Genomics and Proteomics Analyses Revealed Novel Candidate Pesticidal Proteins in a Lepidopteran-Toxic Bacillus thuringiensis Strain

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
2.1. Crystal Morphology, Protein Profile, and Insecticidal Activity of KhF Strain
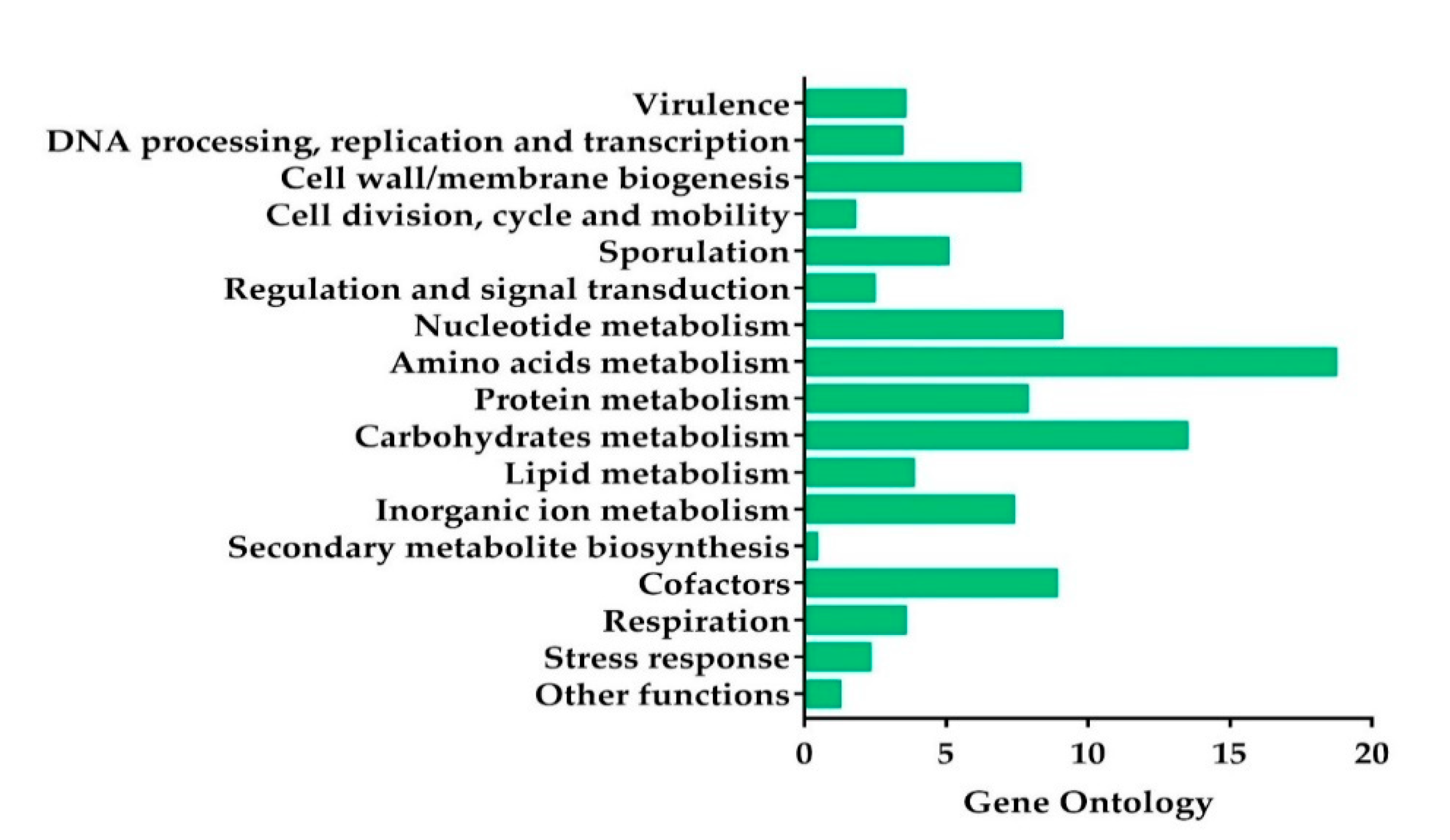
2.2. Genome Sequencing, Gene Annotation, and Phylogenetic Analysis
2.3. Search for Virulence Factors and Bacterial Toxins in the Genome
2.4. Proteomics of Spores and Crystals
2.5. Cluster Tree Analysis of the KhFA and KhFB Toxins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Microscopic Observation, Protein Profiling, and Spore/Crystal Mixture Preparation
5.2. Insect Colonies
5.3. Toxicity Assays
5.4. DNA Isolation, Genome Assembly, and Annotation Analysis
5.5. Protein Identification by LC-MS/MS Analysis
5.6. Phylogenetic Analyses
5.7. Availability of Data and Material
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lacey, L.; Grzywacz, D.; Shapiro-Ilan, D.; Frutos, R.; Brownbridge, M.; Goettel, M. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis Toxins: An Overview of Their Biocidal Activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Almanza, G.; Esparza-Ibarra, E.L.; Ayala-Luján, J.L.; Mercado-Reyes, M.; Godina-González, S.; Hernández-Barrales, M.; Olmos, J. The Cytocidal Spectrum of Bacillus thuringiensis Toxins: From Insects to Human Cancer Cells. Toxins 2020, 12, 301. [Google Scholar] [CrossRef] [PubMed]
- Crickmore, N. Beyond the spore—Past and future developments of Bacillus thuringiensis as a biopesticide. J. Appl. Microbiol. 2006, 101, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Meissle, M.; Bigler, F. Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat. Biotechnol. 2006, 24, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fu, Z.; Ding, X.; Xia, L. Evaluating the Insecticidal Genes and Their Expressed Products in Bacillus thuringiensis Strains by Combining PCR with Mass Spectrometry. Appl. Environ. Microbiol. 2008, 74, 6811–6813. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ding, X.; Sun, Y.; Yang, Q.; Xiao, X.; Cao, Z.; Xia, L. Proteomic Analysis of Bacillus thuringiensis at Different Growth Phases by Using an Automated Online Two-Dimensional Liquid Chromatography-Tandem Mass Spectrometry Strategy. Appl. Environ. Microbiol. 2012, 78, 5270–5279. [Google Scholar] [CrossRef]
- Gomis-Cebolla, J.; Ricietto, A.P.S.; Ferré, J. A Genomic and Proteomic Approach to Identify and Quantify the Expressed Bacillus thuringiensis Proteins in the Supernatant and Parasporal Crystal. Toxins 2018, 10, 193. [Google Scholar] [CrossRef]
- Murawska, E.; Fiedoruk, K.; Bideshi, D.K.; Swiecicka, I. Complete Genome Sequence of Bacillus thuringiensis subsp. thuringiensis Strain IS5056, an Isolate Highly Toxic to Trichoplusia ni. Genome Announc. 2013, 1. [Google Scholar] [CrossRef]
- Fayad, N.; Patiño-Navarrete, R.; Kambris, Z.; Antoun, M.; Osta, M.; Chopineau, J.; Mahillon, J.; El Chamy, L.; Sanchis, V.; Awad, M.K. Characterization and Whole Genome Sequencing of AR23, a Highly Toxic Bacillus thuringiensis Strain Isolated from Lebanese Soil. Curr. Microbiol. 2019, 76, 1503–1511. [Google Scholar] [CrossRef]
- Alves, G.B.; Melo, F.L.; Oliveira, E.E.; Haddi, K.; Costa, L.T.M.; Dias, M.L.; Campos, F.; Pereira, E.J.G.; Corrêa, R.F.T.; Ascêncio, S.D.; et al. Comparative genomic analysis and mosquito larvicidal activity of four Bacillus thuringiensis serovar israelensis strains. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Chen, H.; Jiang, X.; Wang, J.; Gelbič, I.; Guan, X.; Zhang, L. Whole genome sequence analysis of the mosquitocidal Bacillus thuringiensis LLP29. Arch. Microbiol. 2020, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Zhu, L.; Liu, Y.; Crickmore, N.; Peng, D.; Ruan, L.; Sun, M. Mining New Crystal Protein Genes from Bacillus thuringiensis on the Basis of Mixed Plasmid-Enriched Genome Sequencing and a Computational Pipeline. Appl. Environ. Microbiol. 2012, 78, 4795–4801. [Google Scholar] [CrossRef]
- Shikov, A.E.; Malovichko, Y.V.; Skitchenko, R.K.; Nizhnikov, A.A.; Antonets, K. No More Tears: Mining Sequencing Data for Novel Bt Cry Toxins with CryProcessor. Toxins 2020, 12, 204. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Schwartz, J.M.; Finson, N.; Johnson, M.W. Inheritance of Resistance to Bacillus thuringiensis in Diamondback Moth (Lepidoptera: Plutellidae). J. Econ. Èntomol. 1992, 85, 1046–1055. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]
- He, J.; Wang, J.; Yin, W.; Shao, X.; Zheng, H.; Li, M.; Zhao, Y.; Sun, M.; Wang, S.; Yu, Z. Complete Genome Sequence of Bacillus thuringiensis subsp. chinensis Strain CT-43. J. Bacteriol. 2011, 193, 3407–3408. [Google Scholar] [CrossRef]
- Rang, J.; He, H.; Wang, T.; Ding, X.; Zuo, M.; Quan, M.; Sun, Y.; Yu, Z.; Hu, S.; Xia, L. Comparative Analysis of Genomics and Proteomics in Bacillus thuringiensis 4.0718. PLoS ONE 2015, 10, e0119065. [Google Scholar] [CrossRef]
- Zhu, L.; Peng, D.; Wang, Y.; Ye, W.; Zheng, J.; Zhao, C.; Han, D.; Geng, C.; Ruan, L.; He, J.; et al. Genomic and transcriptomic insights into the efficient entomopathogenicity of Bacillus thuringiensis. Sci. Rep. 2015, 5, 14129. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and Its Pesticidal Crystal Proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef]
- Khorramnejad, A.; Talaei-Hassanloui, R.; Hosseininaveh, V.; Bel, Y.; Escriche, B. Characterization of new Bacillus thuringiensis strains from Iran, based on cytocidal and insecticidal activity, proteomic analysis and gene content. BioControl 2018, 63, 807–818. [Google Scholar] [CrossRef]
- Dworzanski, J.P.; Dickinson, D.N.; Deshpande, S.V.; Snyder, A.P.; Eckenrode, B.A. Discrimination and Phylogenomic Classification of Bacillus anthracis-cereus-thuringiensis Strains Based on LC-MS/MS Analysis of Whole Cell Protein Digests. Anal. Chem. 2010, 82, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.-C.; Shen, L.-F.; Tsai, M.-C.; Chak, K.-F. The IspA protease’s involvement in the regulation of the sporulation process of Bacillus thuringiensis is revealed by proteomic analysis. Biochem. Biophys. Res. Commun. 2003, 312, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.J.; Di Falco, M.; Souleimanov, A.; Smith, D.L. Proteomic analysis of the bacteriocin thuricin 17 produced by Bacillus thuringiensis NEB17. FEMS Microbiol. Lett. 2006, 255, 27–32. [Google Scholar] [CrossRef]
- DelVecchio, V.G.; Connolly, J.P.; Alefantis, T.G.; Walz, A.; Quan, M.A.; Patra, G.; Ashton, J.M.; Whittington, J.T.; Chafin, R.D.; Liang, X.; et al. Proteomic Profiling and Identification of Immunodominant Spore Antigens of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Appl. Environ. Microbiol. 2006, 72, 6355–6363. [Google Scholar] [CrossRef]
- Fu, Z.; Sun, Y.; Xia, L.; Ding, X.; Mo, X.; Li, X.; Huang, K.; Zhang, Y. Assessment of protoxin composition of Bacillus thuringiensis strains by use of polyacrylamide gel block and mass spectrometry. Appl. Microbiol. Biotechnol. 2008, 79, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Caballero, J.; Jiménez-Moreno, N.; Orera, I.; Williams, T.; Fernández, A.B.; Villanueva, M.; Ferré, J.; Caballero, P.; Ancín-Azpilicueta, C. Unraveling the Composition of Insecticidal Crystal Proteins in Bacillus thuringiensis: A Proteomics Approach. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Chen, D.; Xu, D.; Li, M.; He, J.; Gong, Y.; Wu, D.; Sun, M.; Yu, Z. Proteomic analysis of Bacillus thuringiensis ΔphaC mutant BMB171/PHB-1 reveals that the PHB synthetic pathway warrants normal carbon metabolism. Proteomics 2012, 75, 5176–5188. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mei, H.; Zheng, C.; Qian, H.; Cui, C.; Fu, Y.; Su, J.; Liu, Z.; Yu, Z.; He, J. The Metabolic Regulation of Sporulation and Parasporal Crystal Formation in Bacillus thuringiensis Revealed by Transcriptomics and Proteomics. Mol. Cell. Proteom. 2013, 12, 1363–1376. [Google Scholar] [CrossRef]
- Wang, L.; Mei, H.; Qian, H.; Tang, Q.; Liu, X.; Yu, Z.; He, J. Expression profile and regulation of spore and parasporal crystal formation-associated genes in Bacillus thuringiensis. J. Proteome Res. 2013, 12, 5487–5501. [Google Scholar] [CrossRef]
- Li, X.; Ding, X.; Xia, L.; Sun, Y.; Yuan, C.; Yin, J. Proteomic Analysis of Bacillus thuringiensis Strain 4.0718 at Different Growth Phases. Sci. World J. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Daas, M.S.; Rosana, A.R.; Acedo, J.Z.; Douzane, M.; Nateche, F.; Kebbouche-Gana, S.; Vederas, J.C. Insights into the draft genome sequence of bioactives-producing Bacillus thuringiensis DNG9 isolated from Algerian soil-oil slough. Stand. Genom. Sci. 2018, 13, 25. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Bacillus thuringiensis Toxin Nomenclature. Available online: http://www.lifesci.sussex.ac.uk/home/NeilCrickmore/Bt/ (accessed on 22 June 2020).
- Silva, J.C.; Gorenstein, M.V.; Li, G.Z.; Vissers, J.P.; Geromanos, S.J. Absolute quantification of proteins by LCMSE: A virtue of parallel MS acquisition. Mol. Cell Proteom. 2006, 5, 144–156. [Google Scholar] [CrossRef]
- Smith, D.G.; Gingras, G.; Aubin, Y.; Cyr, T.D. Design and expression of a QconCAT protein to validate Hi3 protein quantification of influenza vaccine antigens. J. Proteom. 2016, 146, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 107438. [Google Scholar] [CrossRef]
- Raddadi, N.; Belaouis, A.; Tamagnini, I.; Hansen, B.M.; Hendriksen, N.B.; Boudabous, A.; Cherif, A.; Daffonchio, D. Characterization of polyvalent and safe Bacillus thuringiensis strains with potential use for biocontrol. J. Basic Microbiol. 2008, 49, 293–303. [Google Scholar] [CrossRef]
- Guan, P.; Ai, P.; Dai, X.; Zhang, J.; Xu, L.; Zhu, J.; Li, Q.; Deng, Q.; Li, S.; Wang, S.; et al. Complete Genome Sequence of Bacillus thuringiensis Serovar Sichuansis Strain MC28. J. Bacteriol. 2012, 194, 6975. [Google Scholar] [CrossRef]
- Kantas, D.; Papatsiros, V.; Tassis, P.; Giavasis, I.; Bouki, P.; Tzika, E. A feed additive containing Bacillus toyonensis (Toyocerin®) protects against enteric pathogens in post weaning piglets. J. Appl. Microbiol. 2015, 118, 727–738. [Google Scholar] [CrossRef]
- Chen, J.; Hu, S.; Ji, D.; Gao, Z.; Wang, H.; Yang, Y.; Chen, Y.; Gu, J. Hemolysin BL from novel Bacillus toyonensis BV-17 induces antitumor activity both in vitro and in vivo. Gut Microbes 2020, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Berry, C. The bacterium, Lysinibacillus sphaericus, as an insect pathogen. J. Invertebr. Pathol. 2012, 109, 1–10. [Google Scholar] [CrossRef]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Diversity of Bacillus thuringiensis Crystal Toxins and Mechanism of Action. In Advances in Insect Physiology; Elsevier BV: Amsterdam, The Netherlands, 2014; Volume 47, pp. 39–87. [Google Scholar]
- Peng, D.; Chai, L.; Wang, F.; Zhang, F.; Ruan, L.; Sun, M. Synergistic activity between Bacillus thuringiensis Cry6Aa and Cry55Aa toxins against Meloidogyne incognita. Microb. Biotechnol. 2011, 4, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Crickmore, N.; Peng, D.; Sun, M. Are nematodes a missing link in the confounded ecology of the entomopathogen Bacillus thuringiensis? Trends Microbiol. 2015, 23, 341–346. [Google Scholar] [CrossRef]
- Nielsen-Leroux, C.; Gaudriault, S.; RamaRao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef]
- Dalhammar, G.; Steiner, H. Characterization of inhibitor A, a protease from Bacillus thuringiensis which degrades attacins and cecropins, two classes of antibacterial proteins in insects. J. Biol. Inorg. Chem. 1984, 139, 247–252. [Google Scholar] [CrossRef]
- Miyoshi, S.-I.; Shinoda, S. Microbial metalloproteases and pathogenesis. Microbes Infect. 2000, 2, 91–98. [Google Scholar] [CrossRef]
- Altincicek, B.; Linder, M.; Linder, D.; Preissner, K.T.; Vilcinskas, A. Microbial Metalloproteinases Mediate Sensing of Invading Pathogens and Activate Innate Immune Responses in the Lepidopteran Model Host Galleria mellonella. Infect. Immun. 2006, 75, 175–183. [Google Scholar] [CrossRef]
- Lovgren, A.; Zhang, M.; Engström, A.; Dalhammar, G.; Landen, R. Molecular characterization of immune inhibitor A, a secreted virulence protease from Bacillus thuringiensis. Mol. Microbiol. 1990, 4, 2137–2146. [Google Scholar] [CrossRef] [PubMed]
- Fedhila, S.; Nel, P.; Lereclus, D. The InhA2 Metalloprotease of Bacillus thuringiensis Strain 407 Is Required for Pathogenicity in Insects Infected via the Oral Route. J. Bacteriol. 2002, 184, 3296–3304. [Google Scholar] [CrossRef] [PubMed]
- Malovichko, Y.V.; Nizhnikov, A.A.; Antonets, K.S. Repertoire of the Bacillus thuringiensis Virulence Factors Unrelated to Major Classes of Protein Toxins and Its Role in Specificity of Host-Pathogen Interactions. Toxins 2019, 11, 347. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, K.; Lewis, R.J. Molecular insights into the initiation of sporulation in Gram-positive bacteria: New technologies for an old phenomenon. FEMS Microbiol. Rev. 2005, 29, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Lyu, Y.; Feng, E.; Zhu, L.; Pan, C.; Wang, D.; Liu, X.; Zhu, L. SpoVG Is Necessary for Sporulation in Bacillus anthracis. Microorganisms 2020, 8, 548. [Google Scholar] [CrossRef] [PubMed]
- Phadtare, S. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 2004, 6, 125–136. [Google Scholar] [PubMed]
- Gryspeirt, A.; Grégoire, J.-C. Effects of Two Varieties of Bacillus thuringiensis Maize on the Biology of Plodia interpunctella. Toxins 2012, 4, 373–389. [Google Scholar] [CrossRef]
- Shojaaddini, M.; López, M.J.; Moharramipour, S.; Khodabandeh, M.; Talebi, A.A.; Vilanova, C.; Latorre, A.; Porcar, M. A Bacillus thuringiensis strain producing epizootics on Plodia interpunctella: A case study. J. Stored Prod. Res. 2012, 48, 52–60. [Google Scholar] [CrossRef]
- Ricietto, A.P.S.; Gomis-Cebolla, J.; Vilas-Bôas, G.T.; Ferré, J. Susceptibility of Grapholita molesta (Busck, 1916) to formulations of Bacillus thuringiensis, individual toxins and their mixtures. J. Invertebr. Pathol. 2016, 141, 1–5. [Google Scholar] [CrossRef]
- Travers, R.S.; Martin, P.A.W.; Reichelderfer, C.F. Selective Process for Efficient Isolation of Soil Bacillus spp. Appl. Environ. Microbiol. 1987, 53, 1263–1266. [Google Scholar] [CrossRef]
- Stewart, G.S.; Johnstone, K.; Hagelberg, E.; Ellar, D.J. Commitment of bacterial spores to germinate A measure of the trigger reaction. Biochem. J. 1981, 198, 101–106. [Google Scholar] [CrossRef]
- Estela, A.; Escriche, B.; Ferrã, J. Interaction of Bacillus thuringiensis Toxins with Larval Midgut Binding Sites of Helicoverpa armigera (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 2004, 70, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nat. Cell Biol. 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Lozzia, G.C.; Manachini, B. Susceptibility of Ostrinia nubilalis Hübner (Lepidoptera: Crambidae) to Bacillus thuringiensis var. kurstaki. Bull Insectol. 2003, 56, 215–219. [Google Scholar]
- Moar, W.J.; Pusztai-Carey, M.; Van Faassen, H.; Bosch, D.; Frutos, R.; Rang, C.; Luo, K.; Adang, M.J. Development of Bacillus thuringiensis CryIC Resistance by Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 1995, 61, 2086–2092. [Google Scholar] [CrossRef]
- Guennelon, G.; Audemard, H.; Fremond, J.C.; El Idrissi Ammari, M.A. Progrès rélisés dans l’élevage permanent du Carpocapse (Laspeyresia pomonella L.) sur milieu artificiel. Agronomie 1981, 1, 59–64. [Google Scholar] [CrossRef]
- Liao, C.; Heckel, D.G.; Akhurst, R. Toxicity of Bacillus thuringiensis insecticidal proteins for Helicoverpa armigera and Helicoverpa punctigera (Lepidoptera: Noctuidae), major pests of cotton. J. Invertebr. Pathol. 2002, 80, 55–63. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2018, 47, D687–D692. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Şahin, B.; Gomis-Cebolla, J.; Güneş, H.; Ferré, J. Characterization of Bacillus thuringiensis isolates by their insecticidal activity and their production of Cry and Vip3 proteins. PLoS ONE 2018, 13, e0206813. [Google Scholar] [CrossRef] [PubMed]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a Next Generation Search Engine That Uses Sequence Temperature Values and Feature Probabilities to Identify Peptides from Tandem Mass Spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Soufiane, B.; Côté, J.-C. Discrimination among Bacillus thuringiensis H serotypes, serovars and strains based on 16S rRNA, gyrB and aroE gene sequence analyses. Antonie Leeuwenhoek 2008, 95, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Molecular Evolution, Phylogenetics and Epidemiology. FigTree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 8 September 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Species | Spores/mL | % Mortality 1 | % Functional Mortality 1 |
|---|---|---|---|
| G. molesta | 108 | 59 ± 13 | 87 ± 7 |
| 109 | 90 ± 14 | 95 ± 7 | |
| O. nubilalis | 108 | 3 ± 3 | 3 ± 3 |
| 109 | 3 ± 3 | 3 ± 3 | |
| H. armigera | 108 | 6 ± 6 | 6 ± 6 |
| 109 | 6 ± 6 | 6 ± 6 | |
| S. exigua | 108 | 0 ± 0 | 0 ± 0 |
| 109 | 7 ± 6 | 19 ± 9 | |
| P. interpunctella2 | 108 | 42 ± 2 | - |
| Annotation | Features | Gene Content | % Gene Content 1 | Length (Mb) | % Length 2 |
|---|---|---|---|---|---|
| Total | Genome content 3 | 6457 | - | 4.99 | (100) |
| tRNAs | 62 | - | 0.006 | 0.001 | |
| Repeat region | 123 | - | 0.035 | 0.007 | |
| Blast search | Total sequences 4 | 6272 | (100) | 4.95 | 99.19 |
| Annotated genes 5 | 4494 | 71.65 | 4.13 | 82.76 | |
| Hypothetical genes 6 | 1778 | 28.35 | 0.82 | 16.43 |
| Gene Identity | Length (aa) | Scaffold | Closest Homolog | Similarity (%) | Coverage (%) | E-Value | SignalIP Server |
|---|---|---|---|---|---|---|---|
| peg55201 | 867 | 73 | WP_098163495.1|hypothetical protein N | 99.54 | 100 | 0.0 | Yes |
| AEB52299.1|Vip4Aa1 C | 35.81 | 57.83 | 6.0−93 | ||||
| peg59361 | 1223 | 81 | WP_153593625.1|ETX/Mtx2 family from Bt N | 67.67 | 100 | 0.0 | No |
| AGC08395.1|Vip1Ad1C | 26.9 | 60.60 | 1.0−21 | ||||
| peg309 | 333 | 114 | WP_088083339| hypothetical protein N | 97 | 100 | 0.0 | Yes |
| ADK79125.1|Vip1Aa1 C | 27.41 | 59.26 | 4.0−12 | ||||
| peg2096 | 566 | 31 | WP_000734347.1|hypothetical protein N | 100 | 100 | 0.0 | Yes |
| AGC08395.1|Vip1Ad1 C | 25.62 | 58.68 | 5.0−11 | ||||
| peg3627 | 739 | 51 | WP_000811547| hypothetical protein N | 100 | 100 | 0.0 | Yes |
| AAR40886.1|Vip1Ba1 C | 27.97 | 58.05 | 2.0−12 | ||||
| peg4213 | 347 | 59 | WP_098475832| RICIN domain-containing protein N | 80.35 | 100 | 0.0 | Yes |
| CAH56541.1|Cry49Aa1 C | 23.98 | 67.54 | 7.0−25 | ||||
| peg5521 | 184 | 73 | PHE82359.1| hypothetical protein N | 97.81 | 100 | 5.0−124 | No |
| CAI40768.1|Vip2Ad1 C | 27.88 | 61.21 | 9.0−13 | ||||
| peg5522 | 502 | 73 | WP_098163497.1|hypothetical protein N | 99 | 100 | 0 | Yes |
| AAO86513.1|Vip2Ac1 C | 28.64 | 60.56 | 8.0−14 | ||||
| peg5937 | 216 | 81 | WP_153593624.1|hypothetical protein from Bt N | 89.39 | 92 | 1.0−127 | No |
| - 2 | - | - | - |
| Category | Protein Description | Accession Number | Matched Peptides 1 | Sequence Coverage 2 |
|---|---|---|---|---|
| Pathogenesis | Immune inhibitor A | P23382 | 156 | 50 |
| Bacillolysin | P05806 | 10 | 16 | |
| Sporulation proteins | Putative septation protein | A9VN61 | 14 | 57 |
| Stage 0 sporulation protein A | P52933 | 5 | 27 | |
| Protein biosynthesis | 60 kDa chaperonin | Q4MPR6 | 20 | 32 |
| 1-pyrroline-5-carboxylate dehydrogenase | A9VRG6 | 28 | 40 | |
| Homocysteine methyltransferase | B7IVP2 | 20 | 22 | |
| 10 kDa chaperonin | Q81VE2 | 22 | 48 | |
| RNA-binding protein | Q65JA8 | 7 | 21 | |
| Elongation factor | B7IT17 | 5 | 16 | |
| Hut operon positive regulatory protein | Q81Y44 | 4 | 38 | |
| Serine hydroxymethyltransferase | A9VSB4 | 6 | 15 | |
| Leucine dehydrogenase | P0A393 | 4 | 14 | |
| Phosphoribosyl-ATP pyrophosphatase | Q6HLE1 | 6 | 13 | |
| Glutamine synthetase | P19064 | 18 | 21 | |
| Purine biosynthesis protein | Q6HPA0 | 14 | 30 | |
| Translation initiation factor | Q81VQ7 | 5 | 76 | |
| Metabolism | Methylmalonate semialdehyde dehydrogenase | B7IW48 | 85 | 57 |
| Enolase | B7IP20 | 70 | 65 | |
| Glucose-6-phosphate isomerase | Q816G0 | 16 | 41 | |
| Other functions | DNA protection | Q8RPQ1 | 45 | 67 |
| Aminopeptidase | Q81XS5 | 37 | 37 | |
| Superoxide dismutase | Q81LW0 | 38 | 90 | |
| Pyridoxal 5′-phosphate synthase | A9VM99 | 31 | 55 | |
| Cytosol aminopeptidase | Q816E3 | 32 | 37 | |
| Purine nucleoside phosphorylase | Q81T09 | 8 | 21 | |
| Major cold shock protein | Q81TW8 | 2 | 22 |
| Gene Identity | Unused ProtScore | Sequence Coverage (%) | Matched Peptides | Function |
|---|---|---|---|---|
| peg5936 | 188.93 | 92.96 | 171 | WP_153593625.1|ETX/MTX2 family from Bt |
| peg5610 | 40.00 | 74.18 | 23 | MULTISPECIES: elongation factor Tu [Bacillaceae]|WP_001029617.1 |
| peg6159 | 40.09 | 35.65 | 22 | MULTISPECIES: PrkA family serine protein kinase [Bacillaceae]|WP_000353271.1 |
| peg3524 | 26.07 | 67.07 | 20 | MULTISPECIES: N-acetylmuramoyl-L-alanine amidase [Bacillaceae]|WP_000135303.1 |
| peg4292 | 33.24 | 40.36 | 18 | MULTISPECIES: succinate dehydrogenase flavoprotein subunit [Bacillaceae]|WP_000676745.1 |
| peg1615 | 30.53 | 33.68 | 18 | MULTISPECIES: peptide ABC transporter substrate-binding protein [Bacillaceae]|WP_000823458.1 |
| peg5937 | 28.39 | 98.60 | 20 | Hypothetical Protein|WP_087969954.1 |
| Gene Identity | Protein Molecular Mass (kDa) | Unused ProtScore | Sequence Coverage (%) | Matched Peptides | Hi3 (%) 2 | Shared Peptides 3 | Unique Peptides 4 |
|---|---|---|---|---|---|---|---|
| peg5936 | 134.32 1 | 188.93 | 92.96 | 171 | 79.15 | 23 | 23 |
| peg5937 | 24.23 1 | 28.39 | 98.60 | 20 | 4.05 | 6 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khorramnejad, A.; Gomis-Cebolla, J.; Talaei-Hassanlouei, R.; Bel, Y.; Escriche, B. Genomics and Proteomics Analyses Revealed Novel Candidate Pesticidal Proteins in a Lepidopteran-Toxic Bacillus thuringiensis Strain. Toxins 2020, 12, 673. https://doi.org/10.3390/toxins12110673
Khorramnejad A, Gomis-Cebolla J, Talaei-Hassanlouei R, Bel Y, Escriche B. Genomics and Proteomics Analyses Revealed Novel Candidate Pesticidal Proteins in a Lepidopteran-Toxic Bacillus thuringiensis Strain. Toxins. 2020; 12(11):673. https://doi.org/10.3390/toxins12110673
Chicago/Turabian StyleKhorramnejad, Ayda, Joaquín Gomis-Cebolla, Reza Talaei-Hassanlouei, Yolanda Bel, and Baltasar Escriche. 2020. "Genomics and Proteomics Analyses Revealed Novel Candidate Pesticidal Proteins in a Lepidopteran-Toxic Bacillus thuringiensis Strain" Toxins 12, no. 11: 673. https://doi.org/10.3390/toxins12110673
APA StyleKhorramnejad, A., Gomis-Cebolla, J., Talaei-Hassanlouei, R., Bel, Y., & Escriche, B. (2020). Genomics and Proteomics Analyses Revealed Novel Candidate Pesticidal Proteins in a Lepidopteran-Toxic Bacillus thuringiensis Strain. Toxins, 12(11), 673. https://doi.org/10.3390/toxins12110673