Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review

Abstract
1. General Introduction: Aflatoxins, Aflatoxin B1, and Aflatoxicosis
2. Anti-AFB1 Dietary Agents
3. Selenium and Curcumin: General Introduction and Protective Mechanism
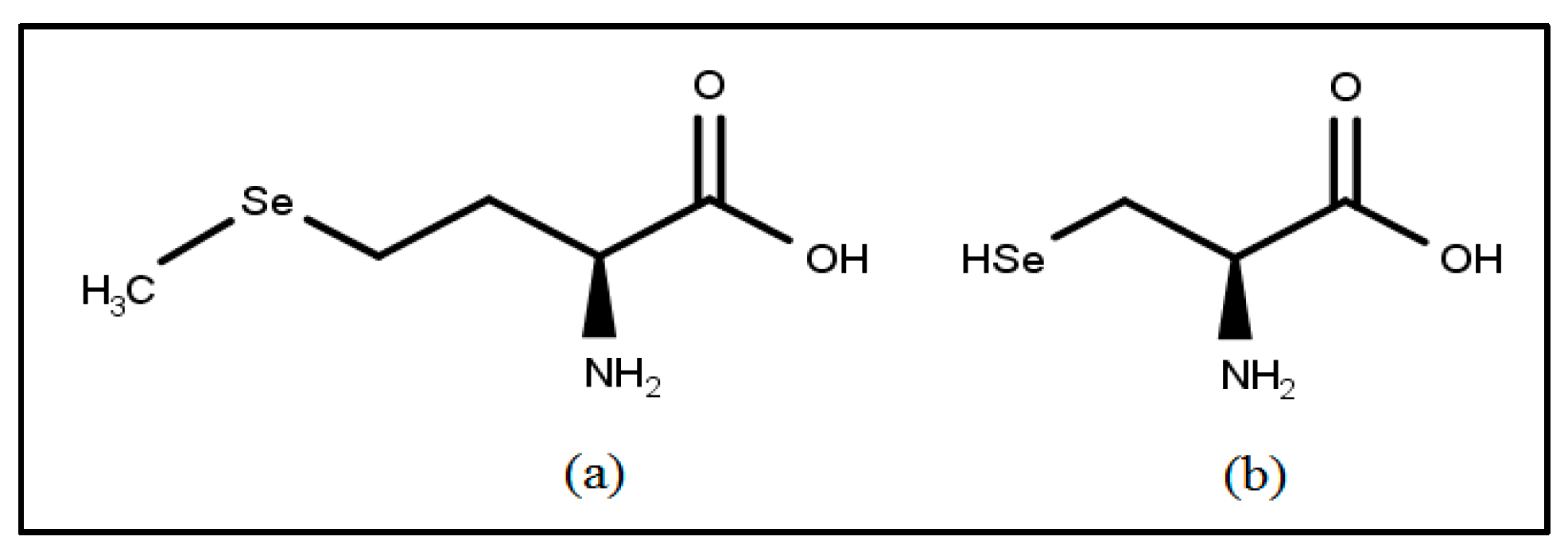
3.1. Selenium: Immunoprotective Antioxidant and Essential Trace Mineral
Selenium: General Mechanism of Action
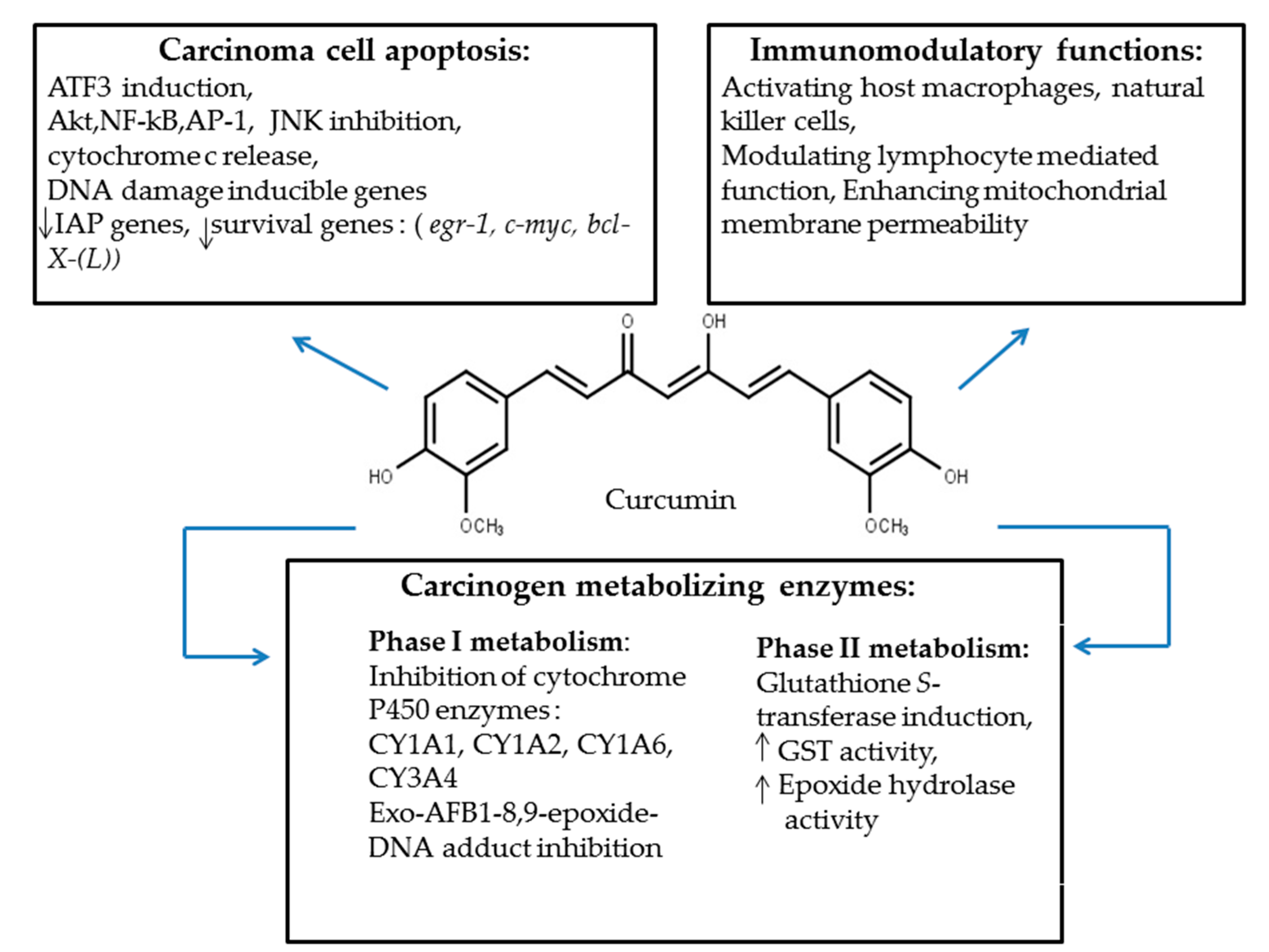
3.2. Curcumin: A Hepatoprotective Phytochemical
Curcumin: General Mechanism of Action
4. Selenium-Mediated AFB1 Toxicity Control and Protective Mechanisms
4.1. Selenium-Mediated Protection
4.1.1. Selenium-Mediated Anti-AFB1 Protection in Ducks
4.1.2. Selenium-Mediated Anti-AFB1 Protection in Poultry
4.1.3. Selenium-Mediated Anti-AFB1 Protection in Swine
5. Curcumin-Mediated AFB1 Toxicity Control and Protective Mechanisms
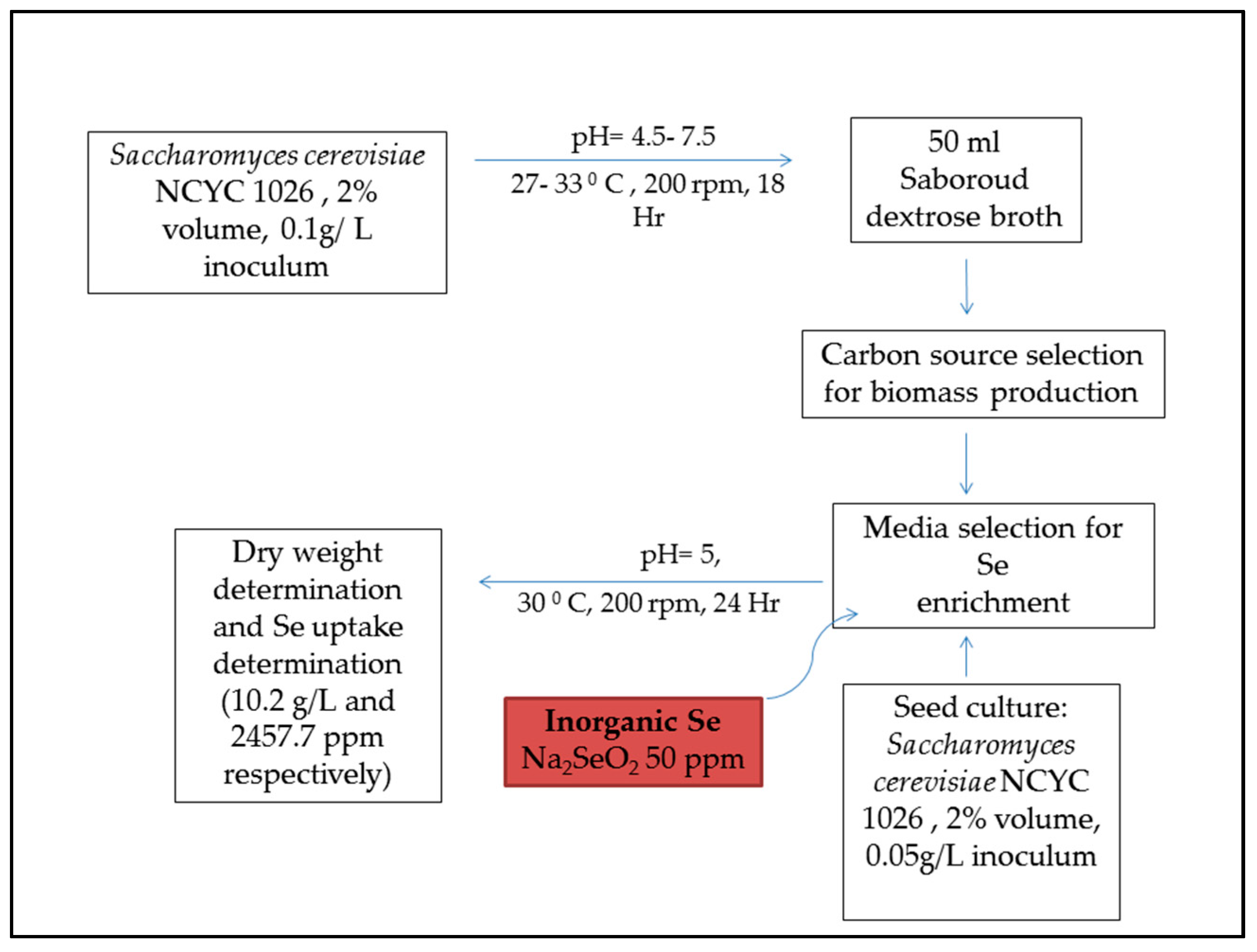
6. Selenium Supplementation of Animal Feeds
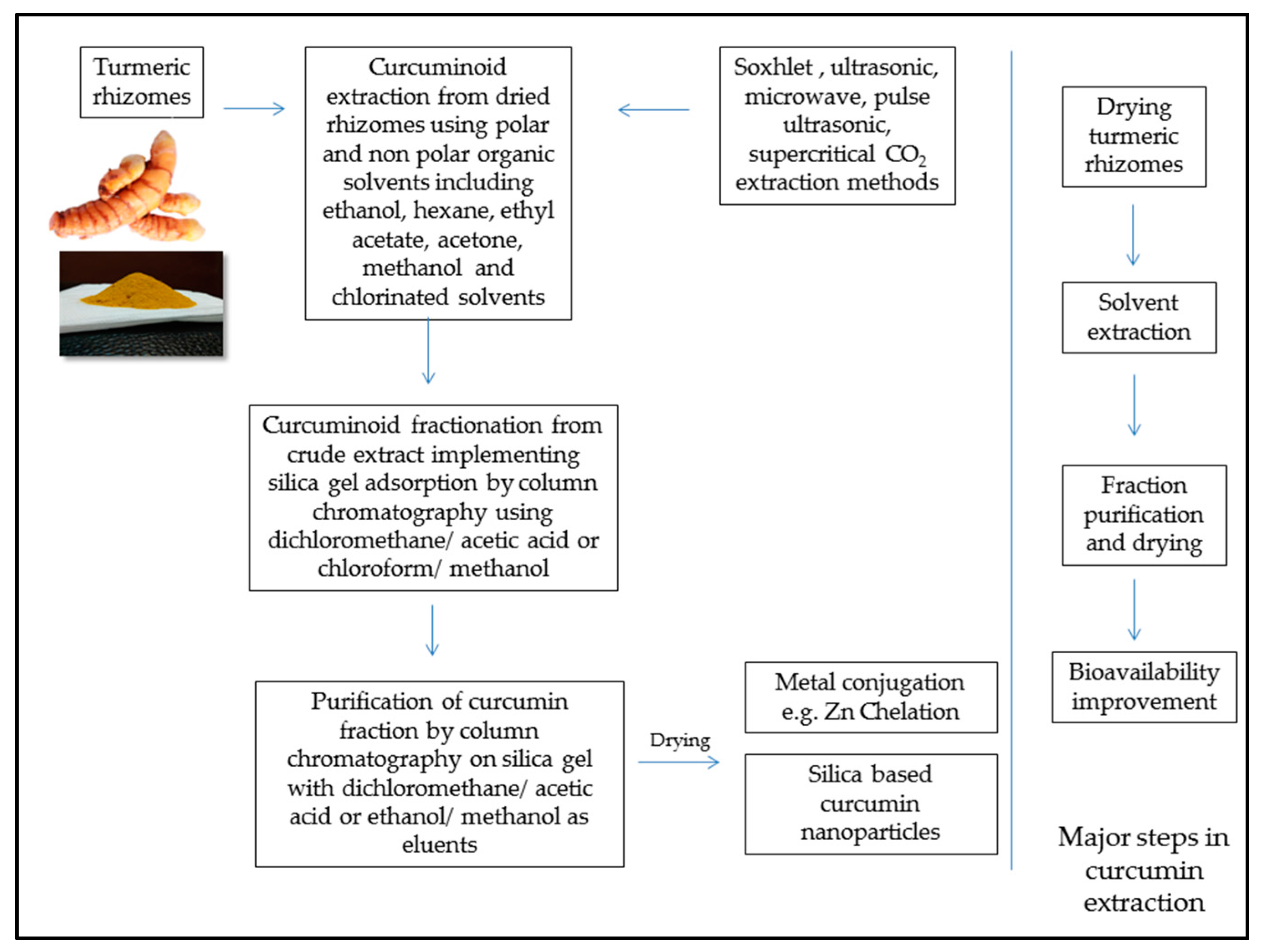
7. Curcumin Extraction and Feed Application
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin b(1) in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed]
- Mulunda, M.; Ngoma, L.; Nyirenda, M.; Motsei, L.; Bakunzi, F. A decade of aflatoxin M1 surveillance in milk and dairy products in developing countries (2001–2011): A review. In Mycotoxin and Food Safety in Developing Countries; Makun, D.H., Ed.; InTech: Rijeka, Croatia, 2013; pp. 39–60. ISBN 978-953-51-1096-5. [Google Scholar]
- Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer; World Health Organization: Lyon, France, 2002; pp. 1017–1606.
- Guchi, E. Aflatoxin contamination in groundnut arachis hypogaea caused by aspergillus species in ethiopia. J. Appl. Environ. Microbiol. 2015, 3, 11–19. [Google Scholar] [CrossRef]
- Mehdi, Y.; Dufrasne, I. Selenium in cattle: A review. Molecules 2016, 21, 545. [Google Scholar] [CrossRef] [PubMed]
- Astoreca, A.; Vaamonde, G.; Dalcero, A.; Marin, S.; Ramos, A. Abiotic factors and their interactions influence on the co-production of aflatoxin b1 and cyclopiazonic acid by Aspergillus flavus isolated from corn. Food Microbiol. 2014, 38, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-Y.; Roze, L.V.; Wee, J.; Linz, J.E. Evidence that a transcription factor regulatory network coordinates oxidative stress response and secondary metabolism in Aspergilli. MicrobiologyOpen 2013, 2, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Walte, H.-G.; Schwake-Anduschus, C.; Geisen, R.; Fritsche, J. Aflatoxin: Food chain transfer from feed to milk. J. Verbrauch. Lebensm. 2016, 11, 295–297. [Google Scholar] [CrossRef]
- Mace, K.; Aguilar, F.; Wang, J.S.; Vautravers, P.; Gomez-Lechon, M.; Gonzalez, F.J.; Groopman, J.; Harris, C.C.; Pfeifer, A.M. Aflatoxin b1-induced DNA adduct formation and p53 mutations in cyp450-expressing human liver cell lines. Carcinogenesis 1997, 18, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y.; Tang, L.; Wang, S.L.; Cai, Q.S.; Wang, J.S.; Hong, J.Y. Efficient activation of aflatoxin b1 by cytochrome p450 2a13, an enzyme predominantly expressed in human respiratory tract. Int. J. Cancer 2006, 118, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Dhanasekaran, D.; Shanmugapriya, S.; Thajuddin, N.; Panneerselvam, A. Aflatoxins and aflatoxicosis in human and animals. In Aflatoxins-Biochemistry and Molecular Biology; Ramon, G.G., Ed.; InTech: Rijeka, Croatia, 2011; pp. 221–254. ISBN 978-953-307-395-8. [Google Scholar]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in food and feed: Present status and future concerns. Compr. Rev. Food Sci. Food Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Dalvi, R.R. An overview of aflatoxicosis of poultry: Its characteristics, prevention and reduction. Vet. Res. Commun. 1986, 10, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Peng, X.; Fang, J.; Cui, H.; Yu, Z.; Chen, Z. Effects of aflatoxin b1 on T-cell subsets and mRNA expression of cytokines in the intestine of broilers. Int. J. Mol. Sci. 2015, 16, 6945–6959. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Bai, S.; Ding, X.; Zeng, Q.; Zhang, K.; Fang, J. Pathological changes in the immune organs of broiler chickens fed on corn naturally contaminated with aflatoxins b1 and b2. Avian Pathol. 2015, 44, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.A.; Kobashigawa, E.; Reis, T.A.; Mestieri, L.; Albuquerque, R.; Correa, B. Aflatoxin b1 residues in eggs of laying hens fed a diet containing different levels of the mycotoxin. Food Addit. Contam. 2000, 17, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Devreese, M.; De Backer, P.; Croubels, S. Overview of the most important mycotoxins for the pig and poultry husbandry. Vlaams Diergeneeskund. Tijdschr. 2013, 82, 171–180. [Google Scholar]
- Marin, D.E.; Taranu, I.; Bunaciu, R.P.; Pascale, F.; Tudor, D.S.; Avram, N.; Sarca, M.; Cureu, I.; Criste, R.D.; Suta, V.; et al. Changes in performance, blood parameters, humoral and cellular immune responses in weanling piglets exposed to low doses of aflatoxin. J. Anim. Sci. 2002, 80, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Rustemeyer, S.; Lamberson, W.; Ledoux, D.; Wells, K.; Austin, K.; Cammack, K. Effects of dietary aflatoxin on the hepatic expression of apoptosis genes in growing barrows. J. Anim. Sci. 2011, 89, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. Part A 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.; Bertuzzi, T.; Nielsen, K. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Applebaum, R.S.; Brackett, R.E.; Wiseman, D.W.; Marth, E.H. Responses of dairy cows to dietary aflatoxin: Feed intake and yield, toxin content, and quality of milk of cows treated with pure and impure aflatoxin. J. Dairy Sci. 1982, 65, 1503–1508. [Google Scholar] [CrossRef]
- Prandini, A.; Tansini, G.; Sigolo, S.; Filippi, L.; Laporta, M.; Piva, G. On the occurrence of aflatoxin M1 in milk and dairy products. Food Chem. Toxicol. 2009, 47, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Scheideler, S.E. Effects of various types of aluminosilicates and aflatoxin b1 on aflatoxin toxicity, chick performance, and mineral status. Poult. Sci. 1993, 72, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, C.E.; Cruz, I.B.; Cadona, F.C.; Machado, A.K.; Assmann, C.; Schlemmer, K.B.; Zanette, R.A.; Leal, D.B.; Santurio, J.M. Cytoprotective and genoprotective effects of beta-glucans against aflatoxin b(1)-induced DNA damage in broiler chicken lymphocytes. Toxicol. In Vitro 2015, 29, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Gurtoo, H.L.; Koser, P.L.; Bansal, S.K.; Fox, H.W.; Sharma, S.D.; Mulhern, A.I.; Pavelic, Z.P. Inhibition of aflatoxin b1-hepatocarcinogenesis in rats by β-naphthoflavone. Carcinogenesis 1985, 6, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, K.; Ho, L.L.; GopalanKriczky, P.; Lotlikar, P.D. A mechanism of inhibition of aflatoxin b1-hepatocarcinogenesis by β-naphthoflavone pretreatment of rats. Exp. Mol. Med. 1996, 28, 135–140. [Google Scholar] [CrossRef]
- Bintvihok, A.; Kositcharoenkul, S. Effect of dietary calcium propionate on performance, hepatic enzyme activities and aflatoxin residues in broilers fed a diet containing low levels of aflatoxin b1. Toxicon 2006, 47, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Stresser, D.M.; Bailey, G.S.; Williams, D.E. Indole-3-carbinol and β-naphthoflavone induction of aflatoxin b1 metabolism and cytochromes P450 associated with bioactivation and detoxication of aflatoxin b1 in the rat. Drug Metab. Dispos. 1994, 22, 383–391. [Google Scholar] [PubMed]
- Bammler, T.K.; Slone, D.H.; Eaton, D.L. Effects of dietary oltipraz and ethoxyquin on aflatoxin b1 biotransformation in non-human primates. Toxicol. Sci. 2000, 54, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, A.G.; Martinez, A.; Damian, F.J.; Quezada, T.; Ortiz, R.; Martinez, C.; Llamas, J.; Rodriguez, M.L.; Yamamoto, L.; Jaramillo, F.; et al. Efficacy of N-acetylcysteine to reduce the effects of aflatoxin b1 intoxication in broiler chickens. Poult. Sci. 2001, 80, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Lotlikar, P.D.; Raj, H.G.; Bohm, L.S.; Ho, L.L.; Jhee, E.C.; Tsuji, K.; Gopalan, P. A mechanism of inhibition of aflatoxin b1-DNA binding in the liver by phenobarbital pretreatment of rats. Cancer Res. 1989, 49, 951–957. [Google Scholar] [PubMed]
- Johnson, N.M.; Egner, P.A.; Baxter, V.K.; Sporn, M.B.; Wible, R.S.; Sutter, T.R.; Groopman, J.D.; Kensler, T.W.; Roebuck, B.D. Complete protection against aflatoxin b(1)-induced liver cancer with a triterpenoid: DNA adduct dosimetry, molecular signature, and genotoxicity threshold. Cancer Prev. Res. 2014, 7, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Alpsoy, L.; Yildirim, A.; Agar, G. The antioxidant effects of vitamin A, C, and E on aflatoxin b1-induced oxidative stress in human lymphocytes. Toxicol. Ind. Health 2009, 25, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Firozi, P.F.; Aboobaker, V.S.; Bhattacharya, R.K. Action of vitamin A on DNA adduct formation by aflatoxin b1 in a microsome catalyzed reaction. Cancer Lett. 1987, 34, 213–220. [Google Scholar] [CrossRef]
- Khan, W.A.; Khan, M.Z.; Khan, A.; Hassan, Z.U.; Rafique, S.; Saleemi, M.K.; Ahad, A. Dietary vitamin E in white leghorn layer breeder hens: A strategy to combat aflatoxin b1-induced damage. Avian Pathol. 2014, 43, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.; Salem, M.; Kamel, K.; Hassan, G.; El-Nouty, F. Influence of ascorbic acid supplementation on the haematological and clinical biochemistry parameters of male rabbits exposed to aflatoxin b1. J. Environ. Sci. Health Part B 2003, 38, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Netke, S.P.; Roomi, M.W.; Tsao, C.; Niedzwiecki, A. Ascorbic acid protects guinea pigs from acute aflatoxin toxicity. Toxicol. Appl. Pharmacol. 1997, 143, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lv, Y.; Huang, K.; Luo, Y.; Xu, W. Zinc inhibits aflatoxin b1-induced cytotoxicity and genotoxicity in human hepatocytes (HepG2 cells). Food Chem. Toxicol. 2016, 92, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, D.; mcCormick, S.P. The inhibitory effect of neem (Azadirachta indica) leaf extracts on aflatoxin synthesis in Aspergillus parasiticus. J. Am. Oil Chem. Soc. 1988, 65, 1166–1168. [Google Scholar] [CrossRef]
- Simonich, M.T.; Egner, P.A.; Roebuck, B.D.; Orner, G.A.; Jubert, C.; Pereira, C.; Groopman, J.D.; Kensler, T.W.; Dashwood, R.H.; Williams, D.E.; et al. Natural chlorophyll inhibits aflatoxin b1-induced multi-organ carcinogenesis in the rat. Carcinogenesis 2007, 28, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, V.; Hendricks, J.; Pereira, C.; Arbogast, D.; Bailey, G. Dietary chlorophyllin is a potent inhibitor of aflatoxin b1 hepatocarcinogenesis in rainbow trout. Cancer Res. 1995, 55, 57–62. [Google Scholar] [PubMed]
- Kelly, V.P.; Ellis, E.M.; Manson, M.M.; Chanas, S.A.; Moffat, G.J.; McLeod, R.; Judah, D.J.; Neal, G.E.; Hayes, J.D. Chemoprevention of aflatoxin b1 hepatocarcinogenesis by coumarin, a natural benzopyrone that is a potent inducer of aflatoxin b1-aldehyde reductase, the glutathione S-transferase A5 and P1 subunits, and NADPH:Quinone oxidoreductase in rat liver. Cancer Res. 2000, 60, 957–969. [Google Scholar] [PubMed]
- Ferk, F.; Huber, W.W.; Grasl-Kraupp, B.; Speer, K.; Buchmann, S.; Bohacek, R.; Mišík, M.; Edelbauer, L.; Knasmüller, S. Protective effects of coffee against induction of DNA damage and pre-neoplastic foci by aflatoxin b1. Mol. Nutr. Food Res. 2014, 58, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Esper, R.H.; Gonçalez, E.; Marques, M.O.M.; Felicio, R.C.; Felicio, J.D. Potential of essential oils for protection of grains contaminated by aflatoxin produced by Aspergillus flavus. Front. Microbiol. 2014, 5, 269. [Google Scholar] [CrossRef] [PubMed]
- Bergès, R.; Siess, M.-H.; Arnault, I.; Auger, J.; Kahane, R.; Pinnert, M.-F.; Vernevaut, M.-F.; le Bon, A.-M. Comparison of the chemopreventive efficacies of garlic powders with different alliin contents against aflatoxin b1 carcinogenicity in rats. Carcinogenesis 2004, 25, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Ahmed, H.H.; Hagazi, M.M. Prevention of aflatoxin b1-initiated hepatotoxicity in rat by marine algae extracts. J. Appl. Toxicol. 2006, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Rasouli-Hiq, A.A.; Bagherzadeh-Kasmani, F.; Mehri, M.; Karimi-Torshizi, M.A. Nigella sativa (black cumin seed) as a biological detoxifier in diet contaminated with aflatoxin b1. J. Anim. Physiol. Anim. Nutr. 2016, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Steidler, S.; Galletti, S.; Tameni, M.; Sonzogni, O.; Ravarotto, L. Efficacy of silymarin-phospholipid complex in reducing the toxicity of aflatoxin b1 in broiler chicks. Poult. Sci. 2004, 83, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Liu, F.; Zhu, Q.; Zhang, M.; Li, T.; Chen, J.; Huang, Y.; Wang, X.; Sheng, J. Aflatoxin b1 can be complexed with oxidised tea polyphenols and the absorption of the complexed aflatoxin b1 is inhibited in rats. J. Sci. Food Agric. 2016, 1910–1915. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Aly, S.E. Antioxidants and radical scavenging properties of vegetable extracts in rats fed aflatoxin-contaminated diet. J. Agric. Food Chem. 2003, 51, 2409–2414. [Google Scholar] [CrossRef] [PubMed]
- Hulikunte Mallikarjunaiah, N.; Jayapala, N.; Puttaswamy, H.; Siddapura Ramachandrappa, N. Characterization of non-aflatoxigenic strains of Aspergillus flavus as potential biocontrol agent for the management of aflatoxin contamination in groundnut. Microb. Pathog. 2017, 102, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.G.; Gao, X.; Zhou, T.; Zhao, L.H.; Fan, Y.; Li, X.Y.; Lei, Y.P.; Ji, C.; Zhang, J.Y. Protective effect of Bacillus subtilis ansb060 on egg quality, biochemical and histopathological changes in layers exposed to aflatoxin b1. Poult. Sci. 2012, 91, 2852–2857. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.; Wu, Q.K.; El-Nezami, H.; Juvonen, R.O.; Mykkanen, H.; Turner, P.C. Lactobacillus rhamnosus strain gg reduces aflatoxin b1 transport, metabolism, and toxicity in caco-2 cells. Appl. Environ. Microbiol. 2007, 73, 3958–3965. [Google Scholar] [CrossRef] [PubMed]
- Rather, I.A.; Seo, B.J.; Kumar, V.J.R.; Choi, U.-H.; Choi, K.-H.; Lim, J.; Park, Y.-H. Biopreservative potential of Lactobacillus plantarum yml007 and efficacy as a replacement for chemical preservatives in animal feed. Food Sci. Biotechnol. 2014, 23, 195–200. [Google Scholar] [CrossRef]
- Zaghini, A.; Martelli, G.; Roncada, P.; Simioli, M.; Rizzi, L. Mannanoligosaccharides and aflatoxin b1 in feed for laying hens: Effects on egg quality, aflatoxins b1 and M1 residues in eggs, and aflatoxin b1 levels in liver. Poult. Sci. 2005, 84, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Poloni, V.; Dogi, C.; Pereyra, C.M.; Fernandez Juri, M.G.; Kohler, P.; Rosa, C.A.; Dalcero, A.M.; Cavaglieri, L.R. Potentiation of the effect of a commercial animal feed additive mixed with different probiotic yeast strains on the adsorption of aflatoxin b1. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 970–976. [Google Scholar] [CrossRef] [PubMed]
- Zuo, R.-Y.; Chang, J.; Yin, Q.-Q.; Wang, P.; Yang, Y.-R.; Wang, X.; Wang, G.-Q.; Zheng, Q.-H. Effect of the combined probiotics with aflatoxin b1-degrading enzyme on aflatoxin detoxification, broiler production performance and hepatic enzyme gene expression. Food Chem. Toxicol. 2013, 59, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Çelik, K.; Denli, M.; Ertürk, M.; Öztürkcan, O.; Doran, F. Evaluation of dry yeast (Saccharomyces cerevisiae) compounds in the feed to reduce aflatoxin b1 (AFB1) residues and toxicity to japanese quails (Coturnix coturnix japonica). J. Appl. Anim. Res. 2001, 20, 245–250. [Google Scholar] [CrossRef]
- Magnoli, A.P.; Rodriguez, M.C.; González Pereyra, M.L.; Poloni, V.L.; Peralta, M.F.; Nilson, A.J.; Miazzo, R.D.; Bagnis, G.; Chiacchiera, S.M.; Cavaglieri, L.R. Use of yeast (Pichia kudriavzevii) as a novel feed additive to ameliorate the effects of aflatoxin b1 on broiler chicken performance. Mycotoxin Res. 2017, 33, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Monson, M.; Coulombe, R.; Reed, K. Aflatoxicosis: Lessons from toxicity and responses to aflatoxin b1 in poultry. Agriculture 2015, 5, 742–777. [Google Scholar] [CrossRef]
- Hua, G.; Woollins, J.D. Chapter 1 Organic phosphorus-selenium chemistry. In Selenium and Tellurium Chemistry: From Small Molecules to Biomolecules and Materials; Woollins, J.D., Laitinen, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–39. ISBN 978-3-642-20699-3. [Google Scholar]
- Perrone, D.; Monteiro, M.; Nunes, J.C. Chapter 1 The chemistry of selenium. In Selenium: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; The Royal Society of Chemistry: Cambridge, UK, 2015; pp. 3–15. ISBN 978-1-84973-891-0. [Google Scholar]
- Fredga, A. Organic selenium chemistry. Ann. N. Y. Acad. Sci. 1972, 192, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Reich, H.J.; Hondal, R.J. Why nature chose selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Fradejas, N.; Braun, D.; Seeher, S.; Schweizer, U. Chapter 2 Biochemistry and molecular biology of selenoproteins. In Selenium: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; The Royal Society of Chemistry: Cambridge, UK, 2015; pp. 16–39. ISBN 978-1-84973-891-0. [Google Scholar]
- Arthur, J.R.; McKenzie, R.C.; Beckett, G.J. Selenium in the immune system. J. Nutr. 2003, 133, 1457S–1459S. [Google Scholar] [PubMed]
- Köhrle, J.; Jakob, F.; Contempré, B.; Dumont, J.E. Selenium, the thyroid, and the endocrine system. Endocr. Rev. 2005, 26, 944–984. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Sies, H. Selenium homeostasis and antioxidant selenoproteins in brain: Implications for disorders in the central nervous system. Arch. Biochem. Biophys. 2013, 536, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Steward, W.P.; Gescher, A.J. Pharmacokinetics and pharmacodynamics of curcumin. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease; Aggarwal, B.B., Surh, Y.-J., Shishodia, S., Eds.; Springer: Boston, MA, USA, 2007; pp. 453–470. ISBN 978-0-387-46400-8. [Google Scholar]
- Priyadarsini, K. The chemistry of curcumin: From extraction to therapeutic agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Chun, K.-S.; Keum, Y.-S.; Han, S.S.; Song, Y.-S.; Kim, S.-H.; Surh, Y.-J. Curcumin inhibits phorbol ester-induced expression of cyclooxygenase-2 in mouse skin through suppression of extracellular signal-regulated kinase activity and nf-κb activation. Carcinogenesis 2003, 24, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Plummer, S.M.; Holloway, K.A.; Manson, M.M.; Munks, R.J.; Kaptein, A.; Farrow, S.; Howells, L. Inhibition of cyclo-oxygenase 2 expression in colon cells by the chemopreventive agent curcumin involves inhibition of nf-kappab activation via the nik/ikk signalling complex. Oncogene 1999, 18, 6013–6020. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zheng, S.; Lin, J.; Ryerse, J.; Chen, A. Curcumin protects the rat liver from ccl4 caused injury and fibrogenesis by attenuating oxidative stress and suppressing inflammation. Mol. Pharmacol. 2008, 73, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Musa, M.A.; Cooperwood, J.S.; Khan, M.O. A review of coumarin derivatives in pharmacotherapy of breast cancer. Curr. Med. Chem. 2008, 15, 2664–2679. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Gescher, A.J.; Steward, W.P. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A review of anti-cancer properties and therapeutic activity in head and neck squamous cell carcinoma. Mol. Cancer 2011, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.-Y.; Qi, M.; Zhao, L.; Zhu, M.-K.; Guo, J.; Liu, J.; Gu, C.-Q.; Rajput, S.; Krumm, C.; Qi, D.-S.; et al. Curcumin prevents aflatoxin b1 hepatoxicity by inhibition of cytochrome P450 isozymes in chick liver. Toxins 2016, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Brucato, M.; Sundlof, S.F.; Bell, J.U.; Edds, G.T. Aflatoxin b1 toxicosis in dairy calves pretreated with selenium-vitamin E. Am. J. Vet. Res. 1986, 47, 179–183. [Google Scholar] [PubMed]
- Chen, J.; Goetchius, M.P.; Combs, G.F., Jr.; Campbell, T.C. Effects of dietary selenium and vitamin E on covalent binding of aflatoxin to chick liver cell macromolecules. J. Nutr. 1982, 112, 350–355. [Google Scholar] [PubMed]
- Davila, J.C.; Edds, G.T.; Osuna, O.; Simpson, C.F. Modification of the effects of aflatoxin b1 and warfarin in young pigs given selenium. Am. J. Vet. Res. 1983, 44, 1877–1883. [Google Scholar] [PubMed]
- Guo, S.; Shi, D.; Liao, S.; Su, R.; Lin, Y.; Pan, J.; Tang, Z. Influence of selenium on body weights and immune organ indexes in ducklings intoxicated with aflatoxin b1. Biol. Trace Elem. Res. 2012, 146, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Zhang, N.Y.; Zhu, M.K.; Zhao, L.; Zhou, J.C.; Qi, D.S. Prevention of aflatoxin b1 hepatoxicity by dietary selenium is associated with inhibition of cytochrome P450 isozymes and up-regulation of 6 selenoprotein genes in chick liver. J. Nutr. 2016, 146, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.L.; Yang, Z.B.; Yang, W.R.; Jiang, S.Z.; Zhang, G.G.; Johnston, S.L.; Chi, F. Toxicity of increasing aflatoxin b1 concentrations from contaminated corn with or without clay adsorbent supplementation in ducklings. Poult. Sci. 2013, 92, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, K.Y.; Chen, D.W.; Ding, X.M.; Feng, G.D.; Ao, X. Effects of vitamin E and selenium yeast on growth performance and immune function in ducks fed maize naturally contaminated with aflatoxin b1. Livest. Sci. 2013, 152, 200–207. [Google Scholar] [CrossRef]
- Liao, S.; Shi, D.; Clemons-Chevis, C.L.; Guo, S.; Su, R.; Qiang, P.; Tang, Z. Protective role of selenium on aflatoxin b1-induced hepatic dysfunction and apoptosis of liver in ducklings. Biol. Trace Elem. Res. 2014, 162, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Guo, S.; Liao, S.; Su, R.; Pan, J.; Lin, Y.; Tang, Z. Influence of selenium on hepatic mitochondrial antioxidant capacity in ducklings intoxicated with aflatoxin b1. Biol. Trace Elem. Res. 2012, 145, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Liao, S.; Guo, S.; Li, H.; Yang, M.; Tang, Z. Protective effects of selenium on aflatoxin b1-induced mitochondrial permeability transition, DNA damage, and histological alterations in duckling liver. Biol. Trace Elem. Res. 2015, 163, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Fang, J.; Peng, X.; Cui, H.; Chen, J.; Wang, F.; Chen, Z.; Zuo, Z.; Deng, J.; Lai, W.; et al. Effect of selenium supplementation on aflatoxin b1-induced histopathological lesions and apoptosis in bursa of Fabricius in broilers. Food Chem. Toxicol. 2014, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, S.M.; Adachi, Y. Comparison of the effects of dietary selenium, zinc, and selenium and zinc supplementation on growth and immune response between chick groups that were inoculated with Salmonella and aflatoxin or Salmonella. Poult. Sci. 2000, 79, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Wang, F.; Peng, X.; Fang, J.; Cui, H.; Chen, Z.; Lai, W.; Zhou, Y.; Geng, Y. Effect of sodium selenite on pathological changes and renal functions in broilers fed a diet containing aflatoxin b1. Int. J. Environ. Res. Public Health 2015, 12, 11196–11208. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Sodhi, S.; Kaur, R. Effects of dietary supplements of selenium, vitamin E or combinations of the two on antibody responses of broilers. Br. Poult. Sci. 2006, 47, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shu, G.; Peng, X.; Fang, J.; Chen, K.; Cui, H.; Chen, Z.; Zuo, Z.; Deng, J.; Geng, Y.; et al. Protective effects of sodium selenite against aflatoxin b1-induced oxidative stress and apoptosis in broiler spleen. Int. J. Environ. Res. Public Health 2013, 10, 2834–2844. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fang, J.; Peng, X.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Lai, W.; Shu, G.; Tang, L. Effects of sodium selenite on aflatoxin b1-induced decrease of ileac T cell and the mRNA contents of IL-2, IL-6, and TNF-α in broilers. Biol. Trace Elem. Res. 2014, 159, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.S.; Coles, B.F.; Raney, K.D.; Thier, R.; Guengerich, F.P.; Harris, T.M. DNA adduction by the potent carcinogen aflatoxin b1: Mechanistic studies. J. Am. Chem. Soc. 1994, 116, 1603–1609. [Google Scholar] [CrossRef]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin b1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Che, C.; Korolchuk, V.I.; Gan, F.; Pan, C.; Huang, K. Selenomethionine alleviates AFB1-induced damage in primary chicken hepatocytes by inhibiting CYP450 1A5 expression via upregulated selw expression. J. Agric. Food Chem. 2017, 65, 2495–2502. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Hu, J.; Song, S.; Huang, D.; Xu, H.; Qian, G.; Gan, F.; Huang, K. Selenium alleviates aflatoxin b1-induced immune toxicity through improving glutathione peroxidase 1 and selenoprotein S expression in primary porcine splenocytes. J. Agric. Food Chem. 2016, 64, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Innao, V.; Russo, S.; Gerace, D.; Alonci, A.; Musolino, C. Anticancer activity of curcumin and its analogues: Preclinical and clinical studies. Cancer Investig. 2016, 35, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bisht, S.; Feldmann, G.; Soni, S.; Ravi, R.; Karikar, C.; Maitra, A.; Maitra, A. Polymeric nanoparticle-encapsulated curcumin (“nanocurcumin”): A novel strategy for human cancer therapy. J. Nanobiotechnol. 2007, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.E.; Kuo, M.L.; Hsu, C.H.; Chen, C.R.; Lin, J.K.; Lai, G.M.; Hsieh, C.Y.; Cheng, A.L. Curcumin-containing diet inhibits diethylnitrosamine-induced murine hepatocarcinogenesis. Carcinogenesis 2000, 21, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Dasiram, J.D.; Ganesan, R.; Kannan, J.; Kotteeswaran, V.; Sivalingam, N. Curcumin inhibits growth potential by g1 cell cycle arrest and induces apoptosis in p53-mutated COLO 320DM human colon adenocarcinoma cells. Biomed. Pharmacother. 2016, 86, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, M.; Piwoni, A.; Filiczak, N.; Janicka, M.; Gubernator, J. Long-circulating curcumin-loaded liposome formulations with high incorporation efficiency, stability and anticancer activity towards pancreatic adenocarcinoma cell lines in vitro. PLoS ONE 2016, 11, e0167787. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, D.S. Comparative effects of curcumin and resveratrol on aflatoxin b1-induced liver injury in rats. Arch. Toxicol. 2010, 84, 389–396. [Google Scholar] [CrossRef] [PubMed]
- El-Mahalaway, A.M. Protective effect of curcumin against experimentally induced aflatoxicosis on the renal cortex of adult male albino rats: A histological and immunohisochemical study. Int. J. Clin. Exp. Pathol. 2015, 8, 6019–6030. [Google Scholar] [PubMed]
- Firozi, P.F.; Aboobaker, V.S.; Bhattacharya, R.K. Action of curcumin on the cytochrome P450-system catalyzing the activation of aflatoxin b1. Chem. Biol. Interact. 1996, 100, 41–51. [Google Scholar] [CrossRef]
- Gowda, N.K.S.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Efficacy of turmeric (Curcuma longa), containing a known level of curcumin, and a hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of aflatoxin in broiler chicks. Poult. Sci. 2008, 87, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.B.; Lahiri, M.; Chackradeo, P.; Bhide, S.V.; Kuttan, R. Protective effect of food additives on aflatoxin-induced mutagenicity and hepatocarcinogenicity. Cancer Lett. 1997, 115, 129–133. [Google Scholar] [CrossRef]
- Soni, K.B.; Rajan, A.; Kuttan, R. Reversal of aflatoxin induced liver damage by turmeric and curcumin. Cancer Lett. 1992, 66, 115–121. [Google Scholar] [CrossRef]
- Verma, R.J.; Chakraborty, B.S.; Patel, C.; Mathuria, N. Curcumin ameliorates aflatoxin-induced changes in SDH and ATPase activities in liver and kidney of mice. Acta Pol. Pharm. 2008, 65, 415–419. [Google Scholar] [PubMed]
- Verma, R.J.; Mathuria, N. Curcumin ameliorates aflatoxin-induced lipid-peroxidation in liver and kidney of mice. Acta Pol. Pharm. 2008, 65, 195–202. [Google Scholar] [PubMed]
- El-Bahr, S.M. Effect of curcumin on hepatic antioxidant enzymes activities and gene expressions in rats intoxicated with aflatoxin b1. Phytother. Res. 2015, 29, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-E.; Campbell, B.C.; Molyneux, R.J.; Hasegawa, S.; Lee, H.-S. Inhibitory effects of naturally occurring compounds on aflatoxin b1 biotransformation. J. Agric. Food Chem. 2001, 49, 5171–5177. [Google Scholar] [CrossRef] [PubMed]
- Gowda, N.K.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Antioxidant efficacy of curcuminoids from turmeric (Curcuma longa L.) powder in broiler chickens fed diets containing aflatoxin b1. Br. J. Nutr. 2009, 102, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Sun, X.; Wang, H.; Li, W.; Wang, X.; Cheng, P.; Li, S.; Zhang, X.; Hamid, S. Curcumin successfully inhibited the computationally identified CYP2A6 enzyme-mediated bioactivation of aflatoxin b1 in arbor acres broiler. Front. Pharmacol. 2017, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Schrauzer, G.N. The nutritional significance, metabolism and toxicology of selenomethionine. Adv. Food Nutr. Res. 2003, 47, 73–112. [Google Scholar] [PubMed]
- Kubachka, K.M.; Hanley, T.; Mantha, M.; Wilson, R.A.; Falconer, T.M.; Kassa, Z.; Oliveira, A.; Landero, J.; Caruso, J. Evaluation of selenium in dietary supplements using elemental speciation. Food Chem. 2017, 218, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Fisinin, V.I. Selenium in poultry breeder nutrition: An update. Anim. Feed Sci. Technol. 2014, 191, 1–15. [Google Scholar] [CrossRef]
- Surai, P.; Dvorska, J. Is organic selenium better for animals than inorganic sources? Feed Mix 2001, 9, 8–10. [Google Scholar]
- Papazyan, T.; Denev, S.; Surai, P. Selenium in poultry nutrition: Lessons from research and wild nature. Krmiva 2006, 48, 275–283. [Google Scholar]
- Koller, L.D.; Exon, J.H. The two faces of selenium-deficiency and toxicity—Are similar in animals and man. Can. J. Vet. Res. 1986, 50, 297–306. [Google Scholar] [PubMed]
- Tiwary, A.K.; Stegelmeier, B.L.; Panter, K.E.; James, L.F.; Hall, J.O. Comparative toxicosis of sodium selenite and selenomethionine in lambs. J. Vet. Diagn. Investig. 2006, 18, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Barger, J.L.; Kayo, T.; Pugh, T.D.; Vann, J.A.; Power, R.; Dawson, K.; Weindruch, R.; Prolla, T.A. Gene expression profiling reveals differential effects of sodium selenite, selenomethionine, and yeast-derived selenium in the mouse. Genes Nutr. 2012, 7, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Guo, F.; Zhang, L.; Dong, B.; Gong, L. Effects of dietary selenomethionine supplementation on growth performance, antioxidant status, plasma selenium concentration, and immune function in weaning pigs. J. Anim. Sci. Biotechnol. 2014, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.L.; Lavergne, T.K.; Southern, L.L. Effect of inorganic versus organic selenium on hen production and egg selenium concentration. Poult. Sci. 2005, 84, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Tait, S.J.; Collings, R.; Hurst, R. Selenium bioavailability: Current knowledge and future research requirements. Am. J. Clin. Nutr. 2010, 91, 1484S–1491S. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Fisinin, V.I. Selenium in sow nutrition. Anim. Feed Sci. Technol. 2016, 211, 18–30. [Google Scholar] [CrossRef]
- Kieliszek, M.; Blazejak, S.; Gientka, I.; Bzducha-Wrobel, A. Accumulation and metabolism of selenium by yeast cells. Appl. Microbiol. Biotechnol. 2015, 99, 5373–5382. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, S.; Khosravi-Darani, K.; Pourahmad, R.; Komeili, R. An experimental design for production of selenium-enriched yeast. World Appl. Sci. 2012, 19, 31–37. [Google Scholar] [CrossRef]
- Rajashree, K. Preparation of selenium tolerant yeast Saccharomyces cerevisiae. J. Microbiol. Biotechnol. Res. 2017, 3, 46–53. [Google Scholar]
- Rajashree, K.; Muthukumar, T. Selection of culture medium and conditions for the production of selenium enriched Saccharomyces cerevisiae. Afr. J. Biotechnol. 2013, 12, 2972–2977. [Google Scholar]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: How does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Sareen, R.; Jain, N.; Dhar, K.L. Curcumin-Zn(II) complex for enhanced solubility and stability: An approach for improved delivery and pharmacodynamic effects. Pharm. Dev. Technol. 2016, 21, 630–635. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agent Type | Dietary Agent | Reference |
|---|---|---|
| Chemical | Aluminosilicates | [27] |
| β-1,3 glucan | [28] | |
| β-napthoflavone | [29] | |
| Butylated hydroxianisole | [30] | |
| Calcium propionate | [31] | |
| Indole-3-carbinol | [32] | |
| Oltipraz & Ethoxyquin | [33] | |
| N-acetylcysteine | [34] | |
| Phenobarbital | [35] | |
| Triterpenoid | [36] | |
| Vitamins A, C and E | [37,38,39,40,41] | |
| Zinc | [42] | |
| Biological | Azadirachta indica extract | [43] |
| (Plant origin) | Chlorophyll | [44] |
| Chlorophyllin | [45] | |
| Coumarin | [46] | |
| Coffee | [47] | |
| Essential oils | [48] | |
| Garlic powders | [49] | |
| Marine algae extract | [50] | |
| Nigella sativa | [51] | |
| Silymarin | [52] | |
| Tea polyphenols | [53] | |
| Vegetable extracts | [54] | |
| Biological | Atoxigenic strains of Aspergillus flavus | [55] |
| (Microbial origin) | Bacillus and Lactobacillus spp. probiotics | [56,57,58] |
| Mannonoligosaccherides | [59] | |
| Probiotics mixed with AFB1 degrading enzyme | [60,61] | |
| Yeast species | [62,63] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limaye, A.; Yu, R.-C.; Chou, C.-C.; Liu, J.-R.; Cheng, K.-C. Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review. Toxins 2018, 10, 25. https://doi.org/10.3390/toxins10010025
Limaye A, Yu R-C, Chou C-C, Liu J-R, Cheng K-C. Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review. Toxins. 2018; 10(1):25. https://doi.org/10.3390/toxins10010025
Chicago/Turabian StyleLimaye, Aniket, Roch-Chui Yu, Cheng-Chun Chou, Je-Ruei Liu, and Kuan-Chen Cheng. 2018. "Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review" Toxins 10, no. 1: 25. https://doi.org/10.3390/toxins10010025
APA StyleLimaye, A., Yu, R.-C., Chou, C.-C., Liu, J.-R., & Cheng, K.-C. (2018). Protective and Detoxifying Effects Conferred by Dietary Selenium and Curcumin against AFB1-Mediated Toxicity in Livestock: A Review. Toxins, 10(1), 25. https://doi.org/10.3390/toxins10010025