Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons


 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Structure of the Genome of Nonreplicative HSV-1 Amplicon Vectors
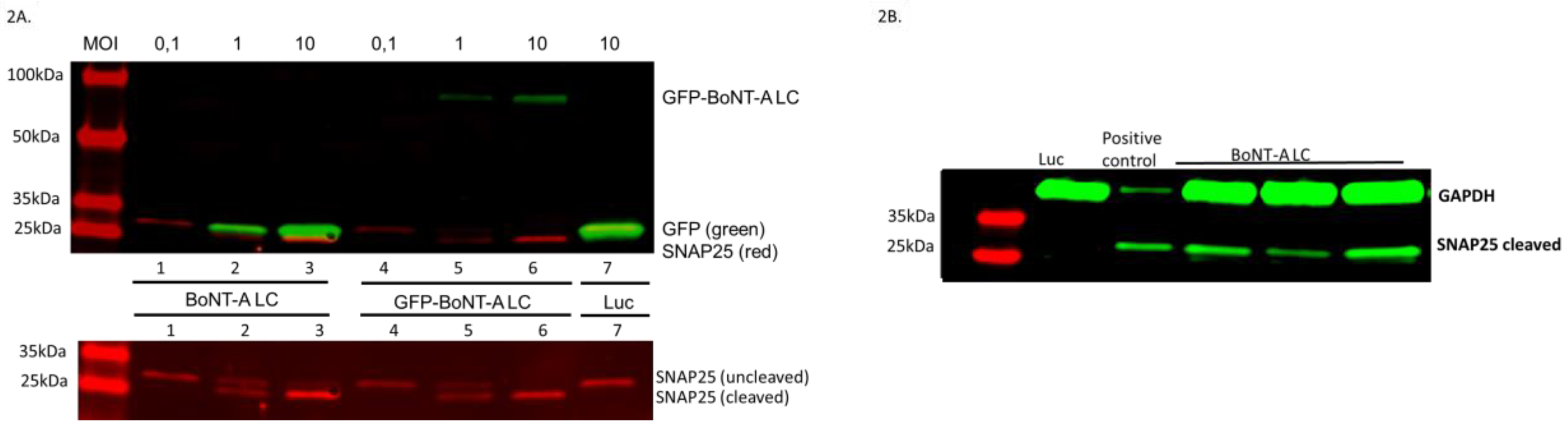
2.2. Vectors Expressing BoNT-A LC Cleave SNAP25 in Infected Neurons
2.3. Vectors Expressing BoNT-A LC Inhibit Secretion of CGRP Neuropeptide in Infected Neurons
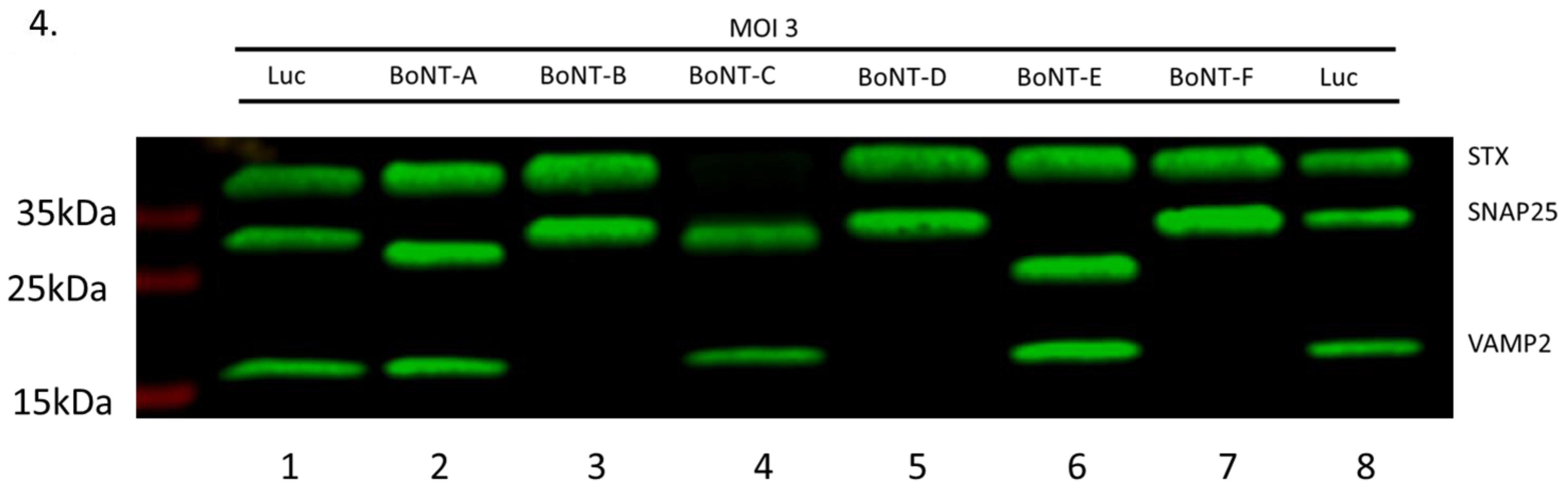
2.4. All BoNT LC Types Display Cleavage of Their Cognate SNARE Protein and Inhibition of CGRP Release
3. Discussion
4. Materials and Methods
4.1. Primary Cultures of Embryonic Rat Dorsal Root Ganglia (DRG) Neurons
4.2. Organotypic Cultures of Rat Adult DRG
4.3. Construction and Titration of Nonreplicative HSV-1 Amplicon Vectors
4.3.1. Plasmids Containing the LC of BoNTs
4.3.2. Generation of Amplicon Plasmids
4.3.3. Production and Titration of Amplicon Vector Particles
4.3.4. Infection Procedure
4.3.5. qRT-PCR
4.3.6. Western Blots (WB)
4.3.7. ELISA
4.4. Note on Biosafety and Biosecurity
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Simpson, L.L. The origin, structure, and pharmacological activity of botulinum toxin. Pharmacol. Rev. 1981, 33, 155–188. [Google Scholar] [PubMed]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum neurotoxins: Biology, pharmacology and toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef]
- Chaddock, J.A.; Herbert, M.H.; Ling, R.J.; Alexander, F.C.; Fooks, S.J.; Revell, D.F.; Quinn, C.P.; Shone, C.C.; Foster, K.A. Expression and purification of catalytically active, non-toxic endopeptidase derivatives of Clostridium botulinum toxin type A. Protein Expr. Purif. 2002, 25, 219–228. [Google Scholar] [CrossRef]
- Jahn, R.; Scheller, R.H. SNAREs--engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Blasi, J.; Chapman, E.R.; Link, E.; Binz, T.; Yamasaki, S.; De Camilli, P.; Sudhof, T.C.; Niemann, H.; Jahn, R. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature 1993, 365, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G. Mechanism of action of tetanus and botulinum neurotoxins. Mol. Microbiol. 1994, 13, 1–8. [Google Scholar] [CrossRef]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef]
- Chen, Y.A.; Scheller, R.H. SNARE-mediated membrane fusion. Nat. Rev. Mol. Cell Biol. 2001, 2, 98–106. [Google Scholar] [CrossRef]
- Humeau, Y.; Doussau, F.; Grant, N.J.; Poulain, B. How botulinum and tetanus neurotoxins block neurotransmitter release. Biochimie 2000, 82, 427–446. [Google Scholar] [CrossRef]
- Schiavo, G.; Benfenati, F.; Poulain, B.; Rossetto, O.; Polverino de Laureto, P.; DasGupta, B.R.; Montecucco, C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992, 359, 832–835. [Google Scholar] [CrossRef]
- Ahnert-Hilger, G.; Weller, U. Comparison of the intracellular effects of clostridial neurotoxins on exocytosis from streptolysin O-permeabilized rat pheochromocytoma (PC 12) and bovine adrenal chromaffin cells. Neuroscience 1993, 53, 547–552. [Google Scholar] [CrossRef]
- Boyd, R.S.; Duggan, M.J.; Shone, C.C.; Foster, K.A. The effect of botulinum neurotoxins on the release of insulin from the insulinoma cell lines HIT-15 and RINm5F. J. Biol Chem. 1995, 270, 18216–18218. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Schiavo, G.; Tugnoli, V.; de Grandis, D. Botulinum neurotoxins: Mechanism of action and therapeutic applications. Mol. Med. Today 1996, 2, 418–424. [Google Scholar] [CrossRef]
- Welch, M.; Purkiss, J.R.; Foster, K. Sensitivity of embryonic rat dorsal root ganglia neurons to Clostridium botulinum neurotoxins. Toxicon 2000, 38, 245–258. [Google Scholar] [CrossRef]
- Poulain, B.; Mochida, S.; Weller, U.; Hogy, B.; Habermann, E.; Wadsworth, J.D.; Shone, C.C.; Dolly, J.O.; Tauc, L. Heterologous combinations of heavy and light chains from botulinum neurotoxin A and tetanus toxin inhibit neurotransmitter release in Aplysia. J. Biol. Chem. 1991, 266, 9580–9585. [Google Scholar] [PubMed]
- Lawrence, G.W.; Foran, P.; Dolly, J.O. Distinct exocytotic responses of intact and permeabilised chromaffin cells after cleavage of the 25-kDa synaptosomal-associated protein (SNAP-25) or synaptobrevin by botulinum toxin A or B. Eur. J. Biochem. 1996, 236, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Tamori, Y.; Hashiramoto, M.; Araki, S.; Kamata, Y.; Takahashi, M.; Kozaki, S.; Kasuga, M. Cleavage of vesicle-associated membrane protein (VAMP)-2 and cellubrevin on GLUT4-containing vesicles inhibits the translocation of GLUT4 in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 1996, 220, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Teng, Q.; Tanase, D.K.; Liu, J.K.; Garrity-Moses, M.E.; Baker, K.B.; Boulis, N.M. Adenoviral clostridial light chain gene-based synaptic inhibition through neuronal synaptobrevin elimination. Gene Ther. 2005, 12, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Krishnaney, A.; Teng, Q.; Yang, J.; Garrity-Moses, M.; Liu, J.K.; Venkiteswaran, K.; Subramanian, Y.; Davis, M.; Boulis, N.M. Anatomically discrete functional effects of adenoviral clostridial light chain gene-based synaptic inhibition in the midbrain. Gene Ther. 2006, 13, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Oehmig, A.; Fraefel, C.; Breakefield, X.O. Update on herpesvirus amplicon vectors. Mol. Ther. 2004, 10, 630–643. [Google Scholar] [CrossRef]
- Marconi, P.; Manservigi, R.; Epstein, A.L. HSV-1-derived helper-independent defective vectors, replicating vectors and amplicon vectors, for the treatment of brain diseases. Curr. Opin. Drug Discov. Devel. 2010, 13, 169–183. [Google Scholar] [PubMed]
- Wilson, S.P.; Yeomans, D.C.; Bender, M.A.; Lu, Y.; Goins, W.F.; Glorioso, J.C. Antihyperalgesic effects of infection with a preproenkephalin-encoding herpes virus. Proc. Natl. Acad. Sci. USA. 1999, 96, 3211–3216. [Google Scholar] [CrossRef] [PubMed]
- Decker, H.; Jürgensen, S.; Adrover, M.F.; Brito-Moreira, J.; Bomfim, T.R.; Klein, W.L.; Epstein, A.L.; De Felice, F.G.; Jerusalinsky, D.; Ferreira, S.T. N-methyl-D-aspartate receptors are required for synaptic targeting of Alzheimer’s toxic amyloid-β peptide oligomers. J. Neurochem. 2010, 115, 1520–1529. [Google Scholar] [CrossRef] [PubMed]
- Foka, P.; Pourchet, A.; Hernandez-Alcoceba, R.; Doumba, P.P.; Pissas, G.; Kouvatsis, V.; Dalagiorgou, G.; Kazazi, D.; Marconi, P.; Foschini, M.G.; et al. Novel tumour-specific promoters for transcriptional targeting of hepatocellular carcinoma by herpes simplex virus vectors. J. Gene Med. 2010, 12, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Gayral, M.; Lulka, H.; Hanoun, N.; Biollay, C.; Sèlves, J.; Berthommé, H.; Trempat, P.; Epstein, A.; Buscail, L.; Béjot, J.L.; et al. Targeted oncolytic herpes simplex virus type 1 eradicates experimental pancreatic tumors. Hum. Gene Ther. 2015, 26, 104–113. [Google Scholar] [CrossRef]
- Ferrera, R.; Cuchet, D.; Zaupa, C.; Michel, P.; Revol-Guyot, V.; Ovize, M.; Epstein, A.L. Efficient and non-toxic gene transfer to cardiomyocytes using novel generation amplicon vectors derived from HSV-1. J. Mol. Cell Cardiol. 2005, 38, 219–223. [Google Scholar] [CrossRef]
- Tsui, J.K.; Eisen, A.; Stoessl, A.J.; Calne, S.; Calne, D.B. Double-blind study of botulinum toxin in spasmodic torticollis. Lancet 1986, 2, 245–247. [Google Scholar] [CrossRef]
- Truong, D.; Duane, D.D.; Jankovic, J.; Singer, C.; Seeberger, L.C.; Comella, C.L.; Lew, M.F.; Rodnitzky, R.L.; Danisi, F.O.; Sutton, J.P.; et al. Efficacy and safety of botulinum type A toxin (Dysport) in cervical dystonia: Results of the first US randomized, double-blind, placebo-controlled study. Mov. Disord. 2005, 20, 783–791. [Google Scholar] [CrossRef]
- Jankovic, J.; Comella, C.; Hanschmann, A.; Grafe, S. Efficacy and safety of incobotulinumtoxinA (NT 201, Xeomin) in the treatment of blepharospasm-a randomized trial. Mov. Disord. 2011, 26, 1521–1528. [Google Scholar] [CrossRef]
- Wabbels, B.; Reichel, G.; Fulford-Smith, A.; Wright, N.; Roggenkämper, P. Double-blind, randomised, parallel group pilot study comparing two botulinum toxin type A products for the treatment of blepharospasm. J. Neural. Transm (Vienna). 2011, 118, 233–239. [Google Scholar] [CrossRef]
- Bakheit, A.M.; Pittock, S.; Moore, A.P.; Wurker, M.; Otto, S.; Erbguth, F.; Coxon, L. A randomized, double-blind, placebo-controlled study of the efficacy and safety of botulinum toxin type A in upper limb spasticity in patients with stroke. Eur. J. Neurol. 2001, 8, 559–565. [Google Scholar] [CrossRef]
- Childers, M.K.; Brashear, A.; Jozefczyk, P.; Reding, M.; Alexander, D.; Good, D.; Walcott, J.M.; Jenkins, S.W.; Turkel, C.; Molloy, P.D. Dose-dependent response to intramuscular botulinum toxin type A for upper-limb spasticity in patients after a stroke. Arch. Phys. Med. Rehabil. 2004, 85, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Binder, W.J.; Brin, M.F.; Blitzer, A.; Schoenrock, L.D.; Pogoda, J.M. Botulinum toxin type A (BOTOX) for treatment of migraine headaches: An open-label study. Otolaryngol. Head Neck Surg. 2000, 123, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Escher, C.M.; Paracka, L.; Dressler, D.; Kollewe, K. Botulinum toxin in the management of chronic migraine: Clinical evidence and experience. Ther. Adv. Neurol. Disord. 2017, 10, 127–135. [Google Scholar] [CrossRef]
- Brubaker, L.; Richter, H.E.; Visco, A.; Mahajan, S.; Nygaard, I.; Braun, T.M.; Barber, M.D.; Menefee, S.; Schaffer, J.; Weber, A.M.; et al. Refractory idiopathic urge urinary incontinence and botulinum A injection. J. Urol. 2008, 180, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Nitti, V.W.; Dmochowski, R.; Herschorn, S.; Sand, P.; Thompson, C.; Nardo, C.; Yan, X.; Haag-Molkenteller, C.; EMBARK Study Group. OnabotulinumtoxinA for the treatment of patients with overactive bladder and urinary incontinence: Results of a phase 3, randomized, placebo controlled trial. J. Urol. 2013, 189, 2186–2193. [Google Scholar] [CrossRef] [PubMed]
- Schurch, B.; de Sèze, M.; Denys, P.; Chartier-Kastler, E.; Haab, F.; Everaert, K.; Plante, P.; Perrouin-Verbe, B.; Kumar, C.; Fraczek, S.; et al. Botulinum toxin type a is a safe and effective treatment for neurogenic urinary incontinence: Results of a single treatment, randomized, placebo controlled 6-month study. J. Urol. 2005, 174, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.; Herschorn, S.; Aliotta, P.; Brin, M.; Thompson, C.; Lam, W.; Daniell, G.; Heesakkers, J.; Haag-Molkenteller, C. Efficacy and safety of onabotulinumtoxinA in patients with urinary incontinence due to neurogenic detrusor overactivity: A randomised, double-blind, placebo-controlled trial. Eur. Urol. 2011, 60, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, P.J.; Ravich, W.J.; Hendrix, T.R.; Sostre, S.; Jones, B.; Kalloo, A.N. Intrasphincteric botulinum toxin for the treatment of achalasia. N. Engl. J. Med. 1995, 332, 774–778. [Google Scholar] [CrossRef]
- Jung, H.E.; Lee, J.S.; Lee, T.H.; Kim, J.N.; Hong, S.J.; Kim, J.O.; Kim, H.G.; Jeon, S.R.; Cho, J.Y. Long-term outcomes of balloon dilation versus botulinum toxin injection in patients with primary achalasia. Korean J. Intern. Med. 2014, 29, 738–745. [Google Scholar] [CrossRef]
- Tejedor, J.; Rodríguez, J.M. Early retreatment of infantile esotropia: Comparison of reoperation and botulinum toxin. Br. J. Ophthalmol. 1999, 83, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, J.D.; Kennedy, R.A.; Bagaric, D. Botulinum vs. adjustable suture surgery in the treatment of horizontal misalignment in adult patients lacking fusion. Arch. Ophthalmol. 1990, 108, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Foster, L.; Clapp, L.; Erickson, M.; Jabbari, B. Botulinum toxin A and chronic low back pain: A randomized, double-blind study. Neurology 2001, 56, 1290–1293. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, Y.; Lee, J.; Park, C.; Moon, D.E. The effects of botulinum toxin A on mechanical and cold allodynia in a rat model of neuropathic pain. Can. J. Anaesth. 2006, 53, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Hou, L.; Wang, X. Comparison of calcitonin gene-related peptide release from rat lymphocytes and dorsal root ganglia neurons. Brain Behav. Immun. 2002, 16, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Foran, P.G.; Mohammed, N.; Lisk, G.O.; Nagwaney, S.; Lawrence, G.W.; Johnson, E.; Smith, L.; Aoki, K.R.; Dolly, J.O. Evaluation of the therapeutic usefulness of botulinum neurotoxin B, C1, E and F compared with the long lasting type A. J. Biol. Chem. 2003, 278, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.P.; Barbieri, J.T. Light Chain Diversity among the Botulinum Neurotoxins. Toxins 2018, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Donald, S.; Elliott, M.; Gray, B.; Hornby, F.; Lewandowska, A.; Marlin, S.; Favre-Guilmard, C.; Périer, C.; Cornet, S.; Kalinichev, M.; et al. A comparison of biological activity of commercially available purified native botulinum neurotoxin serotypes A1 to F1 in vitro, ex vivo, and in vivo. Pharmacol. Res. Perspect. 2018, 6, e00446. [Google Scholar] [CrossRef]
- Fonfria, E.; Maignel, J.; Lezmi, S.; Martin, V.; Splevins, A.; Shubber, S.; Kalinichev, M.; Foster, K.; Picaut, P.; Krupp, J. The Expanding Therapeutic Utility of Botulinum Neurotoxins. Toxin 2018, 10, 208. [Google Scholar] [CrossRef]
- Joussain, C.; Popoff, M.; Phé, V.; Even, A.; Bosset, P.-O.; Pottier, S.; Falcou, L.; Levy, J.; Vaugier, I.; Chartier Kastler, E.; et al. Long-term outcomes and risks factors for failure of intradetrusor onabotulinumtoxin A injections for the treatment of refractory neurogenic detrusor overactivity. Neurourol Urodyn. 2018, 37, 799–806. [Google Scholar] [CrossRef]
- Cuchet, D.; Ferrera, R.; Lomonte, P.; Epstein, A.L. Characterisation of the antiproliferative and cytotoxic properties of the HSV-1 immediate-early ICP0 protein. J. Gene Med. 2005, 7, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Zaupa, C.; Revol-Guyot, V.; Epstein, A.L. Improved packaging system for generation of high levels non-cytotoxic HSV-1 amplicon vectors using Cre-loxP site-specific recombination to delete the packaging signals of defective helper genomes. Hum. Gene Ther. 2003, 14, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Krisky, D.M.; Wolfe, D.; Goins, W.F.; Marconi, P.C.; Ramakrishnan, R.; Mata, M.; Rouse, R.J.; Fink, D.J.; Glorioso, J.C. Deletion of multiple immediate–early genes from herpes simplex virus reduces cytotoxicity and permits long-term gene expression in neurons. Gene Ther. 1998, 5, 1593–1603. [Google Scholar] [CrossRef]
- Casadevall, A.; Dermody, T.S.; Imperiale, M.J.; Sandri-Goldin, R.M.; Shenk, T. Dual-use research of concern (DURC) review at American Society for Microbiology Journals. MBio 2015, 6, e01236. [Google Scholar] [CrossRef] [PubMed]
- Fink, D.J.; Wechuck, J.; Mata, M.; Glorioso, J.C.; Goss, J.; Krisky, D.; Wolfe, D. Gene therapy for pain: Results of a Phase I clinical trial. Ann. Neurol. 2011, 70, 207–212. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Oligonucleotide | Sequence of Oligonucleotide |
|---|---|
| RT-BoNT-A F1 | GCGCCGACATCATCCAGTTC |
| RT-BoNT-A R1 | GTGTCCACCTCCAGGCTCTC |
| Rat.GAPDH-1049F | GTGGACCTCATGGCCTACAT |
| Rat.GAPDH-1190R | TGTGAGGGAGATGCTCAGTG |
| Primary Antibodies | Brand | Reference | Species |
| STX | ABCAM | ab3265 | Rabbit |
| SNAP25 | ABCAM | ab66066 | Rabbit |
| VAMP2 | ABCAM | ab3347 | Rabbit |
| SNAP25 | IPSEN | Rabbit | |
| GAPDH | Millipore | CB1001 | Mouse |
| GFP | Life technologies | A6455 | Rabbit |
| Secondary Antibodies | Brand | Reference | Species |
| Mouse (800) | Li-Cor | 926-32210 | Goat |
| Rabbit (680) | Li-Cor | 926-68071 | Goat |
| Rabbit (800) | Li-Cor | 926-32211 | Goat |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joussain, C.; Le Coz, O.; Pichugin, A.; Marconi, P.; Lim, F.; Sicurella, M.; Salonia, A.; Montorsi, F.; Wandosell, F.; Foster, K.; et al. Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons. Toxins 2019, 11, 123. https://doi.org/10.3390/toxins11020123
Joussain C, Le Coz O, Pichugin A, Marconi P, Lim F, Sicurella M, Salonia A, Montorsi F, Wandosell F, Foster K, et al. Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons. Toxins. 2019; 11(2):123. https://doi.org/10.3390/toxins11020123
Chicago/Turabian StyleJoussain, Charles, Olivier Le Coz, Andrey Pichugin, Peggy Marconi, Filip Lim, Mariaconcetta Sicurella, Andrea Salonia, Francesco Montorsi, Francisco Wandosell, Keith Foster, and et al. 2019. "Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons" Toxins 11, no. 2: 123. https://doi.org/10.3390/toxins11020123
APA StyleJoussain, C., Le Coz, O., Pichugin, A., Marconi, P., Lim, F., Sicurella, M., Salonia, A., Montorsi, F., Wandosell, F., Foster, K., Giuliano, F., Epstein, A. L., & Aranda Muñoz, A. (2019). Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons. Toxins, 11(2), 123. https://doi.org/10.3390/toxins11020123