Zeolite-Containing Mixture Supplementation Ameliorated Dextran Sodium Sulfate-Induced Colitis in Mice by Suppressing the Inflammatory Bowel Disease Pathway and Improving Apoptosis in Colon Mucosa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Dietary Treatment
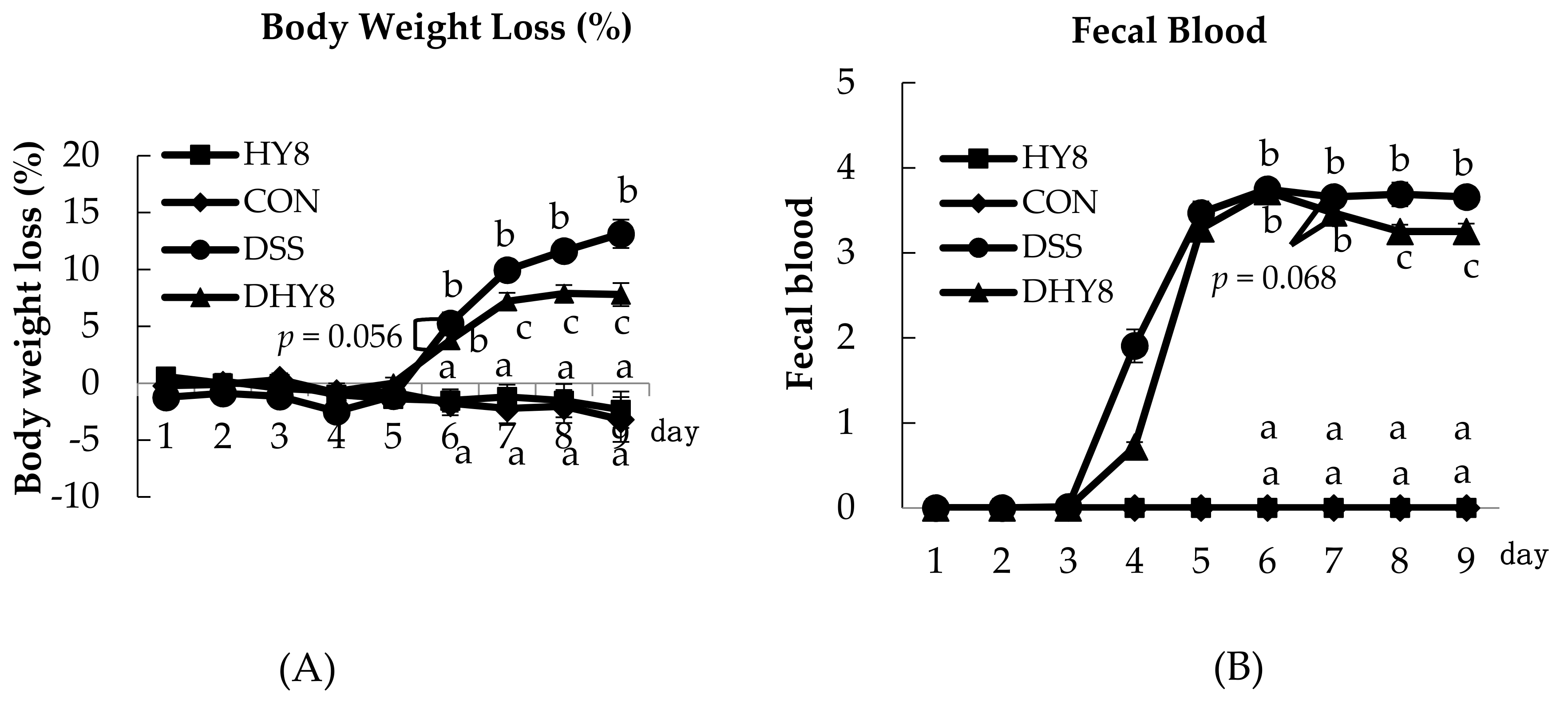
2.2. Evaluation of the Disease Activity Index
2.3. Blood Collection and Tissue Harvesting
2.4. Biochemical Assays
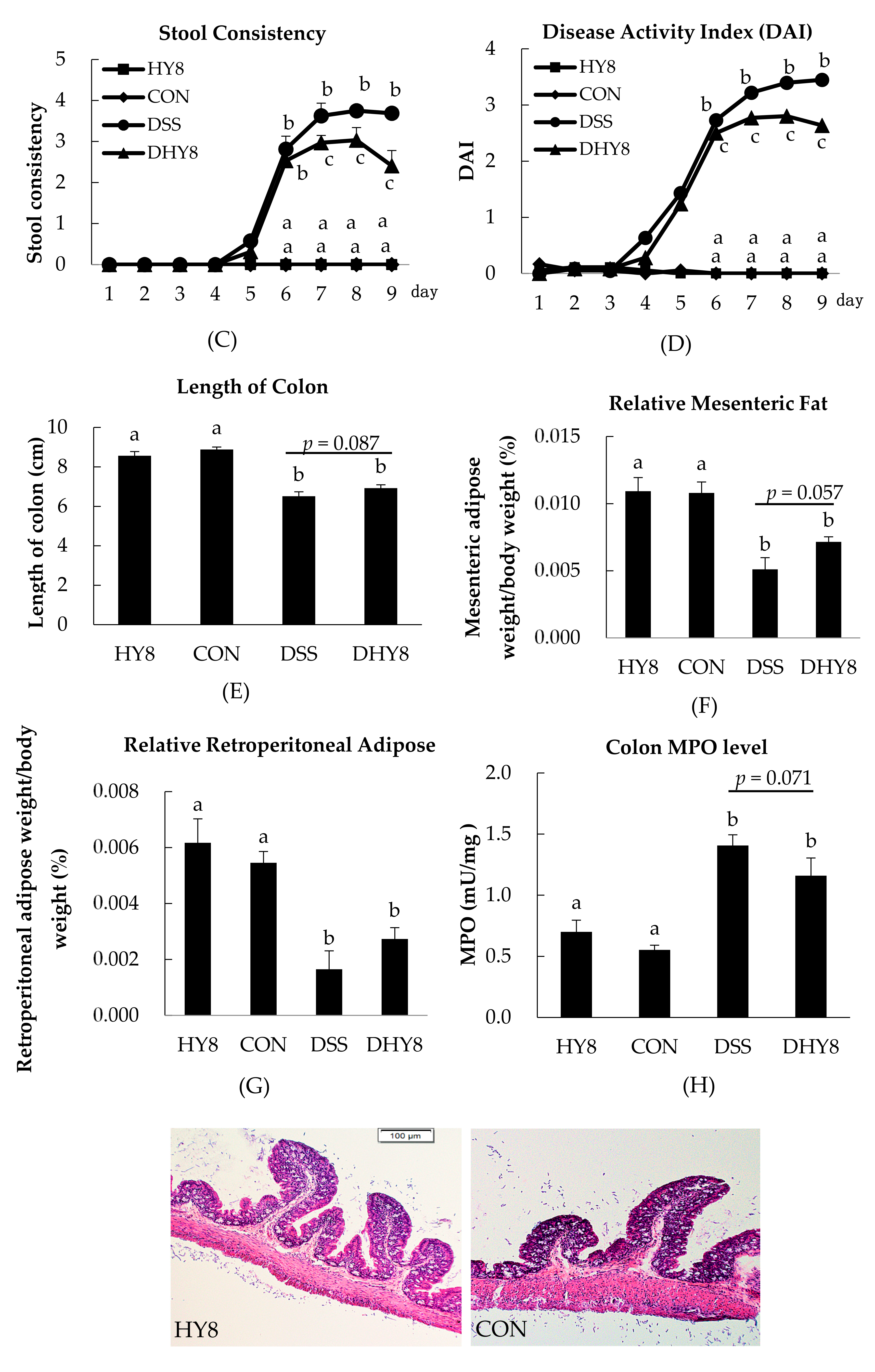
2.5. Colon Histology
2.6. Total RNA Extraction and Quality Assessment
2.7. Transcriptome Analysis
2.7.1. DNA Microarray Preparation
2.7.2. Mapping and Functional Analysis
2.8. Reverse Transcription-Polymerase Chain Reaction
2.9. Protein Preparation, iTRAQ Labeling, and NanoLC-MS/MS Analysis for Proteome Analysis
2.10. Statistical Analysis
3. Results
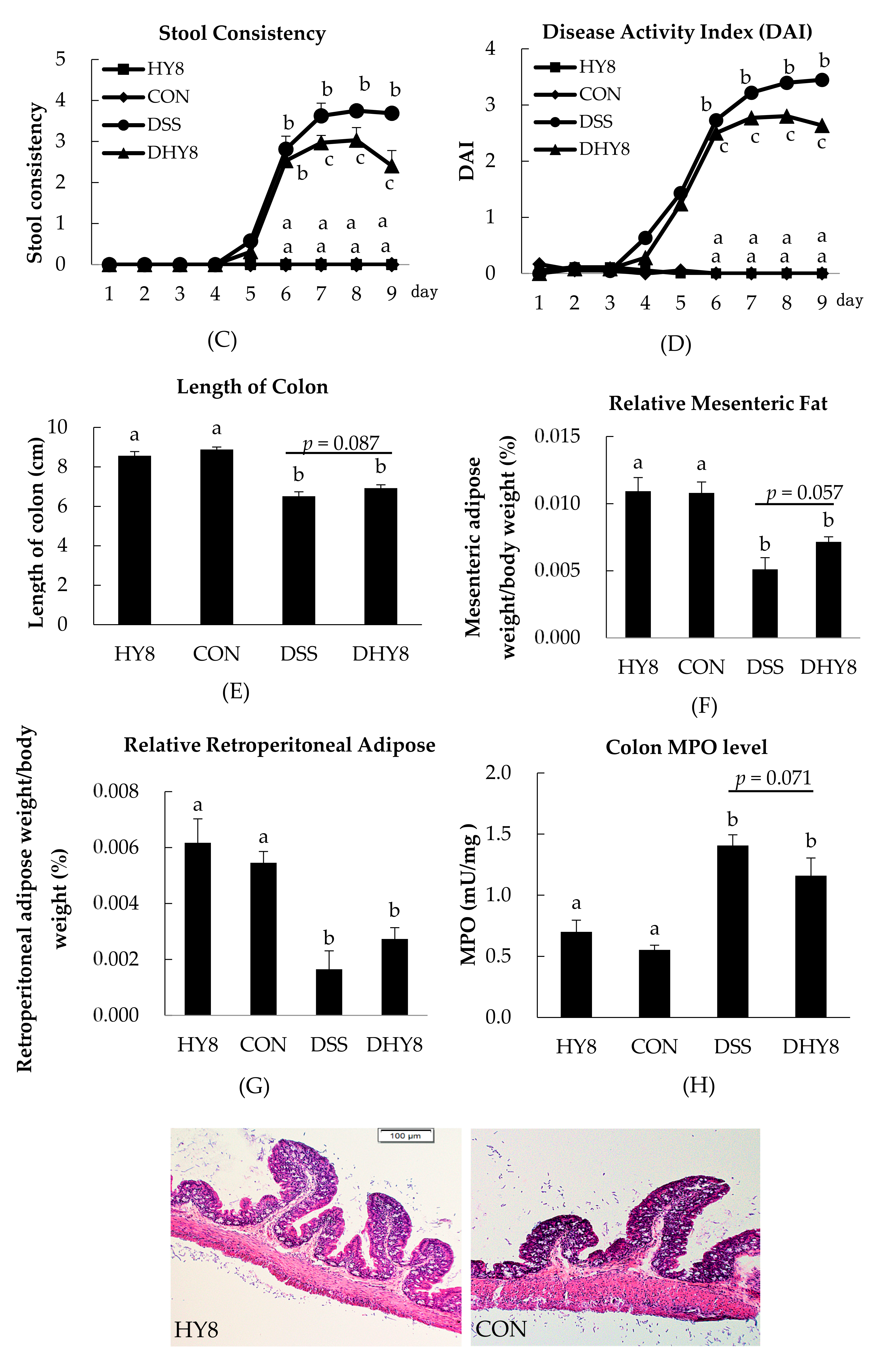
3.1. General Characteristics
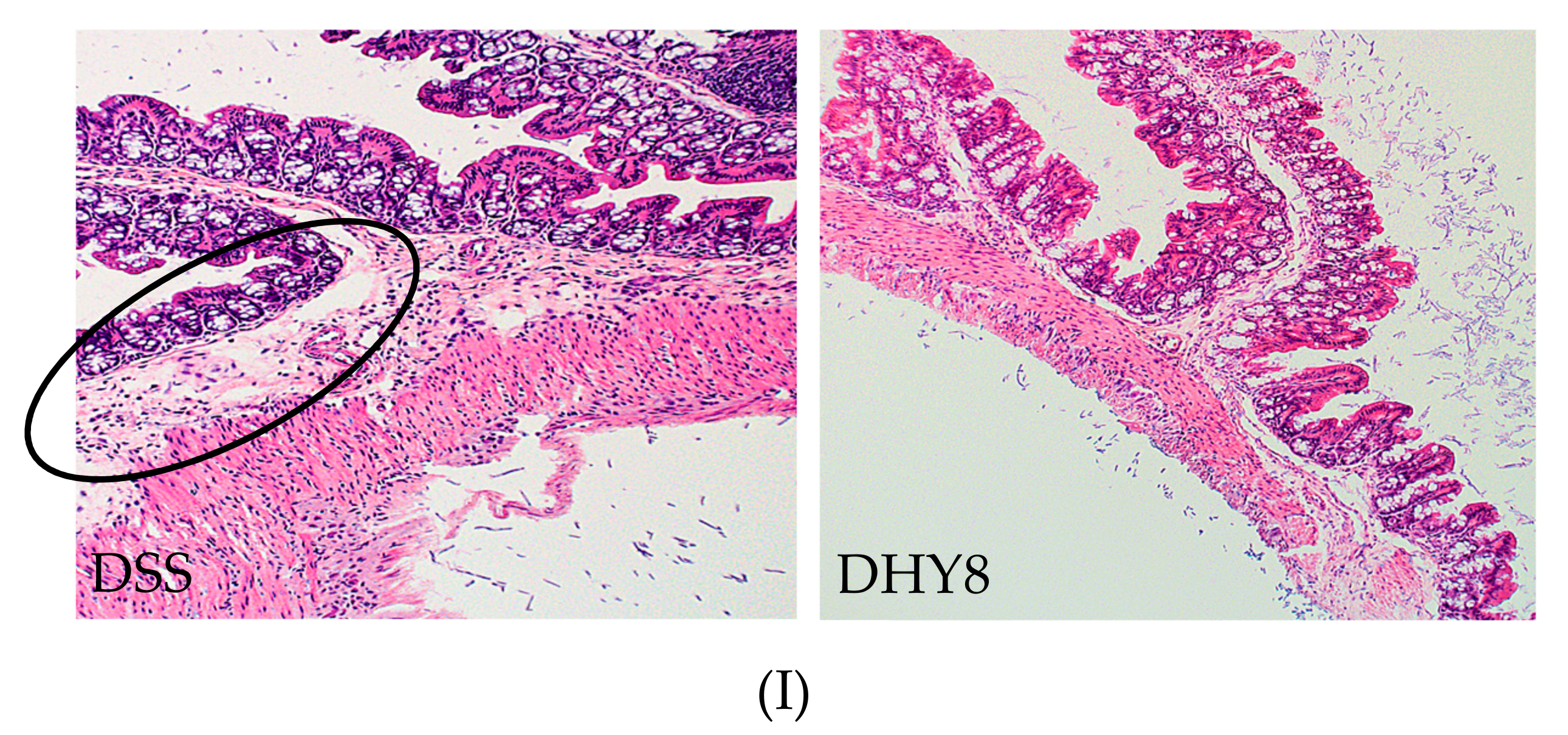
3.2. Colon Histology
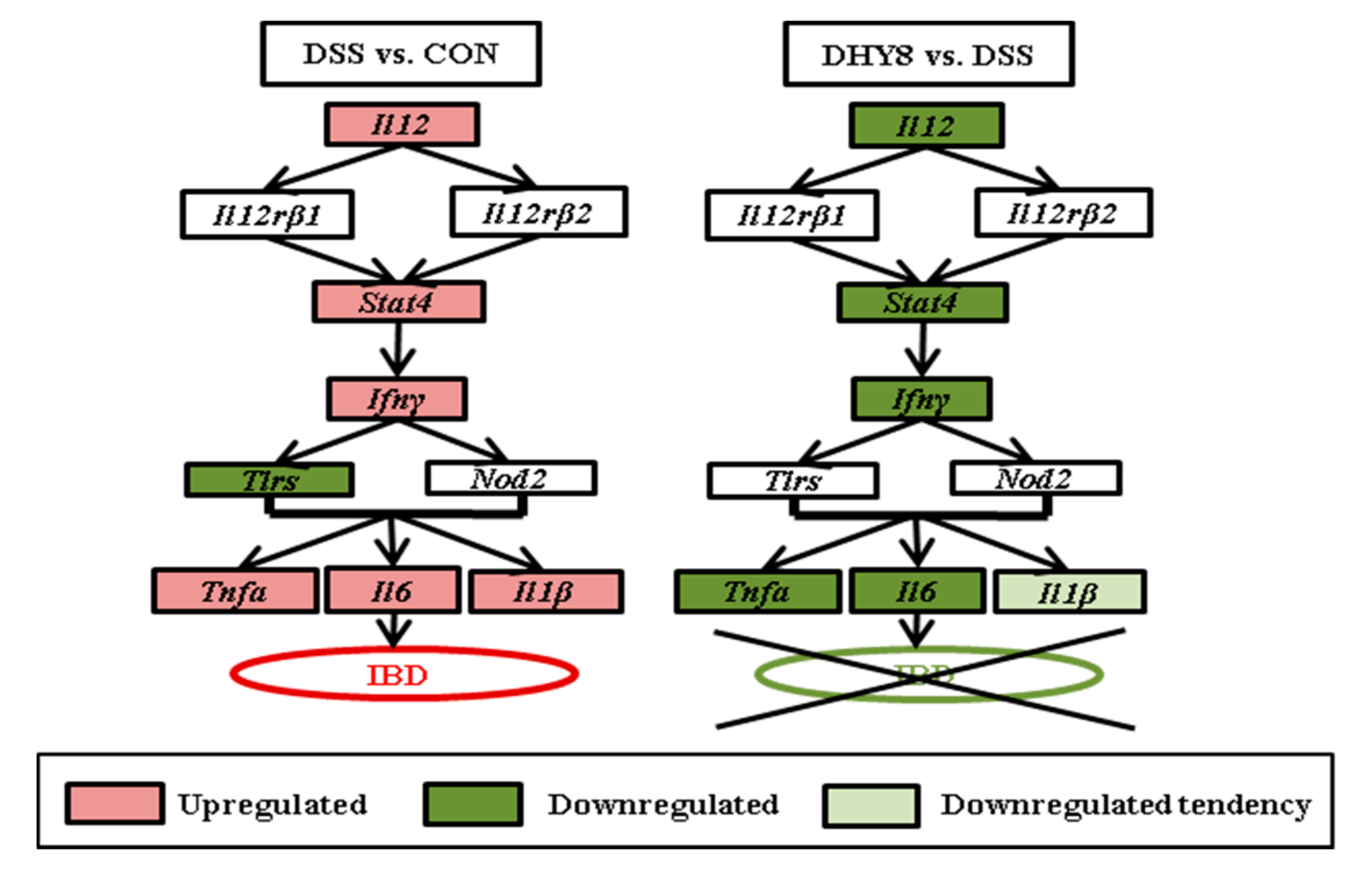
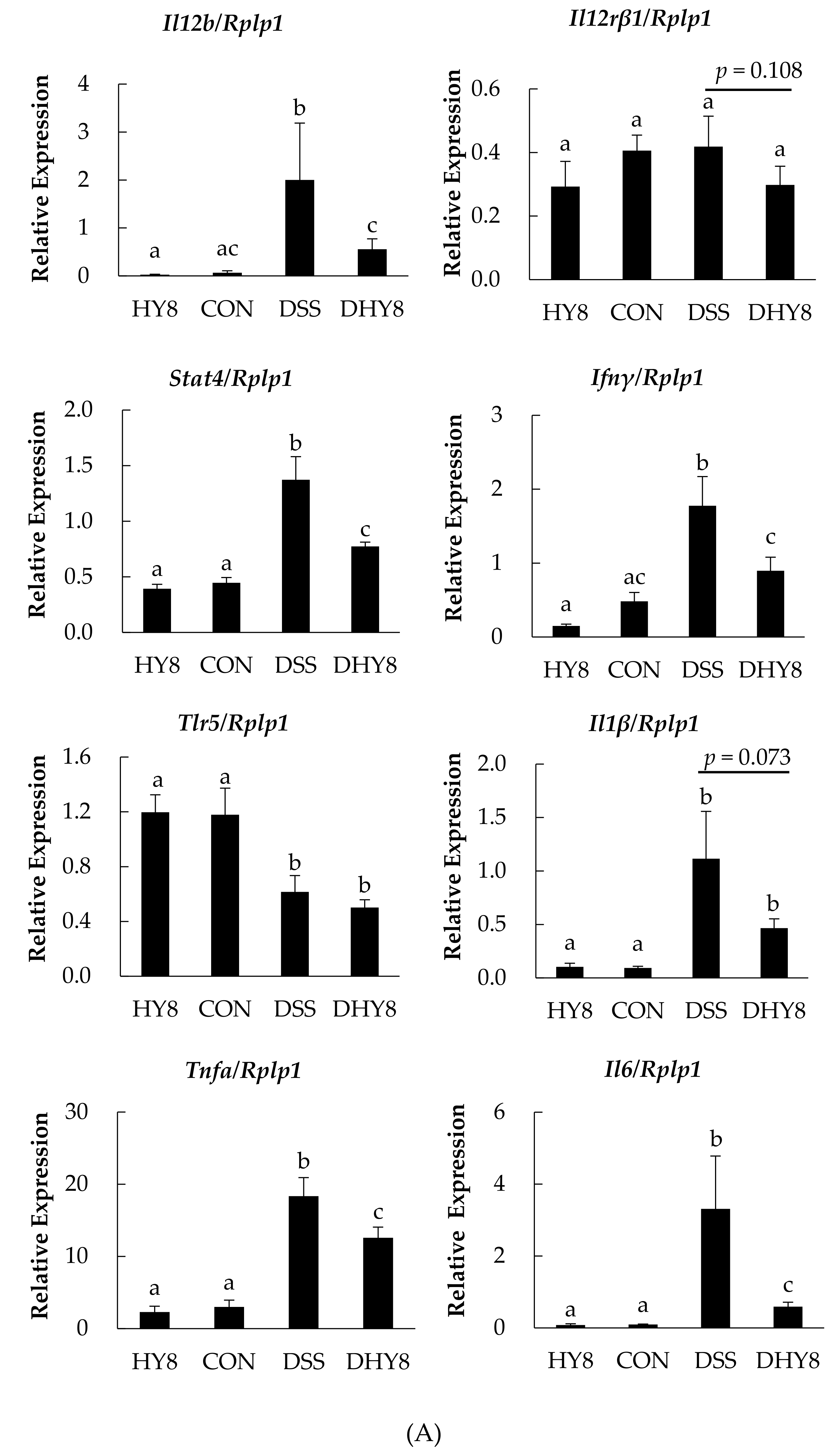
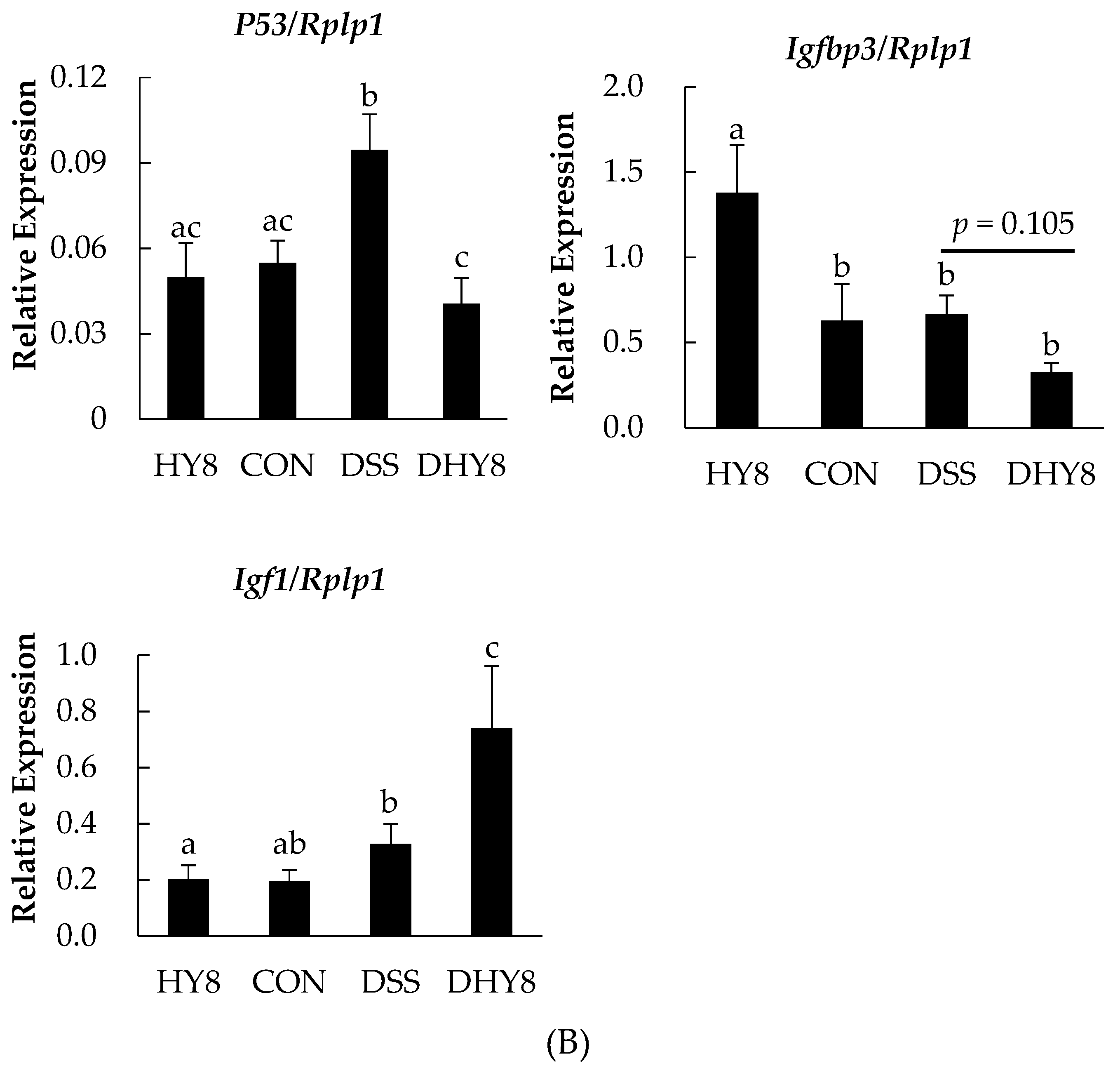
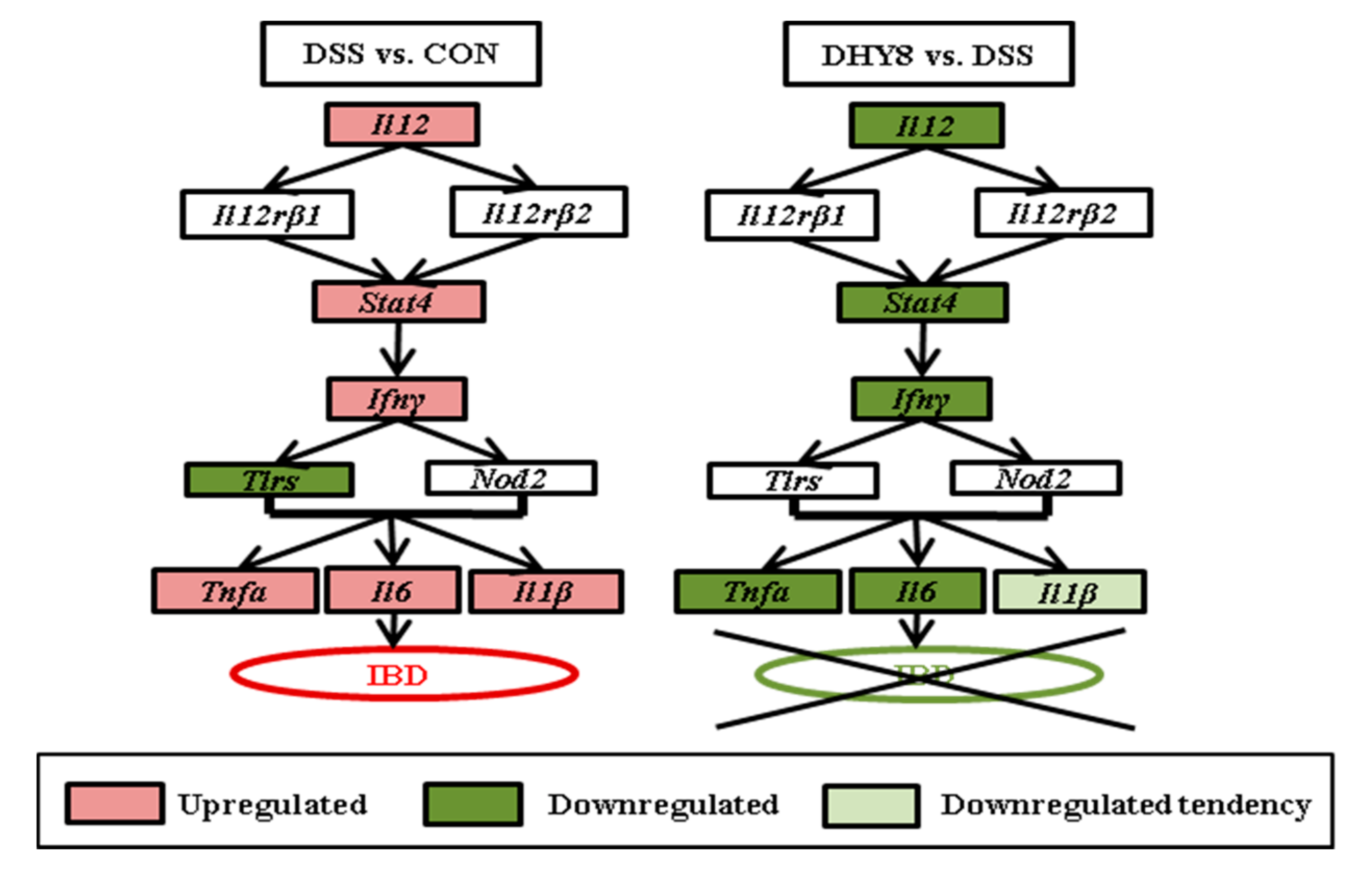
3.3. Colonic Microarray Analysis
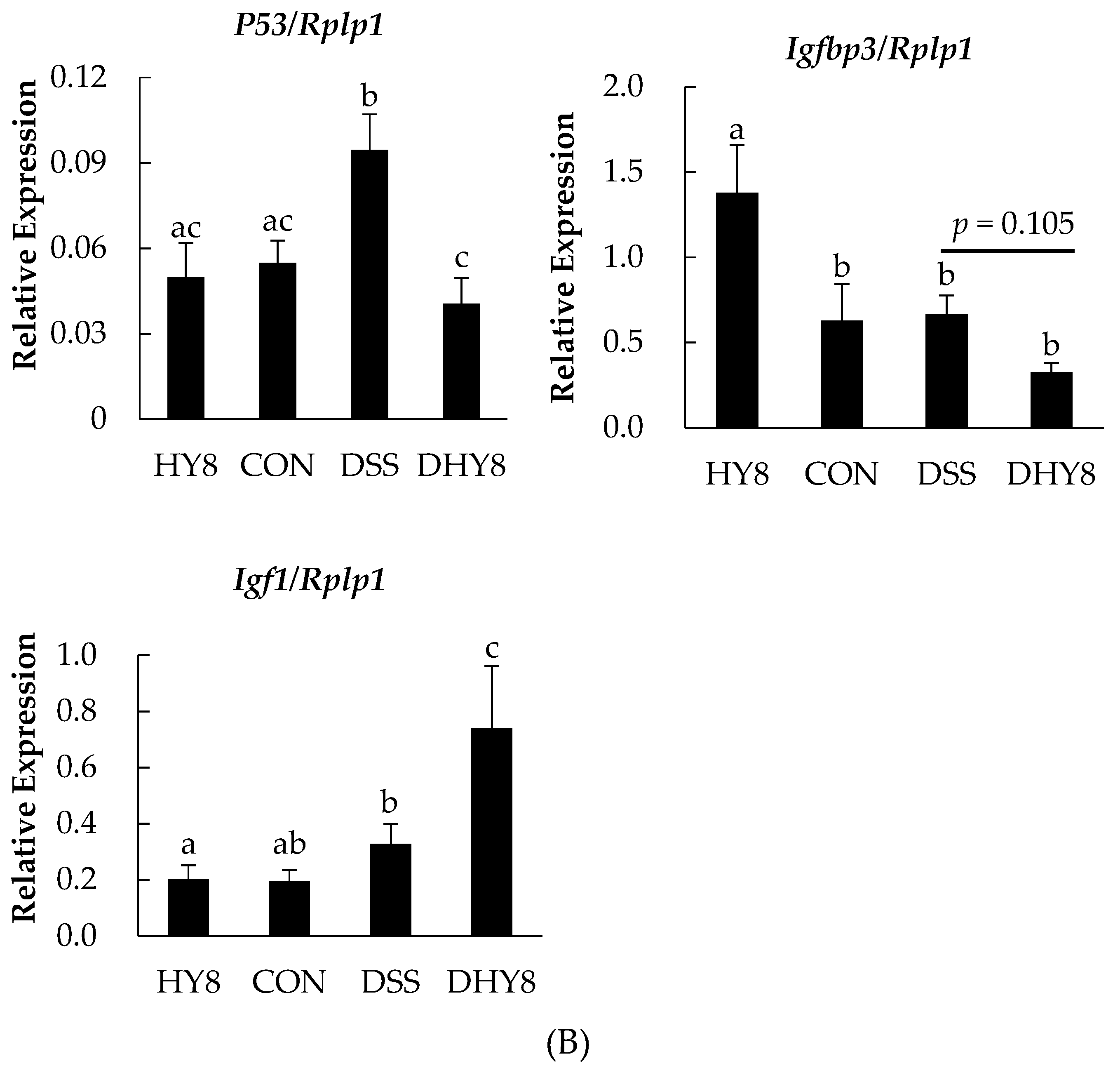
3.4. Comparative Proteomic Analysis by iTRAQ
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflict of Interest
References
- Jakubowski, A.; Zagorowicz, E.; Kraszewska, E.; Bartnik, W. Rising hospitalization rates for inflammatory bowel disease in Poland. Pol. Arch. Med. Wewn. 2014, 124, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Richman, E.; Rhodes, J.M. Review article: Evidence-based dietary advice for patients with inflammatory bowel disease. Aliment. Pharmacol. Ther. 2013, 38, 1156–1171. [Google Scholar] [CrossRef] [PubMed]
- Owczarek, D.; Rodacki, T.; Domagala-Rodacka, R.; Cibor, D.; Mach, T. Diet and nutritional factors in inflammatory bowel diseases. World J. Gastroenterol. 2016, 22, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Tondar, M.; Parsa, M.J.; Yousefpour, Y.; Sharifi, A.M.; Shetab-Boushehri, S.V. Feasibility of clinoptilolite application as a microporous carrier for ph-controlled oral delivery of aspirin. Acta Chim. Slov. 2014, 61, 688–693. [Google Scholar] [PubMed]
- Li, L.; Li, P.; Chen, Y.; Wen, C.; Zhuang, S.; Zhou, Y. Zinc-bearing zeolite clinoptilolite improves tissue zinc accumulation in laying hens by enhancing zinc transporter gene mrna abundance. Anim. Sci. J. 2015, 86, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Katsoulos, P.D.; Zarogiannis, S.; Roubies, N.; Christodoulopoulos, G. Effect of long-term dietary supplementation with clinoptilolite on performance and selected serum biochemical values in dairy goats. Am. J. Vet. Res. 2009, 70, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Y.; Ye, Q.; Li, J.; Hua, Y.; Ju, D.; Zhang, D.; Cooper, R.; Chang, M. Constituents of red yeast rice, a traditional chinese food and medicine. J. Agric. Food Chem. 2000, 48, 5220–5225. [Google Scholar] [CrossRef] [PubMed]
- Lyu, W.; Jia, H.; Kato, H.; The University of Tokyo, Tokyo, Japan. Unpublished work. 2015.
- McHenga, S.S.; Wang, D.; Li, C.; Shan, F.; Lu, C. Inhibitory effect of recombinant il-25 on the development of dextran sulfate sodium-induced experimental colitis in mice. Cell. Mol. Immunol. 2008, 5, 425–431. [Google Scholar] [CrossRef] [PubMed]
- IPA. Available online: https://www.qiagenbioinformatics.com (accessed on 13 April 2016).
- Thierfelder, W.E.; van Deursen, J.M.; Yamamoto, K.; Tripp, R.A.; Sarawar, S.R.; Carson, R.T.; Sangster, M.Y.; Vignali, D.A.; Doherty, P.C.; Grosveld, G.C.; et al. Requirement for stat4 in interleukin-12-mediated responses of natural killer and t cells. Nature 1996, 382, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Dietze, K.K.; Gibbert, K.; Lang, K.S.; Trilling, M.; Yan, H.; Wu, J.; Yang, D.; Lu, M.; Roggendorf, M.; et al. Tlr ligand induced il-6 counter-regulates the anti-viral cd8(+) t cell response during an acute retrovirus infection. Sci. Rep. 2015, 5, 10501. [Google Scholar] [CrossRef] [PubMed]
- Ohama, T.; Hori, M.; Momotani, E.; Iwakura, Y.; Guo, F.; Kishi, H.; Kobayashi, S.; Ozaki, H. Intestinal inflammation downregulates smooth muscle cpi-17 through induction of tnf-alpha and causes motility disorders. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1429–G1438. [Google Scholar] [CrossRef] [PubMed]
- Ohama, T.; Hori, M.; Sato, K.; Ozaki, H.; Karaki, H. Chronic treatment with interleukin-1beta attenuates contractions by decreasing the activities of cpi-17 and mypt-1 in intestinal smooth muscle. J. Biol. Chem. 2003, 278, 48794–48804. [Google Scholar] [CrossRef] [PubMed]
- Vanuytsel, T.; Vermeire, S.; Cleynen, I. The role of haptoglobin and its related protein, zonulin, in inflammatory bowel disease. Tissue Barriers 2013, 1, e27321. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Sakurai, T.; Kashida, H.; Mine, H.; Hagiwara, S.; Matsui, S.; Yoshida, K.; Nishida, N.; Watanabe, T.; Itoh, K.; et al. Involvement of heat shock protein a4/apg-2 in refractory inflammatory bowel disease. Inflamm. Bowel Dis. 2015, 21, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Azuma, Y.T.; Hagi, K.; Shintani, N.; Kuwamura, M.; Nakajima, H.; Hashimoto, H.; Baba, A.; Takeuchi, T. Pacap provides colonic protection against dextran sodium sulfate induced colitis. J. Cell. Physiol. 2008, 216, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Li, Y.Y.; Wang, Y.; Han, G.C.; Wang, R.X.; Xiao, H.; Li, X.Y.; Hou, C.M.; Ma, Y.F.; Sheng, D.S.; et al. Complement activation promotes colitis-associated carcinogenesis through activating intestinal il-1beta/il-17a axis. Mucosal Immunol. 2015, 8, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Tanida, S.; Mizoshita, T.; Ozeki, K.; Katano, T.; Kataoka, H.; Kamiya, T.; Joh, T. Advances in refractory ulcerative colitis treatment: A new therapeutic target, annexin a2. World J. Gastroenterol. 2015, 21, 8776–8786. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, L.; Chi, B. Upregulation of periostin prevents p53-mediated apoptosis in sgc-7901 gastric cancer cells. Mol. Biol. Rep. 2013, 40, 1677–1683. [Google Scholar] [CrossRef] [PubMed]
- Spehlmann, M.E.; Manthey, C.F.; Dann, S.M.; Hanson, E.; Sandhu, S.S.; Liu, L.Y.; Abdelmalak, F.K.; Diamanti, M.A.; Retzlaff, K.; Scheller, J.; et al. Trp53 deficiency protects against acute intestinal inflammation. J. Immunol. 2013, 191, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Takahashi, T.; Yasuhara, N.; Inazawa, J.; Kamada, S.; Tsujimoto, Y. Bis, a bcl-2-binding protein that synergizes with bcl-2 in preventing cell death. Oncogene 1999, 18, 6183–6190. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.D.; Yoon, J.S.; Yoon, H.H.; Youn, H.J.; Kim, J.; Lee, J.H. Expression of bis in the mouse gastrointestinal system. Anat. Cell Biol. 2012, 45, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Hjelm, B.; Brennan, D.J.; Zendehrokh, N.; Eberhard, J.; Nodin, B.; Gaber, A.; Ponten, F.; Johannesson, H.; Smaragdi, K.; Frantz, C.; et al. High nuclear rbm3 expression is associated with an improved prognosis in colorectal cancer. Proteom. Clin. Appl. 2011, 5, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Sureban, S.M.; Ramalingam, S.; Natarajan, G.; May, R.; Subramaniam, D.; Bishnupuri, K.S.; Morrison, A.R.; Dieckgraefe, B.K.; Brackett, D.J.; Postier, R.G.; et al. Translation regulatory factor rbm3 is a proto-oncogene that prevents mitotic catastrophe. Oncogene 2008, 27, 4544–4556. [Google Scholar] [CrossRef] [PubMed]
- Pham, C.G.; Bubici, C.; Zazzeroni, F.; Papa, S.; Jones, J.; Alvarez, K.; Jayawardena, S.; De Smaele, E.; Cong, R.; Beaumont, C.; et al. Ferritin heavy chain upregulation by nf-kappab inhibits tnfalpha-induced apoptosis by suppressing reactive oxygen species. Cell 2004, 119, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Bardia, A.; Tiwari, S.K.; Gunisetty, S.; Anjum, F.; Nallari, P.; Habeeb, M.A.; Khan, A.A. Functional polymorphisms in xrcc-1 and ape-1 contribute to increased apoptosis and risk of ulcerative colitis. Inflamm. Res. 2012, 61, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Mort, R.; Mo, L.; McEwan, C.; Melton, D.W. Lack of involvement of nucleotide excision repair gene polymorphisms in colorectal cancer. Br. J. Cancer 2003, 89, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.J.; Deng, D.J.; Che, J.; Zhao, H.R.; Yu, J.J.; Lu, Y.Y. Hypothalamic paraventricular nucleus stimulation reduces intestinal injury in rats with ulcerative colitis. World J. Gastroenterol. 2016, 22, 3769–3776. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Cadrin, M.; Herrmann, H.; Chen, C.H.; Chalkley, R.J.; Burlingame, A.L.; Omary, M.B. Keratin 20 serine 13 phosphorylation is a stress and intestinal goblet cell marker. J. Biol. Chem. 2006, 281, 16453–16461. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Horiuchi, K.; Sasaki, A.; Tsukamoto, T.; Okabayashi, K.; Hasegawa, H.; Kitagawa, Y.; Okada, Y. Epithelial cell-derived a disintegrin and metalloproteinase-17 confers resistance to colonic inflammation through egfr activation. EBioMedicine 2016, 5, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Judd, L.M.; Chalinor, H.V.; Walduck, A.; Pavlic, D.I.; Dabritz, J.; Dubeykovskaya, Z.; Wang, T.C.; Menheniott, T.R.; Giraud, A.S. Tff2 deficiency exacerbates weight loss and alters immune cell and cytokine profiles in dss colitis, and this cannot be rescued by wild-type bone marrow. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G12–G24. [Google Scholar] [CrossRef] [PubMed]
- Kurt-Jones, E.A.; Cao, L.; Sandor, F.; Rogers, A.B.; Whary, M.T.; Nambiar, P.R.; Cerny, A.; Bowen, G.; Yan, J.; Takaishi, S.; et al. Trefoil family factor 2 is expressed in murine gastric and immune cells and controls both gastrointestinal inflammation and systemic immune responses. Infect. Immun. 2007, 75, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Furth, E.E.; Li, J.; Purev, E.; Solomon, A.C.; Rogler, G.; Mick, R.; Putt, M.; Zhang, T.; Somasundaram, R.; Swoboda, R.; et al. Serum antibodies to epcam in healthy donors but not ulcerative colitis patients. Cancer Immunol. Immunother. 2006, 55, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Balzar, M.; Prins, F.A.; Bakker, H.A.; Fleuren, G.J.; Warnaar, S.O.; Litvinov, S.V. The structural analysis of adhesions mediated by ep-cam. Exp. Cell Res. 1999, 246, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Kusakabe, M.; Horiguchi, K.; Iino, S.; Nakamura, T.; Iwanaga, K.; Hashimoto, H.; Matsumoto, S.; Murata, T.; Hori, M.; et al. Pdgf and tgf-beta promote tenascin-c expression in subepithelial myofibroblasts and contribute to intestinal mucosal protection in mice. Br. J. Pharmacol. 2014, 171, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, C.R.; Chiquet-Ehrismann, R.; Alkan, S.S. Tenascin, an extracellular matrix protein, exerts immunomodulatory activities. Proc. Natl. Acad. Sci. USA 1989, 86, 7437–7441. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, L.; Tian, Y.; Ye, J.; Liu, Y.; Song, L.; Pan, Q.; He, Y.; Chen, W.; Peng, Z.; et al. Numb modulates the paracellular permeability of intestinal epithelial cells through regulating apical junctional complex assembly and myosin light chain phosphorylation. Exp. Cell Res. 2013, 319, 3214–3225. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control Diet (AIN-93G) | Zeolite-Containing Mixture 0.8% |
|---|---|---|
| Cornstarch | 39.75% | 38.95% |
| Casein (85.7% protein) | 20% | 20% |
| Detrinized cornstarch | 13.2% | 13.2% |
| Sucrose | 10.25% | 10.25% |
| Soybean oil (no additives) | 7% | 7% |
| Fiber | 5% | 5% |
| Mineral mix (AIN 93G-MX) | 3.5% | 3.5% |
| Vitamin mix (AIN 93G-MX) | 1% | 1% |
| L-Cystine | 0.3% | 0.3% |
| Zeolite-containing mixture (HY) | 0 | 0.8% |
| Accession No. | Protein Symbol | Protein Name | Gene Symbol | Fold Change | Fold Change |
|---|---|---|---|---|---|
| DSS vs. CON | DHY8 vs. DSS | ||||
| Q62009-5 | Postn | Isoform 5 of Periostin | Postn | 0.31 | 2.36 |
| Q9D8I1 | Perp1 | Plasma cell-induced resident endoplasmic reticulum protein | Pacap | 0.13 | 5.01 |
| Q9CQW5 | Leg2 | Galectin-2 | Lgals2 | 0.62 | 3.02 |
| Q9JLV1 | Bag3 | BAG family molecular chaperone regulator 3 | Bag3 | 0.67 | 2.31 |
| O89086 | Rbm3 | Putative RNA-binding protein 3 | Rbm3 | 0.43 | 4.25 |
| Q8VHX6-2 | Fhc | Ferritin heavy chain | Fhc | 0.37 | 2.99 |
| Q91VC7 | Pp14a | Protein phosphatase 1 regulatory subunit 14A | Ppp1r14a | 0.33 | 6.43 |
| Q99JW5 | Epcam | Epithelial cell adhesion molecule | Epcam | 0.32 | 3.19 |
| Q80YX1-5 | Tena | Isoform 5 of Tenascin | Tnc | 0.65 | 2.31 |
| Q9QZS3-3 | Numb | Isoform 71 kDa of Protein numb homolog | Numb | 0.44 | 4.17 |
| Q61646 | Hpt | Haptoglobin | Hp | 36.31 | 0.32 |
| P01027 | Co3 | Complement C3 | C3 | 6.55 | 0.34 |
| P11247 | Perm | Myeloperoxidase | Mpo | 5.50 | 0.55 |
| Q61316 | Hsp74 | Heat shock 70 kDa protein 4 | Hspa4 | 1.89 | 0.10 |
| Q9D312 | K1c20 | Keratin, type I cytoskeletal 20 | Krt20 | 1.98 | 0.50 |
| P28352 | Apex1 | DNA-(apurinic or apyrimidinic site) lyase | Apex1 | 1.92 | 0.07 |
| P70677 | Casp3 | Caspase-3 | Casp3 | 1.61 | 0.14 |
| P97864 | Casp7 | Caspase-7 | Casp7 | 6.61 | 0.42 |
| Q8VEK3 | Hnrpu | Heterogeneous nuclear ribonucleoprotein U | Hnrnpu | 1.92 | 0.34 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, W.; Jia, H.; Deng, C.; Saito, K.; Yamada, S.; Kato, H. Zeolite-Containing Mixture Supplementation Ameliorated Dextran Sodium Sulfate-Induced Colitis in Mice by Suppressing the Inflammatory Bowel Disease Pathway and Improving Apoptosis in Colon Mucosa. Nutrients 2017, 9, 467. https://doi.org/10.3390/nu9050467
Lyu W, Jia H, Deng C, Saito K, Yamada S, Kato H. Zeolite-Containing Mixture Supplementation Ameliorated Dextran Sodium Sulfate-Induced Colitis in Mice by Suppressing the Inflammatory Bowel Disease Pathway and Improving Apoptosis in Colon Mucosa. Nutrients. 2017; 9(5):467. https://doi.org/10.3390/nu9050467
Chicago/Turabian StyleLyu, Weida, Huijuan Jia, Chuanzong Deng, Kenji Saito, Seigo Yamada, and Hisanori Kato. 2017. "Zeolite-Containing Mixture Supplementation Ameliorated Dextran Sodium Sulfate-Induced Colitis in Mice by Suppressing the Inflammatory Bowel Disease Pathway and Improving Apoptosis in Colon Mucosa" Nutrients 9, no. 5: 467. https://doi.org/10.3390/nu9050467
APA StyleLyu, W., Jia, H., Deng, C., Saito, K., Yamada, S., & Kato, H. (2017). Zeolite-Containing Mixture Supplementation Ameliorated Dextran Sodium Sulfate-Induced Colitis in Mice by Suppressing the Inflammatory Bowel Disease Pathway and Improving Apoptosis in Colon Mucosa. Nutrients, 9(5), 467. https://doi.org/10.3390/nu9050467