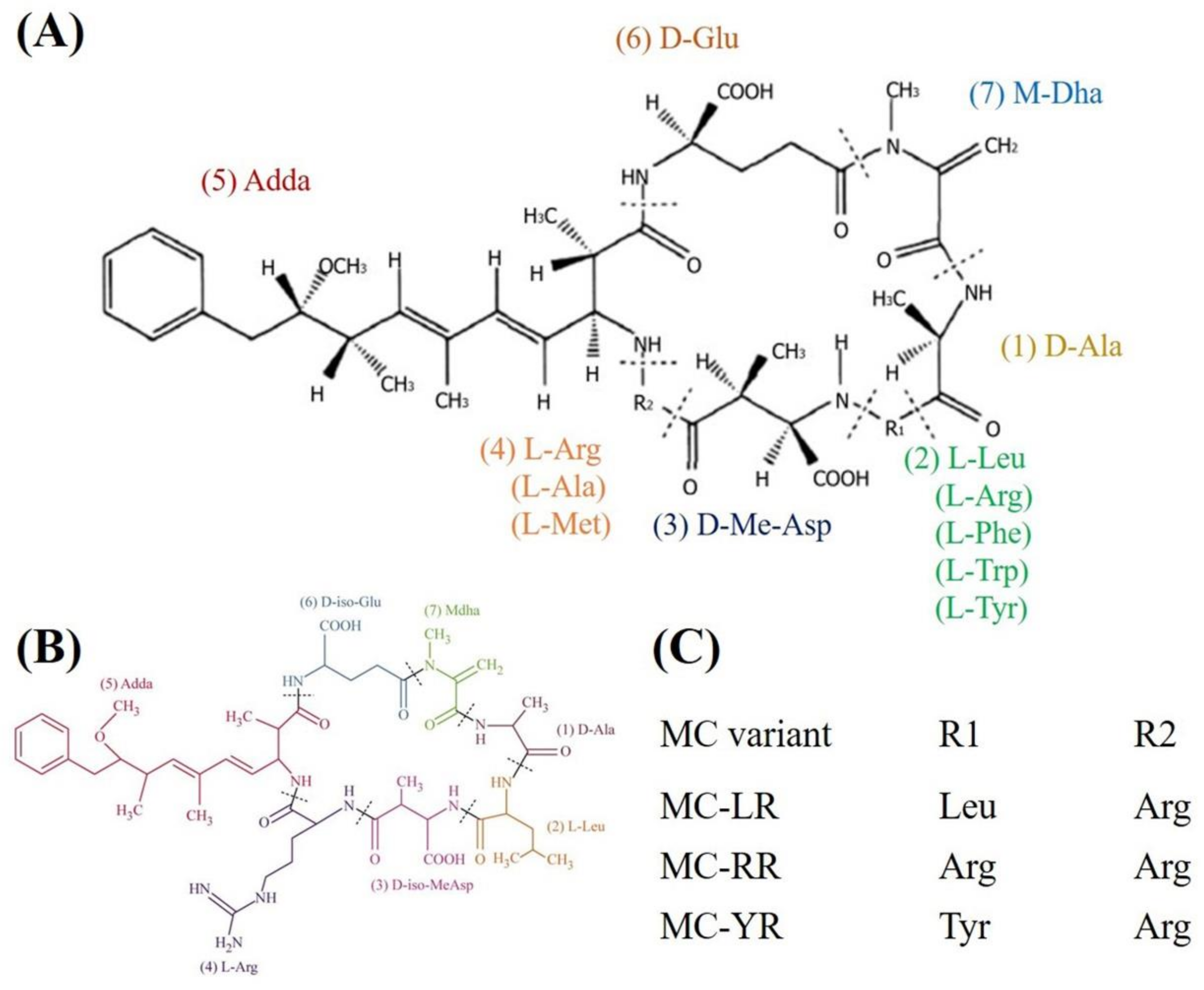{kind=link}
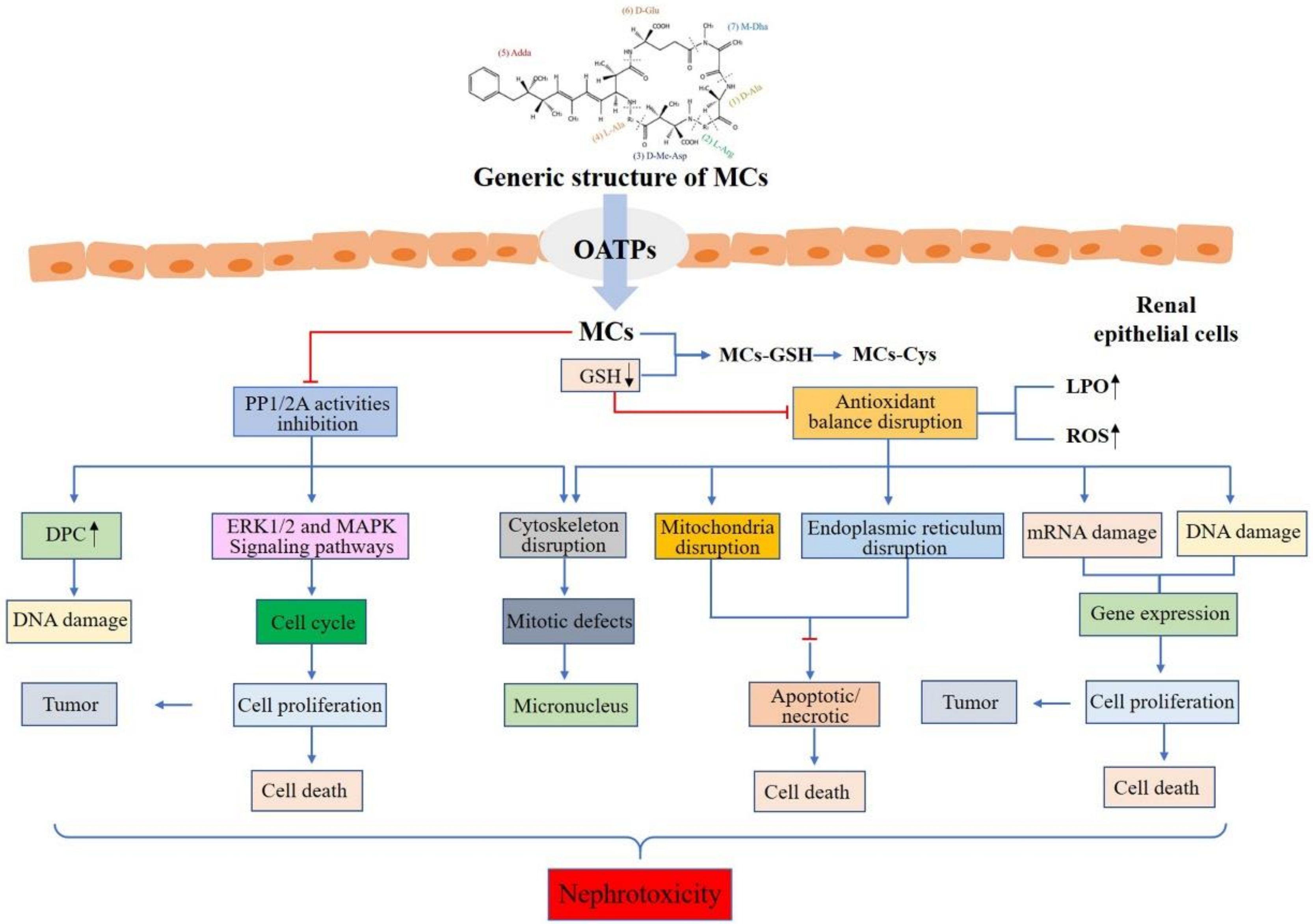{kind=link}
| Country/Year | Sample Size | Investigated Effects | Cyanotoxins(Detection Method) | Conclusion | References |
|---|---|---|---|---|---|
| Brazil/1996 | 95 | Blood biochemical indicators | MCs (ELISA, LC-MS) | Incident— Cyanotoxin poisoning | Hilborn et al. [73] |
| Brazil/2001 | 44 | Blood biochemical indicators | MCs (ELISA, LC-MS) | Incident— Cyanotoxin poisoning | Soares et al. [72] |
| China/2005 | 76 | Renal function indicators | MCs (LC-MS) | Epidemiological study— positive association | Chen et al. [75] |
| China/2013 | 5493 | Renal function indicators | MC-LR (ELISA) | Epidemiological study— positive association | Lin et al. [31] |
| Sri Lanka/2016 | 330 | Chronic kidney disease of unknown etiology (CKDu) | MCs (LC-MS) | Epidemiological study— inconclusive | Liyanage et al. [74] |
| Test Organism/System | Exposure | Toxicant | Concentration/Dose | Time Point | Toxic Effects | References |
|---|---|---|---|---|---|---|
| Male rats | I.P. | Microcystis cell extracts | 0.5, 1.0, 2.0 LD50 | - | BUN↑, SCr↑, LDH↓, GPT↓, followed with hematuria, albuminuria and bilirubinuria | Bhattacharya et al. [76] |
| Male KM mice | I.P. | Cyanobacterial crude extracts | 0.25, 0.5 and 1.0 g/kg | 10, 24 h | BUN↑, SCr↑, T-AOC↓, microstructural damage | Pan et al. [77] |
| Balb/c mice | I.P. | Cyanobacterial crude extracts | 180 and 195 mg/mL | 2, 4, 24 h | Microstructural damage | Humpage et al. [78] |
| Male Wistar rats | I.V. | Cyanobacterial crude extracts | 86.7 μg MC-LR eq/kg | 1, 2, 4, 6, 12, 24 h | mRNA of c-fos ↑, c-jun↑ and c-mys↑; protein of c-fos↑, c-jun↑ | Li et al. [38] |
| Wistar rats | I.V. | Cyanobacterial crude extracts | 80 μg MC-LR eq/kg | 1, 2, 4, 6, 12, 24 h | Modulation of CAT, Mn-SOD, Cu, Zn-SOD,GR, GPX, γ-GCS transcription | Xiong et al. [29] |
| Wistar rats | I.V. | Cyanobacterial crude extracts | 87 μg MC-LR eq/kg | 1, 2, 4, 6, 12, 24 h | Modulation of 14 GSTs transcription | Li et al. [79] |
| KM mice | I.G. | Cyanobacterial crude extracts | 0.1 mL | 1 month | Microstructural damage | Yang et al. [80] |
| Wistar rats | P.O. | Cyanobacterial crude extracts | 136 and 928 μg MC-LR eq/kg | 28 days | Ultrastructural damage, MDA↑, GR↑ and LPO↑ | Adamovsky et al. [81] |
| Test Organism/System | Exposure | Concentration/Dose | Time Point | Toxic Effects | References |
|---|---|---|---|---|---|
| SD rats | I.P. | 20, 40, 80, 120, 160, 180, 200, or 400 μg/kg | 1, 12, 24 h | BUN↑, SCr↑, ALT↑, AST↑, TBIL↑, followed with ultrastructural damage | Hooser et al. [82] |
| Wistar rats | I.P. | 55 μg/kg | 24 h | Glomerular filtration rate↑, albuminuria↑, ROS↑ and Na+ reabsorption↓, microstructural and ultrastructural damage | Lowe et al. [84] |
| Balb/c mice | I.P. | 22, 43 μg/kg | 0.5, 4 h | Kidney relative weight↑, UA↑ | Lei et al. [85] |
| KM mice | I.P. | 10 μg/kg | 13 days | ALT↑, AST↑, ALP↑,BUN↑, SCr↑, MDA↑, LPO↑ and microstructural damage | Xu et al. [86] |
| KM mice | I.P. | 3, 6, 12 μg/kg | 7 days | Kidney absolute weight↓, Kidney relative weight↑, DNA-protein crosslinking↑ and protein carbonyl↑ | Dong et al. [88] |
| ICR mice | I.P. | 20 μg/kg | 21 days | Apoptosis, CHOP↓, caspase-12↓, Bcl-2↑ | Qin et al. [36] |
| Wistar rats | I.P. | 100, 150 μg/kg | 8 h | GSH-Px↓, GR↓, SOD↓, CAT↓, LPO↑ | Moreno et al. [90] |
| KM mice | I.P. | 5 μg/kg | 15 days | MDA↑, GSH↓, SOD↓, CAT↓ | Han et al. [91] |
| KM mice | I.P. | 30 μg/kg | 1, 4, 8 h | ALT↑, SOD↑, CAT↑, BUN first↓ then↑, and all the biochemical indicators are reversible | Li et al. [92] |
| SD rats | I.P. | 30 μg/kg | 1, 3, 7, 12 h | Microstructural damage | Li et al. [92] |
| Male mice | I.P. | 25μg/kg | 1, 2 months | Kidney relative weight↑, GSH↓, GPH-Px↓, SOD↓, CAT↓, NOx↑ and partial of indicators are reversible after one month’s cleaning period | Sedan et al. [93] |
| C57BL/6 mice | P.O. | 1, 30, 60, 90,120 μg/L | 3, 6 months | BUN↓, microstructural and ultrastructural damage | Yi et al. [30] |
| Wistar rats | I.P. | 10 μg/L | 8 months | Microstructural damage | Milutinović et al. [94] |
| Wistar rats | I.P. | 10 μg/L | 8 months | Microstructural damage, apoptosis↑, cytoskeleton disruption↑ | Milutinović et al. [95] |
| Test Organism/System | Toxicant | Concentration/Dose | Time Point | Toxic Effects | References |
|---|---|---|---|---|---|
| Rat kidney epithelial cells (ATCC 1571) | MC-LR | 13.3 μM | 24 h | Apoptosis↑, ultrastructural damage | Khan et al. [96] |
| Rat NRK cell | MC-LR | 10, 100, 1000 nM | 4, 6, 7, 8 h | Bax↑ | Chen et al. [97] |
| Isolated rat kidney | MC-LR | 1 μg/L | 2 h | Urine flow↑, perfusion pressure↑, glomerular filtration rate↑, sodium tubule transport fraction↓, microstructural damage | Nobre et al. [98] |
| Vero-E6 cell | MC-LR, cyanobacterial crude extracts | 1.4–175 nM | 24, 48, 72 h | Cell viability↓ | Dias et al. [99] |
| Vero-E6 cell | MC-LR, cyanobacterial crude extracts | 5, 50, 500, 5000 nM | 24 h | Cell proliferation↑, P38↑, JNK↑, ERK1/2 activity↑ | Dias et al. [40] |
| Vero-E6 cell | MC-LR | 1.3, 2.5, 5, 10, 20, 30, 40, 50, 75, 100, 150 μM | 24, 48, 72 h | Cell viability↓, autophagy, apoptosis, necrosis, ultrastructural damage | Alverca et al. [100] |
| Vero-E6 cell | MC-LR | 6, 12, 25, 50 μM | 24 h | Cell viability↓, autophagy, cytoskeleton disruption↑ | Menezes et al. [101] |
| HEK293 | MC-LR, MC-RR, MC-LW, MC- LF | 0.01, 0.1, 1, 10, 100, 1000 nM | 4 h | Cell viability↓, phosphatase activity↓, modulation of Oatps expression | Fischer et al. [54] |
| HEK293 | MC-LR | 2, 10 μM | 24 h | Ceramide↑, PP2A activity↑, cytoskeleton disruption↑ | Li et al. [102] |
| HEK293 | MC-LR | 10 μM | 24 h | PP2A activity↓, alteration of c-myc expression | Fan et al. [41] |
| Test Organism/System | Exposure | Toxicant | Concentration/Dose | Time Point | Toxic Effects | References |
|---|---|---|---|---|---|---|
| Microcystins crude extracts | ||||||
| Carp | I.G. | cyanobacterial crude extracts | 400 μg/kg MC-LR | 3 days | Microstructural damage, apoptosis, necrosis, cell shedding, proteinaceous casts↑ at the cortico-medullary junction | Fischer et al. [58] |
| Tilapia | Immersion | cyanobacterial crude extracts | 60.0 μg MC-LR/fish | 14, 21 days | Microstructural damage, ALP↑, ACP↑ | Molina et al. [103] |
| Silver carp | Immersion | cyanobacterial crude extracts | - | - | CAT↑, GST↑, GSH↑, GPX↑, SOD↑, ultrastructural changes | Li et al. [104] |
| Bighead Carp | I.P. | cyanobacterial crude extracts | 200, 400 μg/kg MC-LR | 24 h | CAT↑, microstructural damage | Li et al. [105] |
| Tenca | Immersion | cyanobacterial crude extracts | 5, 11, 25, 55 μg | CAT↓, SOD↓, ultrastructural damage | Atencio et al. [106] | |
| Silver carp, Bighead Carp, Carassius auratus, Culter ilishaeformis | Immersion | cyanobacterial crude extracts | - | - | Microstructural damage, alterations of antioxidant enzymes | Qiu et al. [32] |
| Pure microcystins | ||||||
| Oncorhynchus mykiss | I.P. | MC-LR | 400, 1000 μg/kg | 16 h | Microstructural damage | Kotak et al. [107] |
| Silver carp | I.V. | MC-LR | 50, 200 μg/kg | 1, 3, 8, 12 h | Downregulated PP2A-A transcription | Ma et al. [108] |
| Tilapia | Immersion | MC-LR | 120 μg/kg | 7 days | Dysfunction in redox dynamic balance, CAT↓, SOD↓, GSH-Px↓, GR↓ | Prieto et al. [109] |
| Tilapia | Immersion | MC-LR | 60 μg MC-LR | 21 days | LPO↑, alterations of antioxidant enzymes, modulation of GPx and GST genes transcription | Puerto et al. [110] |
| Silver carp | I.V. | MC-LR | 50 and 200 μg/kg | 8, 24, 48 h | Modulation of 7 miRNAs transcription | Feng et al. [111] |
| Zebrafish | Immersion | MC-LR | 1, 5, 25 μg/L | 60 days | Microstructural damage, modulation of genes transcription, apoptosis, ROS↑ | Wang et al. [34] |
| CIK cell | - | MC-LR | 1, 10, 100 μg/L | 24, 48 h | Cell viability↓, G2/M phase arrest, ROS↑, MDA↑, modulation of antioxidant enzymes including CAT and SOD, modulation of cytoskeletal genes (β-actin, lc3a, and keratin) transcription | Huang et al. [112] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Yi, X.; Liu, W.; Zhang, C.; Massey, I.Y.; Yang, F.; Tian, L. A Review of Nephrotoxicity of Microcystins. Toxins 2020, 12, 693. https://doi.org/10.3390/toxins12110693
Xu S, Yi X, Liu W, Zhang C, Massey IY, Yang F, Tian L. A Review of Nephrotoxicity of Microcystins. Toxins. 2020; 12(11):693. https://doi.org/10.3390/toxins12110693
Chicago/Turabian StyleXu, Shuaishuai, Xiping Yi, Wenya Liu, Chengcheng Zhang, Isaac Yaw Massey, Fei Yang, and Li Tian. 2020. "A Review of Nephrotoxicity of Microcystins" Toxins 12, no. 11: 693. https://doi.org/10.3390/toxins12110693
APA StyleXu, S., Yi, X., Liu, W., Zhang, C., Massey, I. Y., Yang, F., & Tian, L. (2020). A Review of Nephrotoxicity of Microcystins. Toxins, 12(11), 693. https://doi.org/10.3390/toxins12110693