1. Introduction
Polyunsaturated fatty acids (PUFAs) are essential for cell and tissue development and a sufficient supply of PUFA is important from fetal life onwards. The fetus is mainly supplied with PUFAs by transfer from the maternal circulation via the placenta [
1]. After birth, PUFAs are available via the diet, including breast milk or formula in infants and from fatty foods later in life. Important long chain PUFAs, such as arachidonic acid and docosahexaenoic acid (DHA) may also be produced in the body from their essential precursor fatty acids linoleic acid (18:2
n-6) and α-linolenic acid (18:3
n-3) that are abundant in the diet [
2]. Substrate fatty acids are elongated by sequential addition of two-carbon atom units and desaturated by introduction of a double bond in the molecule. The elongation step is catalyzed by elongases encoded by the
ELOVL (elongation of very long chain fatty acids) gene family on chromosome 6 [
3], while desaturation is catalyzed by desaturases such as Δ-5 and Δ-6 desaturases, encoded by the
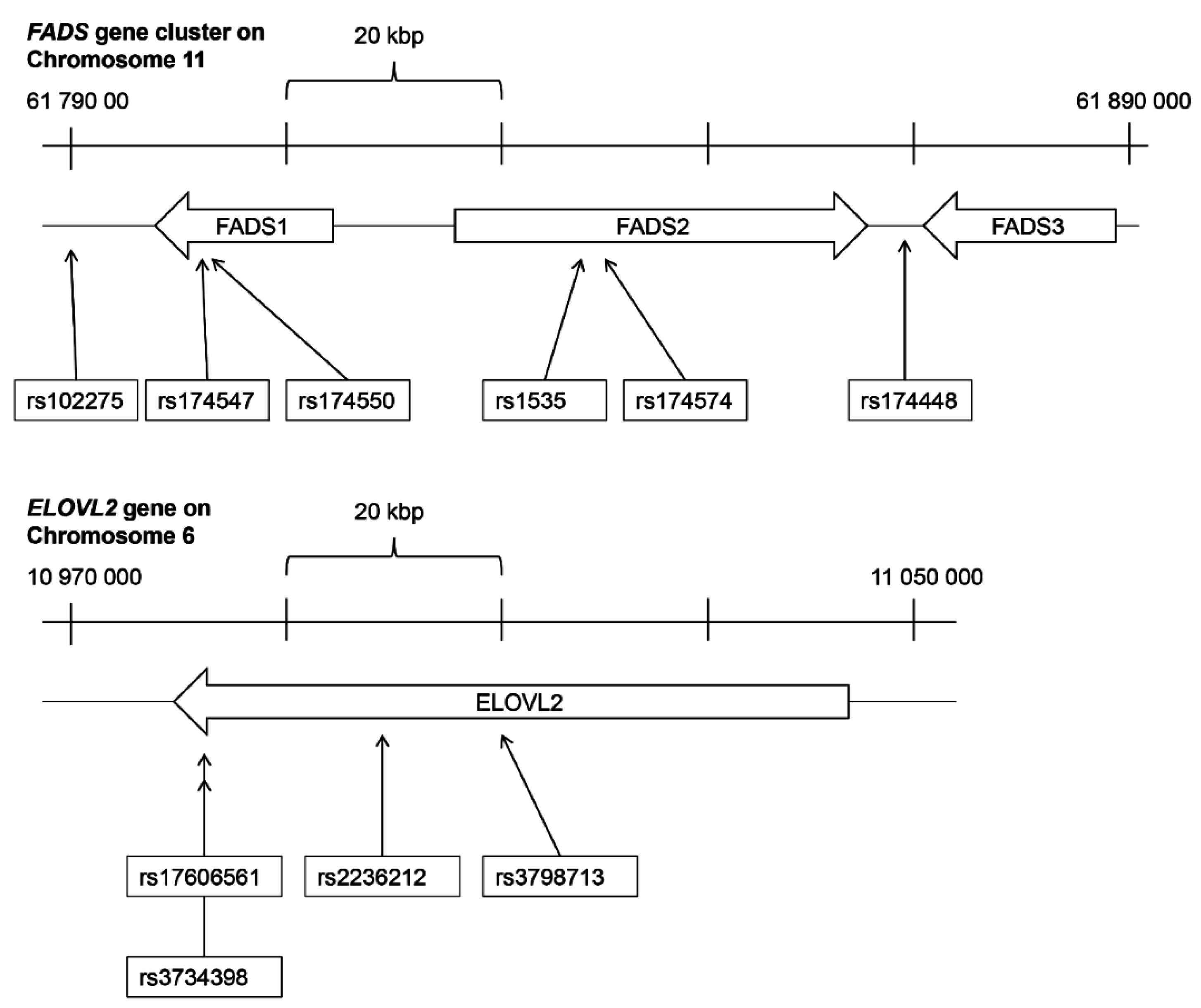
FADS (fatty acid desaturase) gene cluster on chromosome 11 [
4,
5,
6] (
Figure 1). The production of longer
n-6 and longer
n-3 PUFAs involves the same enzymes (elongases and desaturases), hence, there is a competition for the enzymes between the two pathways (
Figure 1). Desaturation is the rate limiting step in this pathway and several studies have revealed that single nucleotide polymorphism (SNPs) in the
FADS gene cluster affect the proportions of PUFA and long chain PUFA in human tissue [
7,
8,
9,
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23]. Polymorphism in the
FADS2 gene that negatively affect the activity of the Δ-6 desaturase, have been associated with increased proportions of linoleic acid (precursor for the
n-6 series) and α-linolenic acid (precursor for the
n-3 series), while the products arachidonic acid (
n-6) and eicosapentaenoic acid (EPA,
n-3) are reduced. Recent genome-wide association studies have also suggested that polymorphism in the
ELOVL2 gene are associated with increased proportions of the substrate and decreased proportions of the products [
23,
24,
25].
Figure 1.
The metabolic pathways of polyunsaturated fatty acids in mammals [
26]. Fatty acids and genes in bold were examined in this study.
Figure 1.
The metabolic pathways of polyunsaturated fatty acids in mammals [
26]. Fatty acids and genes in bold were examined in this study.
PUFAs are important modulators of immune function. Long chain PUFAs are powerful inhibitors of mitogen-induced activation [
27,
28,
29] and secretion of interferon-γ by T cells [
30,
31]. Furthermore, the long chain
n-6 PUFA arachidonic acid is the precursor of prostaglandins [
32] that promote maturation of dendritic cells into a phenotype that favors Th2 lineage commitment from naïve T cells with which the dendritic cell interacts. Th2 cells are central in atopic (IgE-mediated) allergy. We have recently shown that the risk of developing allergy is positively related to a high proportion of both
n-6 and
n-3 long chain PUFAs in cord blood [
33]. Since variation in the
FADS genes affects serum proportions of long chain PUFAs [
24,
25,
34] we speculated that genetic variation in the
FADS genes may also affect the risk of allergy development. Indeed, a German study found that carriers of the minor alleles of several SNPs in
FADS and their respective haplotypes had a lower prevalence of allergic rhinitis and atopic eczema [
13]. The effect of
ELOVL polymorphisms on risk of allergy development has not previously been studied.
The aim of the present study was to evaluate if genetic variations in the FADS gene cluster or in the ELOVL2 gene were associated with proportions of long chain PUFAs in cord serum phospholipids or in adolescents’ serum phospholipids, and to examine any potential association between genetic variation and allergic disease at 13 years of age.
4. Discussion
Both desaturase and elongase enzymes are involved in the endogenous production of long chain PUFAs (≥ 20 carbon atoms) of the
n-6 and
n-3 series from precursors of 18 carbon atom chain-length. To our knowledge, this is the first study to compare the effect of polymorphisms in genes encoding for both these enzymes, the
FADS genes and the
ELOVL2 gene, on fatty acid profiles at two time points, at birth and at 13 years of age in the same children. This is also the first study to analyze the association between allergy development and polymorphism in the
ELOVL2 gene. The results showed that polymorphism in the
FADS genes and, nominally, in the
ELOVL2 gene were associated with the proportions of some
n-6 PUFAs in serum phospholipids. Thus, minor allele carriers were found to have lower proportion of the long-chain product in the serum phospholipids and increased proportions of the substrate. The desaturases and elongases works on both
n-3 and
n-6 PUFAs, but the SNPs investigated here were mainly found to affect the
n-6 PUFA levels, as previously shown [
15,
22].
The potential differential effect of polymorphism on PUFA proportions at the two time points are not assessed. Other factors than polymorphism in the
FADS and
ELOVL genes influence the proportions of PUFA differently at birth and at adolescent state, also difference in storage time between cord serum samples and adolescent serum samples might affect the results of the PUFA measurements. The PUFA levels in cord blood might be partly affected by the efficiency of the transport of PUFAs across the placenta from the maternal to the fetal circulation. While the PUFA levels in adolescence might be more affected by e.g., differences in dietary intake. We have previously shown that fish intake correlated to proportions of
n-3 long chain PUFAs in these subjects at 13 years of age [
36]. In contrast to our findings, a previous study [
22] found stronger correlations between
FADS polymorphisms and long chain PUFA proportions in serum at seven years of age than at birth.
Previous studies have revealed that single nucleotide polymorphism (SNPs) in the
FADS gene cluster affect the proportions of PUFA and long chain PUFA in human tissue [
7,
8,
9,
10,
11,
12,
13,
14,
15,
16,
17,
18,
19,
20,
21,
22,
23]. Polymorphism in the
FADS genes carried by the minority of subjects have generally been associated with enhanced blood proportions of the
n-6 and
n-3 substrate fatty acids and decreased proportions of the product fatty acids, similar to the results in this study. However, the associations between
FADS alleles and cord blood proportions of PUFAs has only been investigated in a single birth-cohort, the ALPSAC cohort from which two studies derive [
15,
22]. Steer
et al. [
22] investigated genetic variation in both the mother and the child in two
FADS2 SNPs, rs1535 and rs174575, in relation to fatty acids at three time points,
i.e., in maternal red blood cells during pregnancy, in cord plasma at birth and in child plasma obtained at seven years of age. Lattka
et al. [
15] analyzed associations between 17 additional SNPs in the whole
FADS gene cluster and cord plasma fatty acid proportions in the same cohort, one of which, rs174448, was analyzed in our study. In both studies investigating the ALPSAC cohort, the minor alleles were associated with reduced amounts of the products and enhanced amounts of the precursors, and stronger associations between
FADS SNPs and the
n-6 series of PUFAs, than with the
n-3 series of PUFAs [
15,
22]. Further, similar to our findings, rs174448 was associated with lower proportions of 20:3
n-6 and higher proportions of 20:4
n-6 and 22:4
n-6 in cord serum phospholipids, but no effect on
n-3 PUFAs [
15].
The pathway of endogenous production of long chain
n-6 and
n-3 PUFAs involves another gene family, elongases, which elongates the fatty acid chain. In the present study two
ELOVL2 SNPs were analyzed: rs17606561 and rs2236212. Minor allele carriers of rs17606561 had decreased proportions of arachidonic acid (20:4
n-6) and minor allele carriers of rs2236212 had a lower ratio of DHA over DPA (22:6
n-3/22:5
n-3) in cord serum. However, none of these associations persisted correction for multiple testing. Similar results, with increased proportions of DPA and decreased proportions of DHA in minor allele carriers of rs2236212 were found by Lematire
et al. [
25] in adults. To our knowledge no previous studies have evaluated the association between
ELOVL2 polymorphism and fatty acid proportions in cord serum phospholipids.
The SNPs in the present study were chosen to represent all SNPs in the
FADS gene cluster or the
ELOVL gene family that have shown significant association to serum phospholipids in two genome-wide association studies [
24,
25]. Lematire
et al. found that minor allele carriers of rs102275 had increased proportions of α-linolenic acid (18:3
n-3) and decreased docosapentaenoic acid (22:5
n-3, DPA) proportions. Minor allele carriers of rs174448 had increased proportions of α-linolenic acid, decreased eicosapentaenoic acid (20:5
n-3, EPA) proportions and decreased docosahexaenoic acid (22:6
n-3, DHA) proportions [
25]. We did not replicate the results found by Lematire
et al. regarding
n-3 PUFAs since we only found significant associations between
n-6 PUFAs and these SNPs. Demirkan
et al. [
24] analyzed the genome-wide association to circulating phospholipid concentrations and found both amounts and proportions of many phospholipids to be associated to
FADS1 and
ELOVL2 polymorphism on a genome wide level. Demirkan
et al. [
24] did not report the fatty acid composition in the phospholipids and our results are therefore not comparable. A recent Genome Wide Association Study showed rs102275 and rs174448 to have a genome wide significant association to a range of long chain fatty acids, e.g., 20:3
n-6 and 20:4
n-6 [
23]. This study also found SNPs in the
ELOVL2 gene to be associated with 22:5
n-3 (DPA) [
23], however none of the
ELOVL SNPs were analyzed in our study.
Two SNPs in the
FADS gene cluster were analyzed here: rs102275 and rs174448. The SNP rs102275 is situated intergenic downstream of
FADS1 and is in strong LD with other SNPs in
FADS1 and
FADS2 genes
. The strongest association of the rs102275 polymorphism was found for the product/precursor ratio for the Δ-5-desaturase in the
n-6 pathway. The other analyzed SNP in the
FADS gene cluster, rs174448, is situated intergenic between
FADS2 and
FADS3. When the two
FADS SNPs were added together in the linear regression models with fatty acids, only rs102275 was significant. The same was true when the haplotypes of rs102275-rs174448 were added. This indicates that the association to rs174448 is just a consequence of the LD with rs102275. These findings are also in accordance with the findings by Ameur
et al. [
44] who found two haplotype blocks in the
FADS region, where rs102275 is situated in their LD block 1 that had the strongest association to long chain PUFAs and rs174448 is in their LD block 2 with weaker association. Additionally, the C and T alleles of rs102275 corresponds to the haplotypes they denoted A and D respectively in block 1 and our association pattern with the
n-6 PUFAs is in agreement with theirs.
The subjects in this study were selected based on their allergic manifestation at 13 years of age. The power of this study may be limited by the relative small population size, however, the prospective birth cohort design allowed us to select very clear cases with only one allergic manifestations as well as non-allergic controls that had not been sensitized nor had any allergic symptoms in any of the follow-ups at 1, 4, 7, or 13 years of age. We aimed for two group of subjects that had allergic symptoms from one organ only,
i.e., atopic eczema or respiratory allergy, since the literature suggests that the association between PUFAs and allergy may differ in different allergic manifestations [
36,
45,
46,
47]. Here, an association between SNPs in the
FADS gene cluster and allergy were found only for subjects with atopic eczema and not for subjects with respiratory allergy. The minor alleles of rs102275 (C) and the minor allele of rs174448 (G) were nominally protective against developing atopic eczema. However, as expected, the significance of rs102275 and rs174448 with atopic eczema disappeared when any of the five fatty acids that are associated with eczema as well as the SNP are added to the logistic model, suggesting that the association between the minor
FADS allele and protection from atopic eczema development acts via a reduced capacity to elongate precursor
n-6 PUFAs to arachidonic acid (20:4
n-6).
The association between SNPs in the
FADS gene cluster and allergic diseases has been reported in three German (ECRHS, LISA, GINI) and one Dutch (KOALA) study with inconsistent results [
13,
17,
48,
49]. Schaeffer
et al. found that adult minor allele carriers of several SNPs had lower prevalence of self-reported allergic rhinitis (
n = 76) and atopic eczema (
n = 49) in the German ECRHS study [
13]. To the contrary, Rzehak
et al. that reported results from the two birth cohort studies, KOALA and LISA, found that minor allele carriers of several SNPs in the
FADS gene cluster had a higher prevalence of parental reported eczema at two years of age (
n = 166) [
17]. However, the association between polymorphism in the
FADS genes and allergy has also been reported for the LISA study when the participants were 6 [
49] and 10 years old [
48], together with subjects from the GINI study. At these two later ages there were no longer any association between polymorphism and asthma, bronchitis, eczema or hay fever at 6 years of age [
49] or at 10 years of age [
48]. Further studies are needed to confirm the protective effects of the minor allele in
FADS genes polymorphism found on atopic eczema in this study and on atopic eczema and allergic rhinitis in the study by Schaeffer
et al. [
13].
We have previously published the association between serum proportions of fatty acids and allergy in the same subjects [
33,
36], this paper additionally adds the polymorphisms. We reported that high proportions of long chain PUFAs at birth, in cord serum, were associated with allergy development [
33]. At 13 years of age, when the allergic disease was manifest, there was no longer any correlation between atopy and serum PUFA proportions [
36], which suggests that the PUFA milieu is important chiefly during early infancy when the naïve immune system of the infant is primed as foreign antigens are encountered. The mechanism by which high proportions of long-chain PUFAs in cord blood increases the risk of allergy development is unknown. Arachidonic acid is a precursor for prostaglandin E
2 (PGE
2) that promotes maturation of dendritic cells into a phenotype that supports Th2 development [
50]. Furthermore, PUFAs counteract T cell activation and production of interferon-gamma; if the immune system is not activated in early infancy because of low microbial stimulation, or a mileu rich in PUFA, immune maturation might be hampered. Interestingly, the other allergic phenotype, respiratory allergy, which was also associated with a higher proportion of long chain PUFAs in cord serum [
33] were not associated with having a different genotype. However, we found the strongest risk of respiratory allergy development to be connected with
n-3 PUFAs [
33] and these were obviously not strongly affected by the gene polymorphisms studied here. This might suggest that
n-3 PUFAs in the fetus are derived chiefly from placental transport from the maternal circulation and are less dependent on synthesis by the fetus, and hence not as strongly associated to genetic variation in the
FADS and
ELOVL genes.

 ,
, 

{kind=link}
{kind=link}