Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Diets
2.2. Tissue Collection and Processing and Fetal Anthropometry
2.3. Measurements of Hepatic and Placenta Metabolites
2.4. Quantitative Real-Time RT-PCR
2.5. Statistical Analysis
3. Results
3.1. Effects of Maternal Choline Supplementation
3.1.1. Pregnancy Outcomes
3.1.2. Fetal and Placental Growth
3.1.3. Hepatic and Placental Choline Metabolites
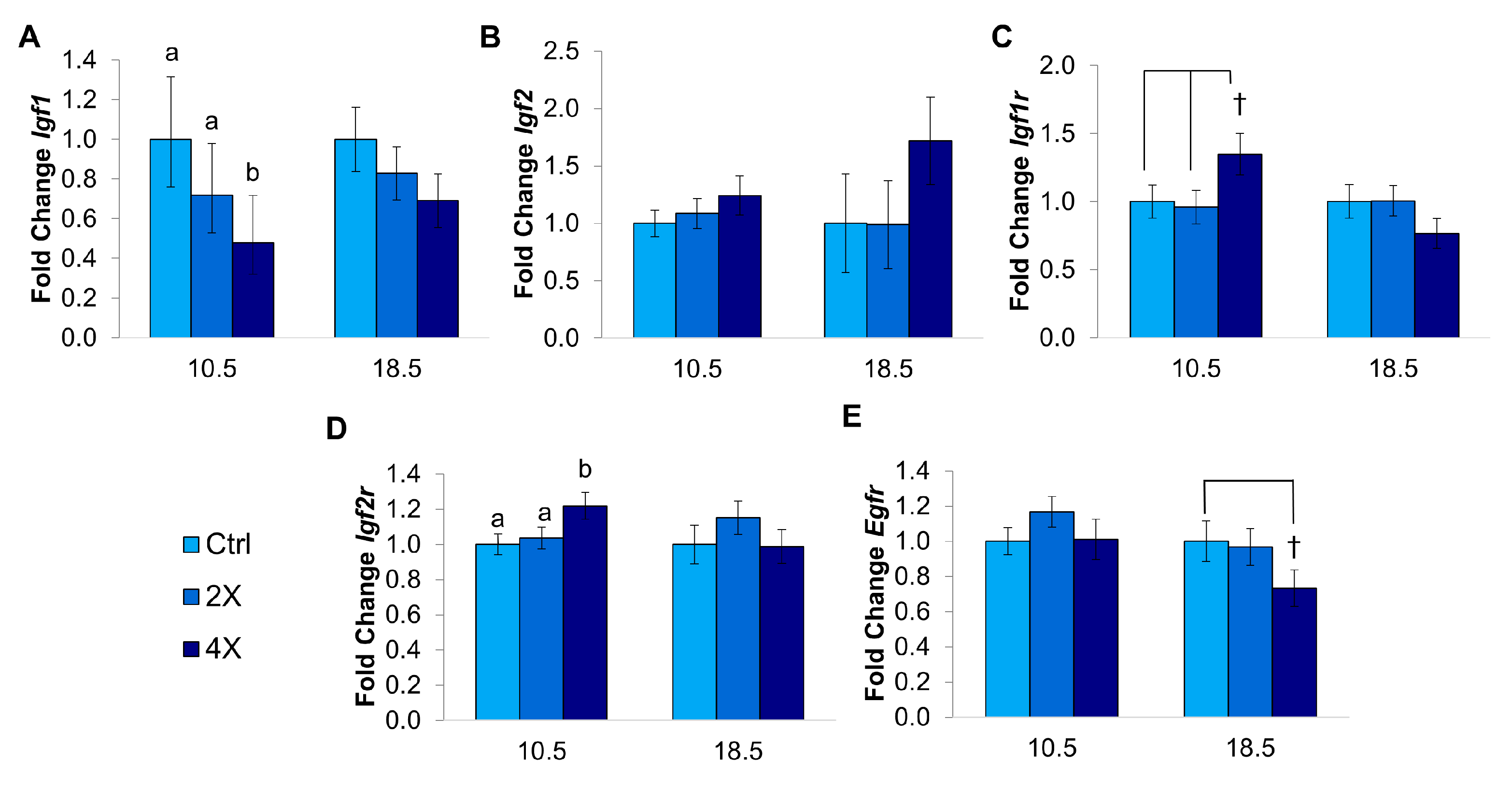
3.1.4. Placental Growth Factor Gene Expression
3.2. Effects of Dlx3 Genotype
3.2.1. Fetal and Placental Growth
3.2.2. Growth Factor Gene Expression in the Placenta
3.3. Determinants of Fetal Growth
3.3.1. Choline Metabolites
3.3.2. Expression of Growth Factor Genes
4. Discussion
4.1. Supplementing the Maternal Diet with Extra Choline Accelerates Fetal Growth during the First Half of Pregnancy
4.2. This Acceleration in Fetal Growth during Early Pregnancy Does Not Result in Overgrowth
4.3. Supplementing the Maternal Diet with Extra Choline May Uniquely Benefit Pregnancies Characterized by Placental Insufficiency
4.4. The Main Metabolic Fate of the Supplemental Choline in Our Study Was Betaine
4.5. Maternal Choline Supplementation also Altered Hepatic Phosphocholine and GPC Levels
4.6. Enhanced Early Growth Led to Placental down Regulation of Igf1
4.7. Dlx3+/− Placentas Had Higher Expression of Growth Factor Genes
4.8. Strengths and Limitations
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fowden, A.L.; Sferruzzi-Perri, A.N.; Coan, P.M.; Constancia, M.; Burton, G.J. Placental efficiency and adaptation: Endocrine regulation. J. Physiol. 2009, 587, 3459–3472. [Google Scholar] [CrossRef] [PubMed]
- Duley, L. Pre-eclampsia and the hypertensive disorders of pregnancy. Br. Med. Bull. 2003, 67, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Salam, R.A.; Das, J.K.; Bhutta, Z.A. Impact of intrauterine growth restriction on long-term health. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Shatenstein, B.; Luo, Z.-C.; Wei, S.; Fraser, W. Role of nutrition in the risk of preeclampsia. Nutr. Rev. 2009, 67, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Vance, D.E. Phosphatidylcholine and choline homeostasis. J. Lipid Res. 2008, 49, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.A. Pre- and Postnatal Health: Evidence of Increased Choline Needs. J. Am. Diet. Assoc. 2010, 110, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Bar, H.Y.; Yan, J.; Jones, S.; Brannon, P.M.; West, A.A.; Perry, C.A.; Ganti, A.; Pressman, E.; Devapatla, S.; et al. A higher maternal choline intake among third-trimester pregnant women lowers placental and circulating concentrations of the antiangiogenic factor fms-like tyrosine kinase-1 (sFLT1). FASEB J. 2013, 27, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Jones, S.; Andrew, B.Y.; Ganti, A.; Malysheva, O.V.; Giallourou, N.; Brannon, P.M.; Roberson, M.S.; Caudill, M.A. Choline inadequacy impairs trophoblast function and vascularization in cultured human placental trophoblasts. J. Cell. Physiol. 2014, 229, 1016–1027. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.T.; King, J.H.; Yan, J.; Jiang, X.; Wei, E.; Fomin, V.G.; Roberson, M.S.; Caudill, M.A. Maternal choline supplementation during murine pregnancy modulates placental markers of inflammation, apoptosis and vascularization in a fetal sex-dependent manner. Placenta 2017, 53, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Morasso, M.I.; Grinberg, A.; Robinson, G.; Sargent, T.D.; Mahon, K.A. Placental failure in mice lacking the homeobox gene Dlx3. Proc. Natl. Acad. Sci. USA 1999, 96, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Clark, P.A.; Brown, J.L.; Li, S.; Woods, A.K.; Han, L.; Sones, J.L.; Preston, R.L.; Southard, T.L.; Davisson, R.L.; Roberson, M.S. Distal-less 3 haploinsufficiency results in elevated placental oxidative stress and altered fetal growth kinetics in the mouse. Placenta 2012, 33, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Bowman, C.J.; Streck, R.D.; Chapin, R.E. Maternal-placental insulin-like growth factor (IGF) signaling and its importance to normal embryo-fetal development. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.; Westwood, M. The IGF axis and placental function. a mini review. Horm. Res. 2008, 69, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Forbes, K.; Westwood, M. Maternal growth factor regulation of human placental development and fetal growth. J. Endocrinol. 2010, 207, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Van der Veeken, J.; Oliveira, S.; Schiffelers, R.M.; Storm, G.; van Bergen En Henegouwen, P.M.P.; Roovers, R.C. Crosstalk between epidermal growth factor receptor- and insulin-like growth factor-1 receptor signaling: Implications for cancer therapy. Curr. Cancer Drug Targets 2009, 9, 748–760. [Google Scholar] [CrossRef] [PubMed]
- Napoli, I.; Blusztajn, J.K.; Mellott, T.J. Prenatal choline supplementation in rats increases the expression of IGF2 and its receptor IGF2R and enhances IGF2-induced acetylcholine release in hippocampus and frontal cortex. Brain Res. 2008, 1237, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Kovacheva, V.P.; Mellott, T.J.; Davison, J.M.; Wagner, N.; Lopez-Coviella, I.; Schnitzler, A.C.; Blusztajn, J.K. Gestational choline deficiency causes global and Igf2 gene DNA hypermethylation by up-regulation of Dnmt1 expression. J. Biol. Chem. 2007, 282, 31777–31788. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Surzenko, N.; Friday, W.B.; Zeisel, S.H. Maternal dietary intake of choline in mice regulates development of the cerebral cortex in the offspring. FASEB J. 2016, 30, 1566–1578. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Jiang, X.; West, A.A.; Perry, C. Maternal choline intake modulates maternal and fetal biomarkers of choline metabolism in humans. Am. J. 2012, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Leshner, A.I.; Litwin, V.A.; Squibb, R.L. A simple method for carcass analysis. Physiol. Behav. 1972, 9, 281–282. [Google Scholar] [CrossRef]
- Koc, H.; Mar, M.H.; Ranasinghe, A.; Swenberg, J.A.; Zeisel, S.H. Quantitation of choline and its metabolites in tissues and foods by liquid chromatography/electrospray ionization-isotope dilution mass spectrometry. Anal. Chem. 2002, 74, 4734–4740. [Google Scholar] [CrossRef] [PubMed]
- Holm, P.I.; Ueland, P.M.; Kvalheim, G.; Lien, E.A. Determination of choline, betaine, and dimethylglycine in plasma by a high-throughput method based on normal-phase chromatography-tandem mass spectrometry. Clin. Chem. 2003, 49, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Mehedint, M.G.; Craciunescu, C.N.; Zeisel, S.H. Maternal dietary choline deficiency alters angiogenesis in fetal mouse hippocampus. Proc. Natl. Acad. Sci. USA 2010, 107, 12834–12839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Denomme, M.M.; White, C.R.; Leung, K.-Y.; Lee, M.B.; Greene, N.D.E.; Mann, M.R.W.; Trasler, J.M.; Baltz, J.M. Both the folate cycle and betaine-homocysteine methyltransferase contribute methyl groups for DNA methylation in mouse blastocysts. FASEB J. 2015, 29, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Kalari, K.; Fridley, B.L.; Jenkins, G.; Ji, Y.; Abo, R.; Hebbring, S.; Zhang, J.; Nye, M.D.; Leeder, J.S.; et al. Betaine-homocysteine methyltransferase: Human liver genotype-phenotype correlation. Mol. Genet. Metab. 2011, 102, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Van Lee, L.; Tint, M.T.; Aris, I.M.; Quah, P.L.; Fortier, M.V.; Lee, Y.S.; Yap, F.K.; Saw, S.M.; Godfrey, K.M.; Gluckman, P.D.; et al. Prospective associations of maternal betaine status with offspring weight and body composition at birth: The Growing Up in Singapore Towards healthy Outcomes (GUSTO) cohort study. Am. J. Clin. Nutr. 2016, 104, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Hogeveen, M.; den Heijer, M.; Semmekrot, B.A.; Sporken, J.M.; Ueland, P.M.; Blom, H.J. Umbilical choline and related methylamines betaine and dimethylglycine in relation to birth weight. Pediatr. Res. 2013, 73, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Chew, T.W.; Jiang, X.; Yan, J.; Wang, W.; Lusa, A.L.; Carrier, B.J.; West, A.A.; Malysheva, O.V.; Brenna, J.T.; Iii, J.F.G.; et al. Folate Intake, Mthfr Genotype, and Sex Modulate Choline Metabolism in Mice. Online 2011, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Austdal, M.; Thomsen, L.C.V.; Tangerås, L.H.; Skei, B.; Mathew, S.; Bjørge, L.; Austgulen, R.; Bathen, T.F.; Iversen, A.-C. Metabolic profiles of placenta in preeclampsia using HR-MAS MRS metabolomics. Placenta 2015, 36, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Jones, H.N.; Crombleholme, T.; Habli, M. Adenoviral-Mediated Placental Gene Transfer of IGF-1 Corrects Placental Insufficiency via Enhanced Placental Glucose Transport Mechanisms. PLoS ONE 2013, 8, e74632. [Google Scholar] [CrossRef] [PubMed]
- Dackor, J.; Caron, K.M.; Threadgill, D.W. Placental and Embryonic Growth Restriction in Mice With Reduced Function Epidermal Growth Factor Receptor Alleles. Genetics 2009, 183, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, K.; Xu, X.; Liu, Z.; Lin, C.; Wang, Y.; Yan, H.; Yang, X. [Correlation of insulin-like growth factor 1 expression in placenta with DNA methylation and fetal macrosomia]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2015, 32, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.; Kar, S. The insulin-like growth factor-II/mannose-6-phosphate receptor: Structure, distribution and function in the central nervous system. Brain Res. Brain Res. Rev. 2004, 44, 117–140. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.L.; Hassan, A.B. Cell survival and proliferation are modified by insulin-like growth factor 2 between days 9 and 10 of mouse gestation. Development 2001, 128, 3819–3830. [Google Scholar] [PubMed]
- Kaffer, C.R.; Grinberg, A.; Pfeifer, K. Regulatory mechanisms at the mouse Igf2/H19 locus. Mol. Cell. Biol. 2001, 21, 8189–8196. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sheikh, S.; Satoskar, P.; Bhartiya, D. Expression of insulin-like growth factor-I and placental growth hormone mRNA in placentae: A comparison between normal and intrauterine growth retardation pregnancies. Mol. Hum. Reprod. 2001, 7, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez, G.; González, C.A.; Argandoña, F.; Kakarieka, E.; Johnson, M.C.; Cassorla, F. Expression and protein content of IGF-I and IGF-I receptor in placentas from small, adequate and large for gestational age newborns. Horm. Res. Paediatr. 2010, 73, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Swanson, A.M.; David, A.L. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta 2015, 36, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet (Lond. UK) 2008, 371, 75–84. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Time Point | Diet | Choline (nmol/g) | Methionine (nmol/g) | GPC (nmol/g) | LPC (nmol/g) | Betaine (µmol/g) | PCho (µmol/g) | PC (µmol/g) | SM (µmol/g) |
|---|---|---|---|---|---|---|---|---|---|
| 10.5 | 1X | 214 (173, 265) | 124 (95, 161) | 948 ± 105 | 332 (269, 411) | 0.81 ± 0.1 | 1.03 ± 0.2 | 22.2 ± 0.9 | 3.94 ± 0.2 |
| 2X | 242 (186, 313) | 92.7 (67, 127) | 1021 ± 126 | 326 (252, 421) | 1.49 ± 0.2 * | 1.41 ± 0.2 | 21.6 ± 1.1 | 3.70 ± 0.2 | |
| 4X | 247 (197, 310) | 99.1 (75, 131) | 895 ± 110 | 347 (277, 435) | 1.71 ± 0.1 * | 1.07 ± 0.2 | 22.2 ± 0.9 | 4.16 ± 0.2 | |
| 12.5 | 1X | 206 (173, 245) | 85.5 (70, 104) | 865 ± 54 | 363 (302, 437) | 0.99 ± 0.1 | 1.08 (0.7, 1.6) | 22.6 ± 0.7 | 3.97 ± 0.2 |
| 2X | 188 (156, 226) | 108 (88, 133) | 687 ± 57 * | 353 (292, 429) | 1.77 ± 0.2 * | 1.20 (0.8, 1.8) | 22.5 ± 0.7 | 4.03 ± 0.2 | |
| 4X | 197 (166, 233) | 97.5 (81, 118) | 687 ± 52 * | 371 (308, 446) | 2.13 ± 0.1 * | 1.38 (1.0, 2.0) | 22.5 ± 0.7 | 3.75 ± 0.2 | |
| 15.5 | 1X | 138 (116, 163) | 88.7 ± 5.9 | 294 (225, 382) | 339 (259, 443) | 1.21 ± 0.2 | 0.70 ± 0.2 | 22.4 ± 0.8 | 3.44 ± 0.2 |
| 2X | 133 (111, 160) | 88.7 ± 6.4 | 337 (252, 449) | 322 (245, 424) | 1.60 ± 0.2 * | 0.73 ± 0.3 | 21.6 ± 0.8 | 3.43 ± 0.2 | |
| 4X | 144 (121, 171) | 75.6 ± 6.0 | 262 (200, 344) | 318 (243, 414) | 2.32 ± 0.2 * | 1.63 ± 0.3 * | 22.3 ± 0.8 | 3.20 ± 0.2 | |
| 18.5 | 1X | 117 (97, 141) | 104 ± 7.1 | 301 ± 34 | 286 (231, 355) | 0.53 ± 0.2 | 0.58 ± 0.2 | 22.3 ± 1.0 | 3.10 ± 0.1 |
| 2X | 141 (116, 170) | 83.3 ± 7.2 | 302 ± 34 | 262 (211, 325) | 1.34 ± 0.2 * | 1.04 ± 0.2 | 21.3 ± 1.0 | 3.23 ± 0.1 | |
| 4X | 137 (113, 168) | 97.4 ± 7.6 | 257 ± 36 | 252 (200, 316) | 2.35 ± 0.3 ** | 0.91 ± 0.2 | 20.8 ±1.0 | 2.94 ± 0.1 |
| Time Point | Diet | Choline | Methionine | Betaine |
|---|---|---|---|---|
| E10.5 | 1X | 0.50 ± 0.04 | 0.27 ± 0.02 | 3.09 ± 0.24 |
| 2X | 0.56 ± 0.04 | 0.30 ± 0.02 | 3.31 ± 0.27 * | |
| 4X | 0.52 ± 0.05 | 0.29 ± 0.03 | 4.28 ± 0.31 ** | |
| E12.5 | 1X | 0.65 ± 0.06 | 0.25 ± 0.03 | 5.34 ± 0.50 |
| 2X | 0.62 ± 0.06 | 0.23 ± 0.03 | 5.59 ± 0.51 | |
| 4X | 0.57 ± 0.05 | 0.25 ± 0.02 | 6.33 ± 0.42 # | |
| E15.5 | 1X | 0.90 ± 0.06 | 0.19 ± 0.02 | 7.99 ± 0.40 |
| 2X | 0.94 ± 0.06 | 0.21 ± 0.02 | 8.74 ± 0.43 | |
| 4X | 0.87 ± 0.07 | 0.19 ± 0.02 | 8.27 ± 0.48 | |
| E18.5 | 1X | 1.08 ± 0.08 | 0.20 ± 0.02 | 7.66 ± 0.46 |
| 2X | 1.14 ± 0.07 | 0.18 ± 0.02 | 7.42 ± 0.41 | |
| 4X | 1.03 ± 0.08 | 0.16 ± 0.02 # | 7.80 ± 0.42 |
| E10.5 | E12.5 | E15.5 | E18.5 | |||||
|---|---|---|---|---|---|---|---|---|
| β | p Value | β | p Value | β | p Value | β | p Value | |
| Embryo weight | ||||||||
| Choline | 0.0051 | 0.37 | 0.040 | 0.065 # | −0.171 | 0.021 * | −0.025 | 0.71 |
| Betaine | 0.0019 | 0.0082 * | −0.0011 | 0.61 | 0.018 | 0.040 * | 0.0095 | 0.42 |
| Methionine | −0.0032 | 0.75 | −0.080 | 0.090 # | −0.45 | 0.11 | 0.018 | 0.95 |
| Placenta weight | ||||||||
| Choline | −0.0055 | 0.64 | −0.031 | 0.0025 * | −0.068 | 0.000013 * | −0.00058 | 0.94 |
| Betaine | −0.00053 | 0.721 | −0.00092 | 0.37 | 0.0059 | 0.0013 * | 0.0040 | 0.0015 * |
| Methionine | −0.018 | 0.36 | 0.022 | 0.35 | −0.072 | 0.25 | −0.052 | 0.10 |
| Crown rump length | ||||||||
| Choline | 0.731 | 0.38 | 2.3 | 0.041 * | −0.65 | 0.53 | 2.1 | 0.073 # |
| Betaine | 0.262 | 0.011 * | 0.063 | 0.60 | 0.17 | 0.18 | −0.064 | 0.76 |
| Methionine | −0.012 | 0.99 | −6.3 | 0.010 * | −3.1 | 0.43 | 2.7 | 0.60 |
| Placental efficiency | ||||||||
| Choline | 0.19 | 0.279 | 1.3 | 0.0020 * | 0.33 | 0.63 | −0.018 | 0.86 |
| Betaine | 0.042 | 0.080 # | −0.045 | 0.30 | −0.017 | 0.84 | −0.43 | 0.010 * |
| Methionine | −0.039 | 0.90 | −0.23 | 0.018 * | −1.8 | 0.49 | 6.4 | 0.13 |
| E10.5 | E18.5 | |||
|---|---|---|---|---|
| β | p Value | β | p Value | |
| Embryo weight | ||||
| Igf1 | −0.0063 | 0.0072 * | −0.045 | 0.15 |
| Igf2 | 0.00012 | 0.26 | −0.00015 | 0.26 |
| Igf1r | 0.0021 | 0.55 | 0.072 | 0.12 |
| Igf2r | −0.00025 | 0.84 | −0.12 | 0.11 |
| Egfr | −0.020 | 0.0019 * | 0.14 | 0.077 # |
| Placenta weight | ||||
| Igf1 | −0.0043 | 0.33 | −0.00093 | 0.82 |
| Igf2 | 0.070 | 0.71 | 0.000013 | 0.43 |
| Igf1r | 0.0084 | 0.20 | −0.00033 | 0.59 |
| Igf2r | −0.0014 | 0.51 | −0.0014 | 0.14 |
| Egfr | 0.00065 | 0.096 # | −0.000080 | 0.99 |
| Crown rump length | ||||
| Igf1 | −0.64 | 0.043 * | −0.62 | 0.20 |
| Igf2 | −0.18 | 0.21 | −0.0013 | 0.54 |
| Igf1r | 0.072 | 0.88 | 0.80 | 0.28 |
| Igf2r | −0.17 | 0.33 | −0.20 | 0.099 # |
| Egfr | −1.8 | 0.033 * | 2.0 | 0.13 |
| Placental efficiency | ||||
| Igf1 | −0.88 | 0.24 | −0.29 | 0.57 |
| Igf2 | 0.0028 | 0.39 | −0.00019 | 0.93 |
| Igf1r | −0.10 | 0.36 | 1.1 | 0.17 |
| Igf2r | −0.020 | 0.60 | 0.018 | 0.89 |
| Egfr | −0.57 | 0.0041 * | 1.3 | 0.36 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
King, J.H.; Kwan, S.T.; Yan, J.; Klatt, K.C.; Jiang, X.; Roberson, M.S.; Caudill, M.A. Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency. Nutrients 2017, 9, 765. https://doi.org/10.3390/nu9070765
King JH, Kwan ST, Yan J, Klatt KC, Jiang X, Roberson MS, Caudill MA. Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency. Nutrients. 2017; 9(7):765. https://doi.org/10.3390/nu9070765
Chicago/Turabian StyleKing, Julia H., Sze Ting (Cecilia) Kwan, Jian Yan, Kevin C. Klatt, Xinyin Jiang, Mark S. Roberson, and Marie A. Caudill. 2017. "Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency" Nutrients 9, no. 7: 765. https://doi.org/10.3390/nu9070765
APA StyleKing, J. H., Kwan, S. T., Yan, J., Klatt, K. C., Jiang, X., Roberson, M. S., & Caudill, M. A. (2017). Maternal Choline Supplementation Alters Fetal Growth Patterns in a Mouse Model of Placental Insufficiency. Nutrients, 9(7), 765. https://doi.org/10.3390/nu9070765