Efficacy of Bacillus subtilis ANSB060 Biodegradation Product for the Reduction of the Milk Aflatoxin M1 Content of Dairy Cows Exposed to Aflatoxin B1


 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Production Performances
2.2. Plasma Biochemical Indices
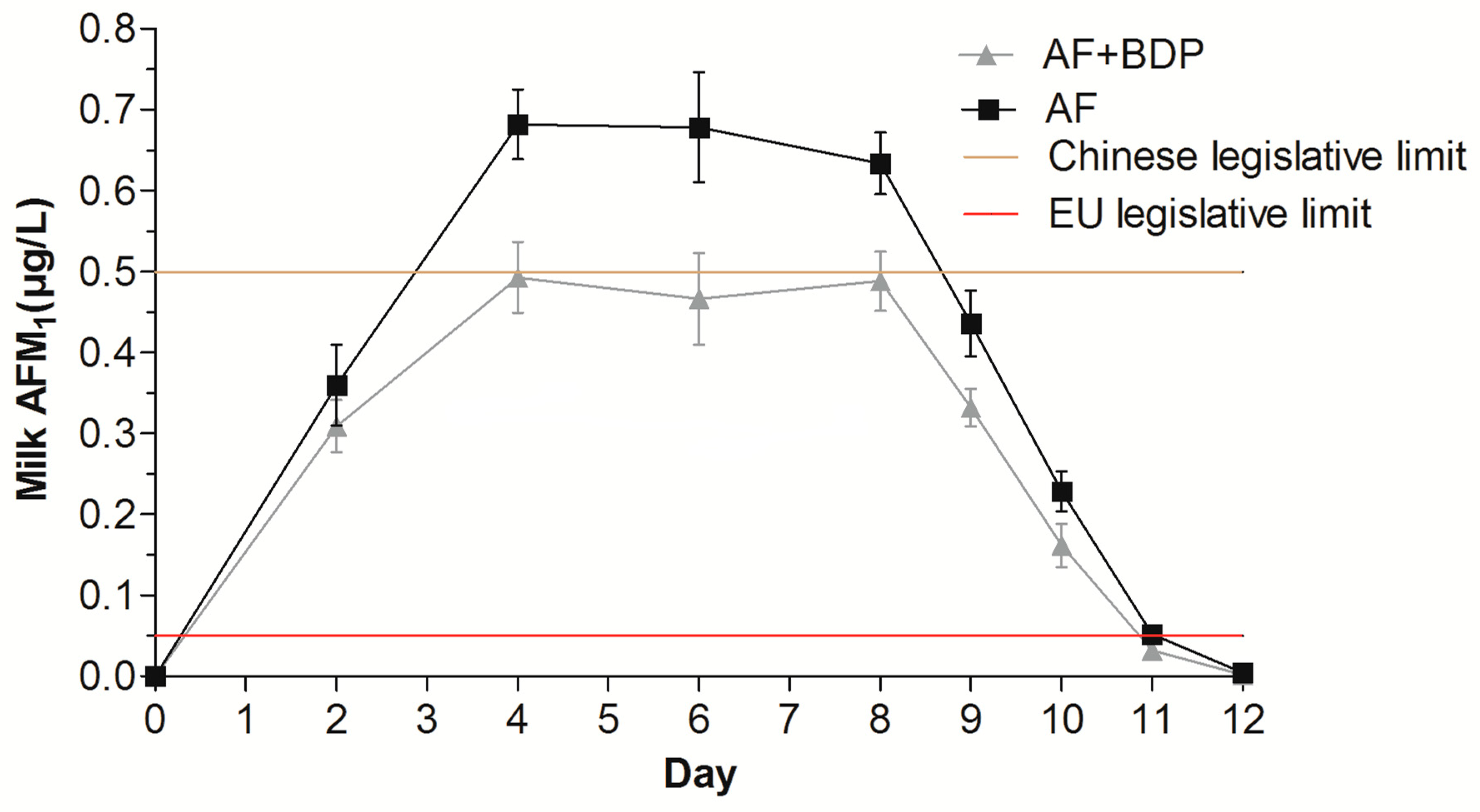
2.3. AFM1 Content in Milk
3. Conclusions
4. Materials and Methods
4.1. Animals, Experimental Design and Diets
4.2. Sample Collection and Analysis
4.3. Quantification of Aflatoxin M1 in Milk
4.4. Calculations
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Britzi, M.; Friedman, S.; Miron, J.; Solomon, R.; Cuneah, O.; Shimshoni, J.A.; Soback, S.; Ashkenazi, R.; Armer, S.; Shlosberg, A. Carry-over of aflatoxin B1 to aflatoxin M1 in high yielding Israeli cows in mid- and late-lactation. Toxins 2013, 5, 173–183. [Google Scholar] [CrossRef]
- Sadia, A.; Jabbar, M.A.; Deng, Y.; Hussain, E.A.; Riffat, S.; Naveed, S.; Arif, M. A survey of aflatoxin M1 in milk and sweets of Punjab, Pakistan. Food Control 2012, 26, 235–240. [Google Scholar] [CrossRef]
- Accinelli, C.; Mencarelli, M.; Saccà, M.L.; Vicari, A.; Abbas, H.K. Managing and monitoring of Aspergillus flavus in corn using bioplastic-based formulations. Crop Prot. 2012, 32, 30–35. [Google Scholar] [CrossRef]
- Verheecke, C.; Liboz, T.; Mathieu, F. Microbial degradation of aflatoxin B1: Current status and future advances. Int. J. Food Microbiol. 2016, 237, 1–9. [Google Scholar] [CrossRef]
- Maki, C.R.; Thomas, A.D.; Elmore, S.E.; Romoser, A.A.; Harvey, R.B.; Ramirez-Ramirez, H.A.; Phillips, T.D. Effects of calcium montmorillonite clay and aflatoxin exposure on dry matter intake, milk production, and milk composition. J. Dairy Sci. 2016, 99, 1039–1046. [Google Scholar] [CrossRef]
- Pate, R.T.; Paulus Compart, D.M.; Cardoso, F.C. Aluminosilicate clay improves production responses and reduces inflammation during an aflatoxin challenge in lactating Holstein cows. J. Dairy Sci. 2018, 101, 11421–11434. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.L.; Wang, Y.M.; Nennich, T.D.; Li, Y.; Liu, J.X. Transfer of dietary aflatoxin B1 to milk aflatoxin M1 and effect of inclusion of adsorbent in the diet of dairy cows. J. Dairy Sci. 2015, 98, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed Sci. Tech. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- De Mil, T.; Devreese, M.; Broekaert, N.; Fraeyman, S.; De Backer, P.; Croubels, S. In vitro adsorption and in vivo pharmacokinetic interaction between doxycycline and frequently used mycotoxin binders in broiler chickens. J. Agric. Food Chem. 2015, 63, 4370–4375. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mule, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef]
- Vanhoutte, I.; Audenaert, K.; De Gelder, L. Biodegradation of mycotoxins: Tales from known and unexplored worlds. Front. Microbiol. 2016, 7, 561. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Liu, D.; Mo, X.; Xie, C.; Yao, D. A fungal enzyme with the ability of aflatoxin B1 conversion: Purification and ESI-MS/MS identification. Microbiol. Res. 2011, 166, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E.; et al. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Eshelli, M.; Harvey, L.; Edrada-Ebel, R.; McNeil, B. Metabolomics of the bio-degradation process of aflatoxin B1 by actinomycetes at an initial pH of 6.0. Toxins 2015, 7, 439–456. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Q.; Zhao, L.; Lei, Y.; Shan, Y.; Ji, C. Isolation of Bacillus subtilis: Screening for aflatoxins B1, M1, and G1 detoxification. Eur. Food Res. Technol. 2011, 232, 957–962. [Google Scholar] [CrossRef]
- Rao, K.R.; Vipin, A.V.; Hariprasad, P.; Anu Appaiah, K.A.; Venkateswaran, G. Biological detoxification of aflatoxin B1 by Bacillus licheniformis CFR1. Food Control 2017, 71, 234–241. [Google Scholar]
- Ma, Q.G.; Gao, X.; Zhou, T.; Zhao, L.H.; Fan, Y.; Li, X.Y.; Lei, Y.P.; Ji, C.; Zhang, J.Y. Protective effect of Bacillus subtilis ANSB060 on egg quality, biochemical and histopathological changes in layers exposed to aflatoxin B1. Poult. Sci. 2012, 91, 2852–2857. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, L.; Ma, Q.; Li, X.; Shi, H.; Zhou, T.; Zhang, J.; Ji, C. Effects of Bacillus subtilis ANSB060 on growth performance, meat quality and aflatoxin residues in broilers fed moldy peanut meal naturally contaminated with aflatoxins. Food Chem. Toxicol. 2013, 59, 748–753. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, Q.; Ma, S.; Zhang, J.; Jia, R.; Ji, C.; Zhao, L. Ameliorating effects of Bacillus subtilis ANSB060 on growth performance, antioxidant functions, and aflatoxin residues in ducks fed diets contaminated with aflatoxins. Toxins 2016, 9, 1. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, L.; Zhao, L.; Wang, X.; Wang, D.; Huang, C.; Zhang, J.; Ji, C.; Ma, Q. Influence of Bacillus subtilis ANSB060 on growth, digestive enzyme and aflatoxin residue in Yellow River carp fed diets contaminated with aflatoxin B1. Food Chem. Toxicol. 2018, 113, 108–114. [Google Scholar] [CrossRef]
- Deng, K.D.; Xiao, Y.; Ma, T.; Tu, Y.; Diao, Q.Y.; Chen, Y.H.; Jiang, J.J. Ruminal fermentation, nutrient metabolism, and methane emissions of sheep in response to dietary supplementation with Bacillus licheniformis. Anim. Feed Sci. Tech. 2018, 241, 38–44. [Google Scholar] [CrossRef]
- Souza, V.L.; Lopes, N.M.; Zacaroni, O.F.; Silveira, V.A.; Pereira, R.A.N.; Freitas, J.A.; Almeida, R.; Salvati, G.G.S.; Pereira, M.N. Lactation performance and diet digestibility of dairy cows in response to the supplementation of Bacillus subtilis spores. Livest. Sci. 2017, 200, 35–39. [Google Scholar] [CrossRef]
- Kutz, R.E.; Sampson, J.D.; Pompeu, L.B.; Ledoux, D.R.; Spain, J.N.; Vázquez-Añón, M.; Rottinghaus, G.E. Efficacy of Solis, NovasilPlus, and MTB-100 to reduce aflatoxin M1 levels in milk of early to mid lactation dairy cows fed aflatoxin B1. J. Dairy Sci. 2009, 92, 3959–3963. [Google Scholar] [CrossRef] [PubMed]
- Sulzberger, S.A.; Melnichenko, S.; Cardoso, F.C. Effects of clay after an aflatoxin challenge on aflatoxin clearance, milk production, and metabolism of Holstein cows. J. Dairy Sci. 2017, 100, 1856–1869. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O.C.M.; Han, J.H.; Staples, C.R.; Adesogan, A.T. Effect of adding a mycotoxin-sequestering agent on milk aflatoxin M1 concentration and the performance and immune response of dairy cattle fed an aflatoxin B1-contaminated diet. J. Dairy Sci. 2012, 95, 5901–5908. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Palomba, M.; Mazzette, A.; Pulina, G. The transfer of aflatoxin M1 in milk of ewes fed diet naturally contaminated by aflatoxins and effect of inclusion of dried yeast culture in the diet. J. Dairy Sci. 2009, 92, 4997–5004. [Google Scholar] [CrossRef]
- Xiong, J.L.; Wang, Y.M.; Zhou, H.L.; Liu, J.X. Effects of dietary adsorbent on milk aflatoxin M1 content and the health of lactating dairy cows exposed to long-term aflatoxin B1 challenge. J. Dairy Sci. 2018, 101, 8944–8953. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Arriola, K.G.; Jiang, Y.; Driver, J.P.; Staples, C.R.; Adesogan, A.T. Effects of 3 sequestering agents on milk aflatoxin M1 concentration and the performance and immune status of dairy cows fed diets artificially contaminated with aflatoxin B1. J. Dairy Sci. 2016, 99, 6263–6273. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Palomba, M.; Pascale, M.; Nicolussi, P.; Pulina, G. Transfer of aflatoxin B1 from feed to milk and from milk to curd and whey in dairy sheep fed artificially contaminated concentrates. J. Dairy Sci. 2005, 88, 3063–3069. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Rassu, S.P.G.; Decandia, M.; Pulina, G. Excretion pattern of aflatoxin M1 in milk of goats fed a single dose of aflatoxin B1. J. Dairy Sci. 2012, 95, 2656–2661. [Google Scholar] [CrossRef]
- Moschini, M.; Masoero, F.; Gallo, A.; Diaz, D. Mucosal absorption of aflatoxin B1 in lactating dairy cows. Ital. J. Anim. Sci. 2007, 6, 324–326. [Google Scholar] [CrossRef]
- Masoero, F.; Gallo, A.; Moschini, M.; Piva, G.; Diaz, D. Carryover of aflatoxin from feed to milk in dairy cows with low or high somatic cell counts. Animal 2007, 1, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Van der Fels-Klerx, H.J.; Camenzuli, L. Effects of milk yield, feed composition, and feed contamination with aflatoxin B1 on the aflatoxin M1 concentration in dairy cows’ milk investigated using Monte Carlo Simulation Modelling. Toxins 2016, 8, 290. [Google Scholar] [CrossRef]
- Van Eijkeren, J.C.H.; Bakker, M.I.; Zeilmaker, M.J. A simple steady-state model for carry-over of aflatoxins from feed to cow’s milk. Food Addit. Contam. 2006, 23, 833–838. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, L.; Ji, C.; Li, X.; Jia, R.; Xi, L.; Zhang, J.; Ma, Q. Protective effects of Bacillus subtilis ANSB060 on serum biochemistry, histopathological changes and antioxidant enzyme activities of broilers fed moldy peanut meal naturally contaminated with aflatoxins. Toxins 2015, 7, 3330–3343. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle, 7th revised ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Li, X.; Zhao, L.; Fan, Y.; Jia, Y.; Sun, L.; Ma, S.; Ji, C.; Ma, Q.; Zhang, J. Occurrence of mycotoxins in feed ingredients and complete feeds obtained from the Beijing region of China. J. Anim. Sci. Biotechnol. 2014, 5, 37. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Item | Dietary Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | AF | AF + BDP | |||
| Milk yield (kg d−1) | 20.18 | 19.33 | 19.32 | 0.83 | 0.71 |
| Fat (%) | 3.43 | 3.34 | 3.37 | 0.18 | 0.94 |
| Protein (%) | 3.34 | 3.48 | 3.31 | 0.09 | 0.40 |
| Lactose (%) | 4.86 | 4.75 | 4.89 | 0.12 | 0.69 |
| MUN 2 (mg dL−1) | 11.38 | 12.07 | 11.88 | 0.53 | 0.65 |
| SCC 3 (log cells μL−1) | 5.37 | 5.70 | 5.50 | 0.26 | 0.67 |
| Item | Dietary Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | AF | AF + BDP | |||
| TP (g dL−1) | 7.10 | 7.41 | 7.23 | 0.21 | 0.57 |
| ALB (g dL−1) | 3.19 | 3.19 | 3.05 | 0.07 | 0.34 |
| AST (U L−1) | 89.63 | 92.13 | 83.63 | 3.09 | 0.16 |
| ALP (U L−1) | 33.13 | 34.13 | 32.38 | 1.88 | 0.81 |
| ALT (U L−1) | 42.50 | 43.88 | 39.75 | 1.84 | 0.29 |
| GGT (U L−1) | 39.38 | 39.88 | 37.13 | 2.89 | 0.78 |
| LDH (U L−1) | 945.88 b | 1106.75 a | 1032.88 ab | 36.52 | 0.02 |
| Item | Dietary Treatment 1 | SEM | p-Value | |
|---|---|---|---|---|
| AF | AF + BDP | |||
| AFM1 concentration (ng L−1) | 665 a | 483 b | 9 | <0.01 |
| AFM1 excretion 2 (μg d−1) | 12.71 a | 9.14 b | 1.58 | <0.01 |
| Transfer rate 3 (%) | 1.06 a | 0.76 b | 0.01 | <0.01 |
| Item | Amount |
|---|---|
| Ingredient composition (% of DM) | |
| Corn silage | 41.5 |
| Alfalfa silage | 14.9 |
| Flaked corn | 12.1 |
| Cottonseed meal | 4.9 |
| Distillers dried grains with soluble (DDGS) | 9.7 |
| Soybean meal (47% CP) | 2.5 |
| Corn gluten feed | 12.2 |
| Mineral and vitamin mix 1 | 2.2 |
| NEL (Mcal kg−1 of DM) | 1.51 |
| Chemical composition (% of DM) | |
| Crude protein (CP) | 15.5 |
| Fat | 3.5 |
| Neutral detergent fiber(NDF) | 42.6 |
| Acid detergent fiber(ADF) | 25.5 |
| Starch | 17.2 |
| Aflatoxins B1, B2, G1, and G2 | ND 2 |
| Deoxynivalenol | ND 2 |
| T-2 toxin | ND 2 |
| Zearalenone | ND 2 |
| Ochratoxin A | ND 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Zhang, Y.; Wei, C.; Ma, Q.; Ji, C.; Zhang, J.; Zhao, L. Efficacy of Bacillus subtilis ANSB060 Biodegradation Product for the Reduction of the Milk Aflatoxin M1 Content of Dairy Cows Exposed to Aflatoxin B1. Toxins 2019, 11, 161. https://doi.org/10.3390/toxins11030161
Guo Y, Zhang Y, Wei C, Ma Q, Ji C, Zhang J, Zhao L. Efficacy of Bacillus subtilis ANSB060 Biodegradation Product for the Reduction of the Milk Aflatoxin M1 Content of Dairy Cows Exposed to Aflatoxin B1. Toxins. 2019; 11(3):161. https://doi.org/10.3390/toxins11030161
Chicago/Turabian StyleGuo, Yongpeng, Yong Zhang, Chen Wei, Qiugang Ma, Cheng Ji, Jianyun Zhang, and Lihong Zhao. 2019. "Efficacy of Bacillus subtilis ANSB060 Biodegradation Product for the Reduction of the Milk Aflatoxin M1 Content of Dairy Cows Exposed to Aflatoxin B1" Toxins 11, no. 3: 161. https://doi.org/10.3390/toxins11030161
APA StyleGuo, Y., Zhang, Y., Wei, C., Ma, Q., Ji, C., Zhang, J., & Zhao, L. (2019). Efficacy of Bacillus subtilis ANSB060 Biodegradation Product for the Reduction of the Milk Aflatoxin M1 Content of Dairy Cows Exposed to Aflatoxin B1. Toxins, 11(3), 161. https://doi.org/10.3390/toxins11030161