Candidacidal Activity of a Novel Killer Toxin from Wickerhamomyces anomalus against Fluconazole-Susceptible and -Resistant Strains

 ,
, 

 ,
, 

Abstract
1. Introduction
2. Results
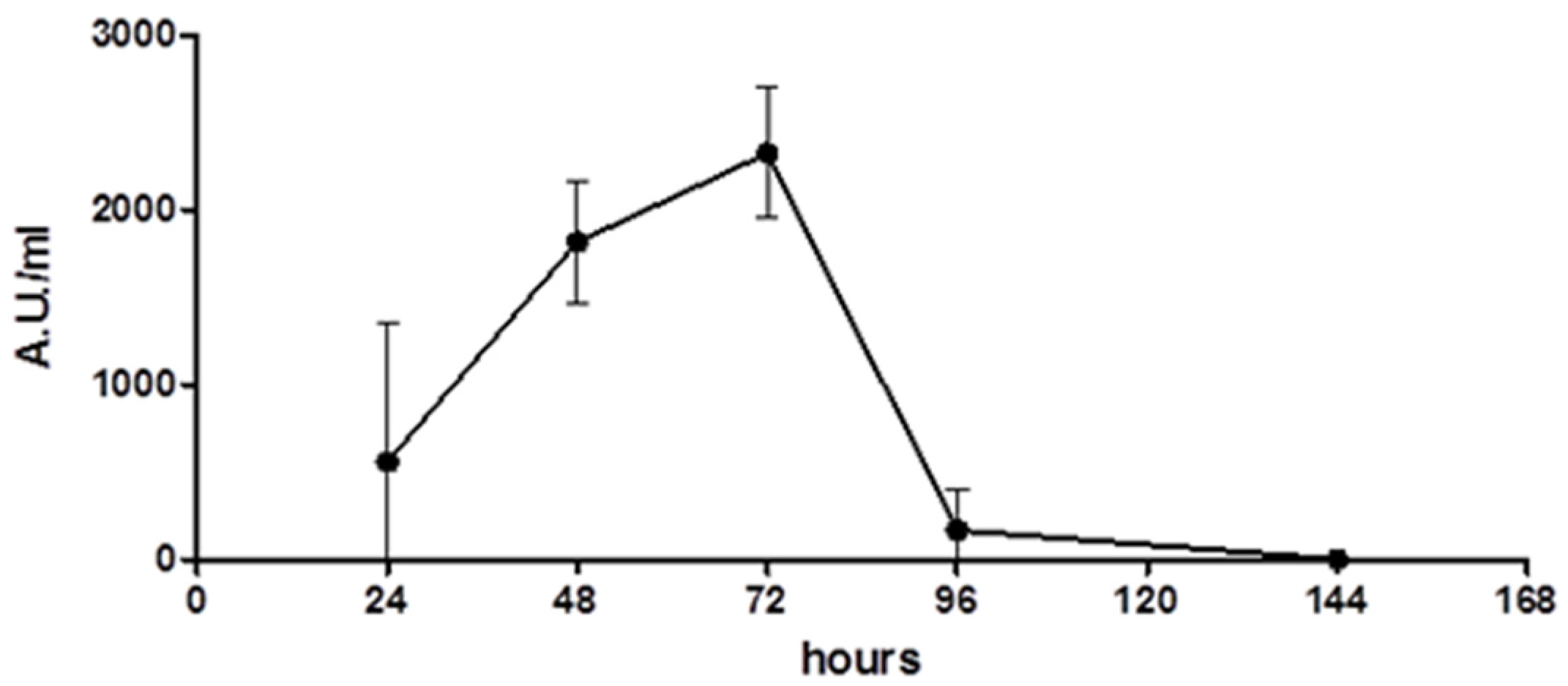
2.1. Wa1F1-KT Production
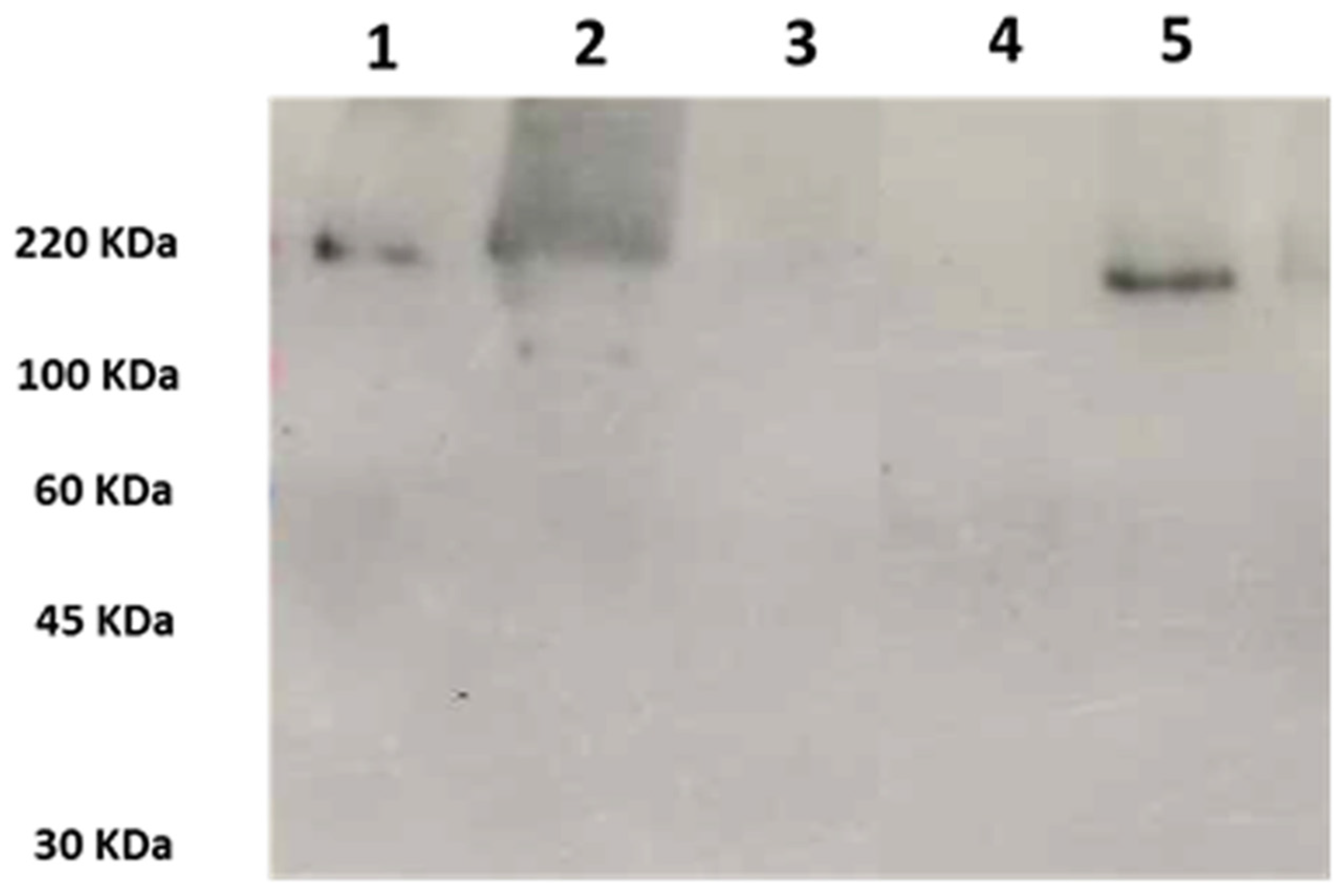
2.2. Western Blot Analysis of Wa1F1-KT
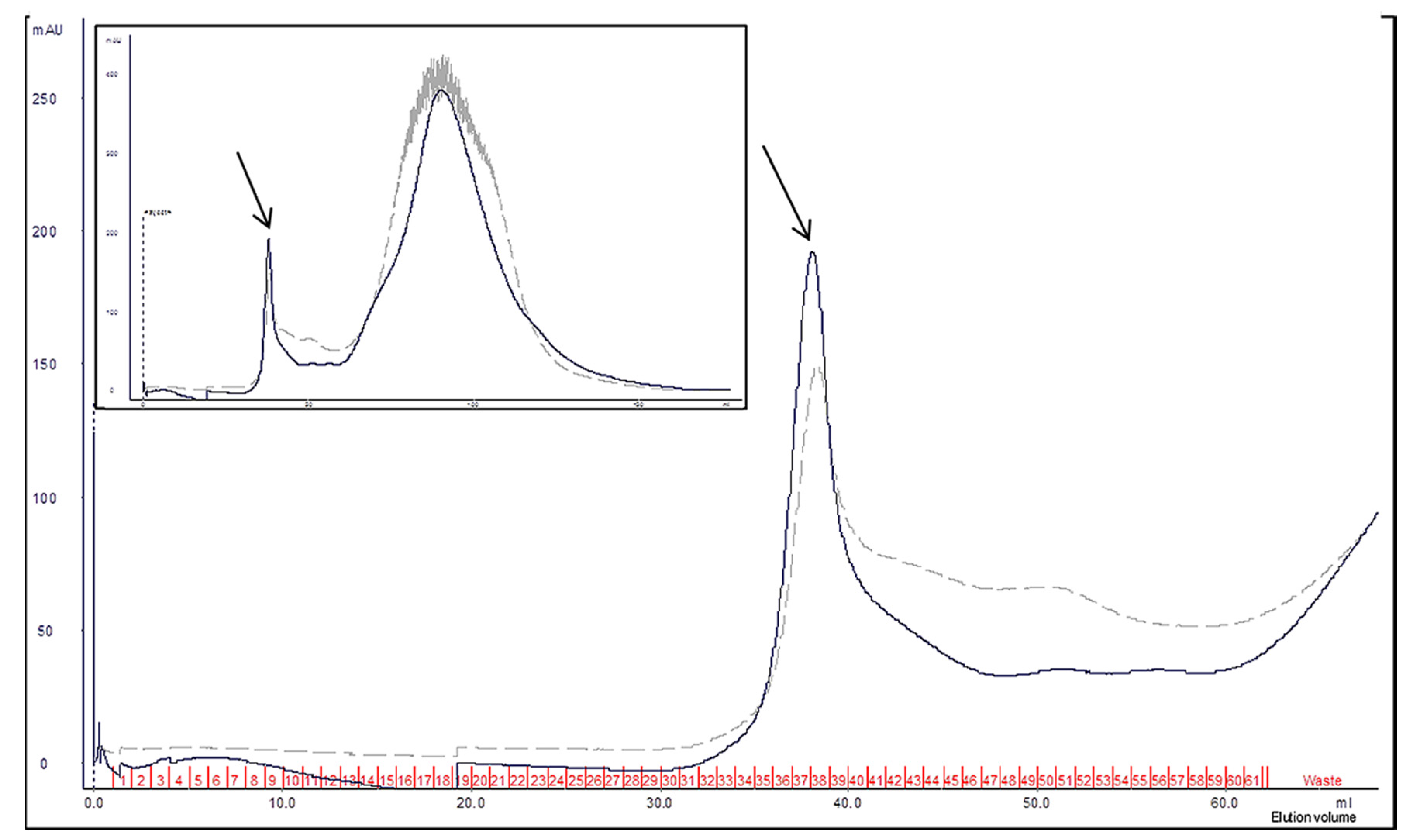
2.3. Characterization of Wa1F1-KT Using Size Exclusion Chromatography
2.4. Activity of Wa1F1-KT
2.5. Exo-β-1,3-Glucanase Activity
3. Discussion
4. Materials and Methods
4.1. Yeast Strains
4.2. Media
4.3. Production of KTs
4.4. Western Blot Analysis
4.5. Characterization of Wa1F1-KT Using Size Exclusion Chromatography
4.6. Evaluation of Wa1F1-KT Activity
4.7. Laminarin Hydrolysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bevan, E.A.; Makower, M. The Physiological Basis of the Killer Character in Yeast; XIth Int Congr Genet, Leiden, The Netherlands; Pergamon Press: Oxford, England, 1963; pp. 202–203. [Google Scholar]
- Magliani, W.; Conti, S.; Gerloni, M.; Bertolotti, D.; Polonelli, L. Yeast killer systems. Clin. Microbiol. Rev. 1997, 10, 369–400. [Google Scholar] [PubMed]
- Muccilli, S.; Restuccia, C. Bioprotective role of yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [PubMed]
- Buzzini, P.; Turchetti, B.; Vaughan-Martini, A.E. The use of killer sensitivity patterns for biotyping yeast strains: The state of the art, potentialities and limitations. FEMS Yeast Res. 2007, 7, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Conti, S. Biotyping of Candida albicans and other fungi by yeast killer toxins sensitivity. Methods Mol. Biol. 2009, 499, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Magliani, W.; Ciociola, T.; Giovati, L.; Conti, S. From Pichia anomala killer toxin through killer antibodies to killer peptides for a comprehensive anti-infective strategy. Antonie Van Leeuwenhoek 2011, 99, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.M. Pichia anomala: Cell physiology and biotechnology relative to other yeasts. Antonie Van Leeuwenhoek 2011, 99, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Valzano, M.; Cecarini, V.; Cappelli, A.; Capone, A.; Bozic, J.; Cuccioloni, M.; Epis, S.; Petrelli, D.; Angeletti, M.; Eleuteri, A.M.; et al. A yeast strain associated to Anopheles mosquitoes produces a toxin able to kill malaria parasites. Malaria J. 2016, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Kagiyama, S.; Aiba, T.; Kadowaki, K.; Mogi, K. New killer toxins of halophilic Hansenula anomala. Agric. Biol. Chem. 1988, 52, 1–7. [Google Scholar] [CrossRef]
- Fredlund, E.; Druvefors, U.; Boysen, M.E.; Lingsten, K.-J.; Schnürer, J. Physiological characteristics of the biocontrol yeast Pichia anomala J121. FEMS Yeast Res. 2002, 2, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.F.; Contreras, L.; Garnica, N.M.; Fernández-Zenoff, M.V.; Farías, M.E.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Native killer yeasts as biocontrol agents of postharvest fungal diseases in lemons. PLoS ONE 2016, 11, e0165590. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chi, Z.; Yue, L.; Li, J.; Li, M.; Wu, L. A marine killer yeast against the pathogenic yeast strain in crab (Portunus trituberculatus) and an optimization of the toxin production. Microbiol. Res. 2007, 162, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Ricci, I.; Damiani, C.; Scuppa, P.; Mosca, M.; Crotti, E.; Rossi, P.; Rizzi, A.; Capone, A.; Gonella, E.; Ballarini, P.; et al. The yeast Wickerhamomyces anomalus (Pichia anomala) inhabits the midgut and reproductive system of the Asian malaria vector Anopheles stephensi. Environ. Microbiol. 2011, 13, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, A.; Ulissi, U.; Valzano, M.; Damiani, C.; Epis, S.; Gabrielli, M.G.; Conti, S.; Polonelli, L.; Bandi, C.; Favia, G.; et al. A Wickerhamomyces anomalus killer strain in the malaria vector Anopheles stephensi. PLoS ONE 2014, 9, e95988. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Bongiorno, G.; Giovati, L.; Montagna, M.; Crotti, E.; Damiani, C.; Gradoni, L.; Polonelli, L.; Ricci, I.; Favia, G.; et al. Isolation of a Wickerhamomyces anomalus yeast strain from the sandfly Phlebotomus perniciosus, displaying the killer phenotype. Med. Vet. Entomol. 2016, 30, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Passoth, V.; Fredlund, E.; Druvefors, U.Ä.; Schnürer, J. Biotechnology, physiology and genetics of the yeast Pichia anomala. FEMS Yeast Res. 2006, 6, 3–13. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S. Risk assessment of fungi reported from humans and animals. Mycoses 1996, 39, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Epis, S.; Capone, A.; Martin, E.; Paolucci, M.; Bazzocchi, C.; Valzano, M.; Bozic, J.; Novati, S.; Favia, G.; Ricci, I. A rapid qPCR method to investigate the circulation of the yeast Wickerhamomyces anomalus in humans. New Microbiol. 2015, 38, 577–581. [Google Scholar] [PubMed]
- İzgü, F.; Altınbay, D.; Türeli, A.E. In vitro activity of panomycocin, a novel exo-β-1,3-glucanase isolated from Pichia anomala NCYC 434, against dermatophytes. Mycoses 2007, 50, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Tay, S.-T.; Lim, S.-L.; Tan, H.-W. Growth inhibition of Candida species by Wickerhamomyces anomalus mycocin and a lactone compound of Aureobasidium pullulans. BMC Complement. Altern. Med. 2014, 14, 439. [Google Scholar] [CrossRef] [PubMed]
- Paris, A.P.; Persel, C.; Serafin, C.F.; de Cassia Garcia Simao, R.; Gandra, R.F. Susceptibility of Candida albicans isolated from blood to Wickerhamomyces anomalous mycocins. Curr. Microbiol. 2016, 73, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The changing epidemiology of invasive fungal infections. In Human Fungal Pathogen Identification: Methods and Protocols; Lion, T., Ed.; Springer: New York, NY, USA, 2017; pp. 17–65. [Google Scholar]
- Guinea, J. Global trends in the distribution of Candida species causing candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R. Current epidemiology of Candida infection. Clin. Microbiol. Newsl. 2014, 36, 131–136. [Google Scholar] [CrossRef]
- Orasch, C.; Marchetti, O.; Garbino, J.; Schrenzel, J.; Zimmerli, S.; Mühlethaler, K.; Pfyffer, G.; Ruef, C.; Fehr, J.; Zbinden, R.; et al. Candida species distribution and antifungal susceptibility testing according to European Committee on Antimicrobial Susceptibility Testing and new vs. old Clinical and Laboratory Standards Institute clinical breakpoints: A 6-year prospective candidaemia survey from the fungal infection network of Switzerland. Clin. Microbiol. Infect. 2014, 20, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Hsu, L.Y.; Alejandria, M.M.; Chaiwarith, R.; Chinniah, T.; Chayakulkeeree, M.; Choudhury, S.; Chen, Y.H.; Shin, J.H.; Kiratisin, P.; et al. Antifungal susceptibility of invasive Candida bloodstream isolates from the Asia-Pacific region. Med. Mycol. 2016, 54, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Kuchler, K.; Ischer, F.; Pagani, J.L.; Monod, M.; Bille, J. Mechanisms of resistance to azole antifungal agents in Candida albicans isolates from AIDS patients involve specific multidrug transporters. Antimicrob. Agents Chemother. 1995, 39, 2378–2386. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Calabrese, D.; Majcherczyk, P.A.; Bille, J. The ATP binding cassette transporter gene CgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 1999, 43, 2753–2765. [Google Scholar] [PubMed]
- Coste, A.; Selmecki, A.; Forche, A.; Diogo, D.; Bougnoux, M.-E.; d’Enfert, C.; Berman, J.; Sanglard, D. Genotypic evolution of azole resistance mechanisms in sequential Candida albicans isolates. Eukaryot. Cell 2007, 6, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.T.; Crittin, J.; Bauser, C.; Rohde, B.; Sanglard, D. Functional analysis of cis- and trans-acting elements of the Candida albicans CDR2 promoter with a novel promoter reporter system. Eukaryot. Cell 2009, 8, 1250–1267. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Ischer, F.; Calabrese, D.; Posteraro, B.; Sanguinetti, M.; Fadda, G.; Rohde, B.; Bauser, C.; Bader, O.; Sanglard, D. Gain of function mutations in CgPDR1 of Candida glabrata not only mediate antifungal resistance but also enhance virulence. PLoS Pathog. 2009, 5, e1000268. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Séguy, N.; Conti, S.; Gerloni, M.; Bertolotti, D.; Cantelli, C.; Magliani, W.; Cailliez, J.C. Monoclonal yeast killer toxin-like candidacidal anti-idiotypic antibodies. Clin. Diagn. Lab. Immunol. 1997, 4, 142–146. [Google Scholar] [PubMed]
- Guyard, C.; Evrard, P.; Corbisier-Colson, A.M.; Louvart, H.; Dei-Cas, E.; Menozzi, F.D.; Polonelli, L.; Cailliez, J. Immuno-crossreactivity of an anti-Pichia anomala killer toxin monoclonal antibody with a Williopsis saturnus var. mrakii killer toxin. Med. Mycol. 2001, 39, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Guyard, C.; Dehecq, E.; Tissier, J.-P.; Polonelli, L.; Dei-Cas, E.; Cailliez, J.-C.; Menozzi, F.D. Involvement of beta-glucans in the wide-spectrum antimicrobial activity of Williopsis saturnus var. mrakii MUCL 41968 killer toxin. Mol. Med. 2002, 8, 686–694. [Google Scholar] [PubMed]
- López-Martínez, R. Candidosis, a new challenge. Clin. Dermatol. 2010, 28, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, C.; Leonardi, D. Candida infections, causes, targets, and resistance mechanisms: Traditional and alternative antifungal agents. Biomed. Res. Int. 2013, 2013, 204237. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Archibusacci, C.; Sestito, M.; Morace, G. Killer system: A simple method for differentiating Candida albicans strains. J. Clin. Microbiol. 1983, 17, 774–780. [Google Scholar] [PubMed]
- Polonelli, L.; Conti, S.; Gerloni, M.; Campani, L.; Mantovani, M.P.; Morace, G. Production of yeast killer toxin in experimentally infected animals. Mycopathologia 1990, 110, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.-J.; Ma, Y.; Xu, H.-M.; Wang, X.-H.; Chi, Z.-M. A novel killer toxin produced by the marine-derived yeast Wickerhamomyces anomalus YF07b. Antonie Van Leeuwenhoek 2013, 103, 737–746. [Google Scholar] [CrossRef] [PubMed]
- İzgü, F.; Altınbay, D. Isolation and characterization of the K5-type yeast killer protein and its homology with an exo-beta-1,3-glucanase. Biosci. Biotechnol. Biochem. 2004, 68, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yue, L.; Chi, Z.; Wang, X. Marine killer yeasts active against a yeast strain pathogenic to crab Portunus trituberculatus. Dis. Aquat. Organ. 2008, 80, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Mathews, H.L.; Conti, S.; Witek-Janusek, L.; Polonelli, L. Effect of Pichia anomala killer toxin on Candida albicans. Med. Mycol. 1998, 36, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Beninati, C.; Teti, G.; Felici, F.; Ciociola, T.; Giovati, L.; Sperindè, M.; Passo, C.L.; Pernice, I.; Domina, M.; et al. Yeast killer toxin-like candidacidal Ab6 antibodies elicited through the manipulation of the idiotypic cascade. PLoS ONE 2014, 9, e105727. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P.; Sullivan, P.A.; Shepherd, M.G. Isolation and structure of glucan from regenerating spheroplasts of Candida albicans. Microbiology 1984, 130, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- İzgü, F.; Altınbay, D.; Acun, T. Killer toxin of Pichia anomala NCYC 432; purification, characterization and its exo-β-1,3-glucanase activity. Enzyme Microb. Technol. 2006, 39, 669–676. [Google Scholar] [CrossRef]
- Chi, Z.; Chi, Z.; Zhang, T.; Liu, G.; Li, J.; Wang, X. Production, characterization and gene cloning of the extracellular enzymes from the marine-derived yeasts and their potential applications. Biotechnol. Adv. 2009, 27, 236–255. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Lorenzini, R.; De Bernardis, F.; Morace, G. Potential therapeutic effect of yeast killer toxin. Mycopathologia 1986, 96, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Franz, R.; Kelly, S.L.; Lamb, D.C.; Kelly, D.E.; Ruhnke, M.; Morschhäuser, J. Multiple molecular mechanisms contribute to a stepwise development of fluconazole resistance in clinical Candida albicans strains. Antimicrob. Agents Chemother. 1998, 42, 3065–3072. [Google Scholar] [PubMed]
- Kanafani, Z.A.; Perfect, J.R. Resistance to antifungal agents: Mechanisms and clinical impact. Clin. Infect. Dis. 2008, 46, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sheng, F.; Zhao, J.; Chen, L.; Li, C. ERG11 mutations and expression of resistance genes in fluconazole-resistant Candida albicans isolates. Arch. Microbiol. 2015, 197, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.T.; Karababa, M.; Ischer, F.; Bille, J.; Sanglard, D. TAC1, transcriptional activator of CDR genes, is a new transcription factor involved in the regulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 2004, 3, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Sheng, J.; Chi, Z.; Li, J. Inulinase production by a marine yeast Pichia guilliermondii and inulin hydrolysis by the crude inulinase. J. Ind. Microbiol. Biotechnol. 2007, 34, 179–185. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Features and Genotype | Wa1F1-KT Sensitivity (mm Growth Inhibition) a |
|---|---|---|
| C. albicans SC5314 | Reference laboratory strain, FluS | 0 |
| C. albicans DSY347 | FluS clinical strain [27] | 0 |
| C. albicans DSY289 | FluR, DSY347 ERG11: S405F, Y132H; TAC1: A736V [27,29] | 14 |
| C. albicans DSY544 | FluS clinical strain [30] | 0 |
| C. albicans DSY775 | FluR, DSY544 ERG11: G464S, TAC1: G980W [30] | 0 |
| C. glabrata DSY562 | FluS clinical strain [28] | 13 |
| C. glabrata DSY565 | FluR clinical strain [28] | 12 |
| C. glabrata SFY93 | FluR, DSY562 pdr1∆ [31] | 12 |
| C. glabrata SFY105 | FluR, DSY562 pdr1∆-T588A [31] | 13 |
| C. glabrata SFY115 | FluR, DSY562 pdr1∆-L280F [31] | 13 |
| C. glabrata SFY116 | FluR, DSY562 pdr1∆-P822L [31] | 13 |
| C. lusitaniae NEQAS6208 | Reference laboratory strain, FluS | 17 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovati, L.; Santinoli, C.; Ferrari, E.; Ciociola, T.; Martin, E.; Bandi, C.; Ricci, I.; Epis, S.; Conti, S. Candidacidal Activity of a Novel Killer Toxin from Wickerhamomyces anomalus against Fluconazole-Susceptible and -Resistant Strains. Toxins 2018, 10, 68. https://doi.org/10.3390/toxins10020068
Giovati L, Santinoli C, Ferrari E, Ciociola T, Martin E, Bandi C, Ricci I, Epis S, Conti S. Candidacidal Activity of a Novel Killer Toxin from Wickerhamomyces anomalus against Fluconazole-Susceptible and -Resistant Strains. Toxins. 2018; 10(2):68. https://doi.org/10.3390/toxins10020068
Chicago/Turabian StyleGiovati, Laura, Claudia Santinoli, Elena Ferrari, Tecla Ciociola, Elena Martin, Claudio Bandi, Irene Ricci, Sara Epis, and Stefania Conti. 2018. "Candidacidal Activity of a Novel Killer Toxin from Wickerhamomyces anomalus against Fluconazole-Susceptible and -Resistant Strains" Toxins 10, no. 2: 68. https://doi.org/10.3390/toxins10020068
APA StyleGiovati, L., Santinoli, C., Ferrari, E., Ciociola, T., Martin, E., Bandi, C., Ricci, I., Epis, S., & Conti, S. (2018). Candidacidal Activity of a Novel Killer Toxin from Wickerhamomyces anomalus against Fluconazole-Susceptible and -Resistant Strains. Toxins, 10(2), 68. https://doi.org/10.3390/toxins10020068