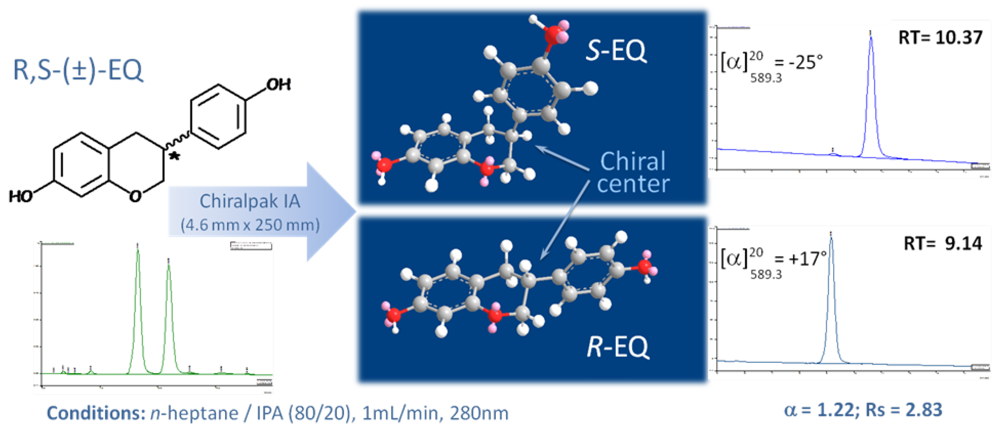
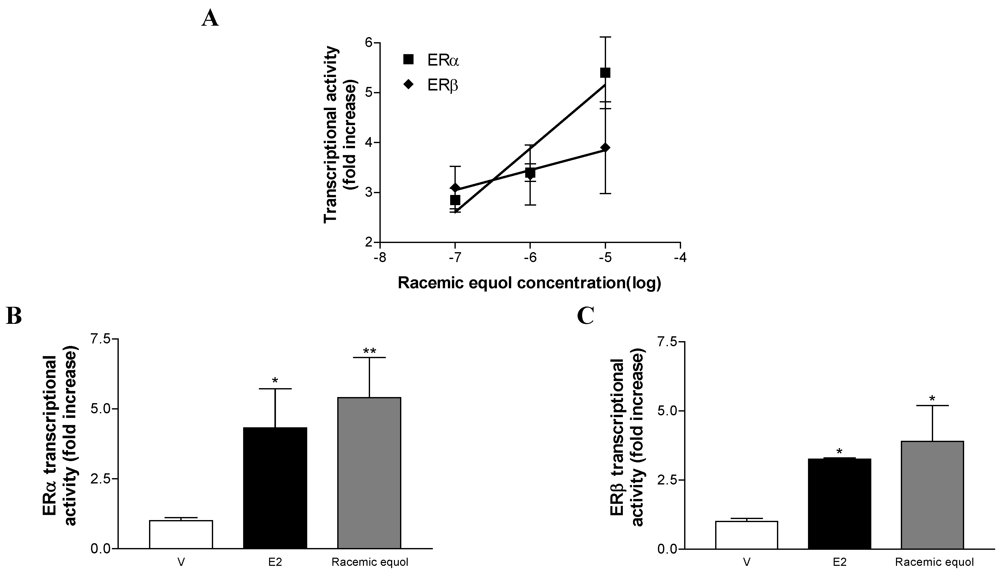
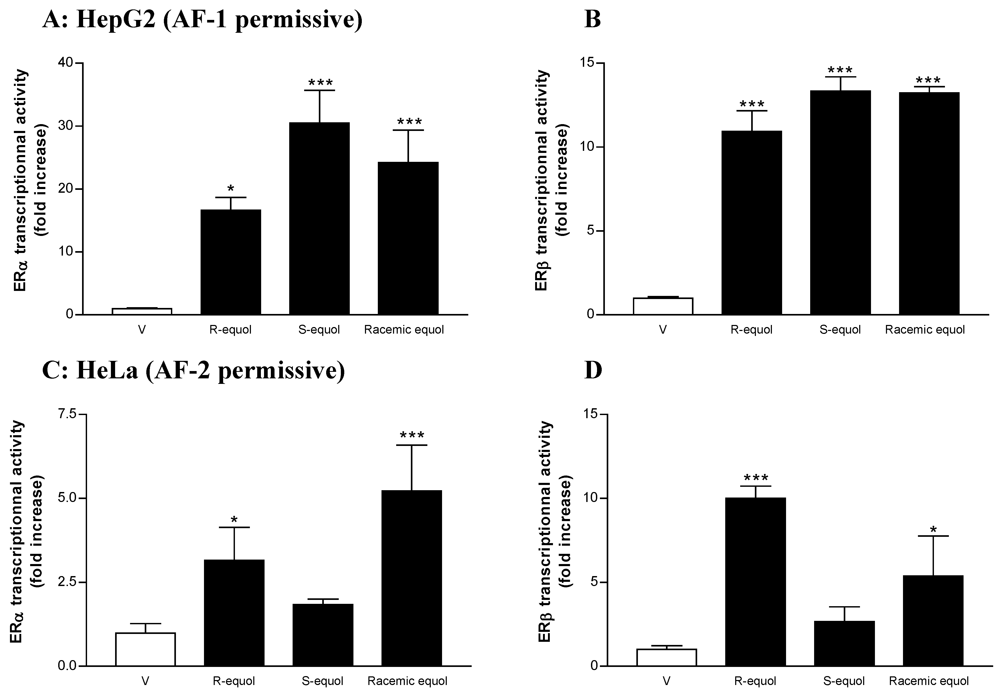
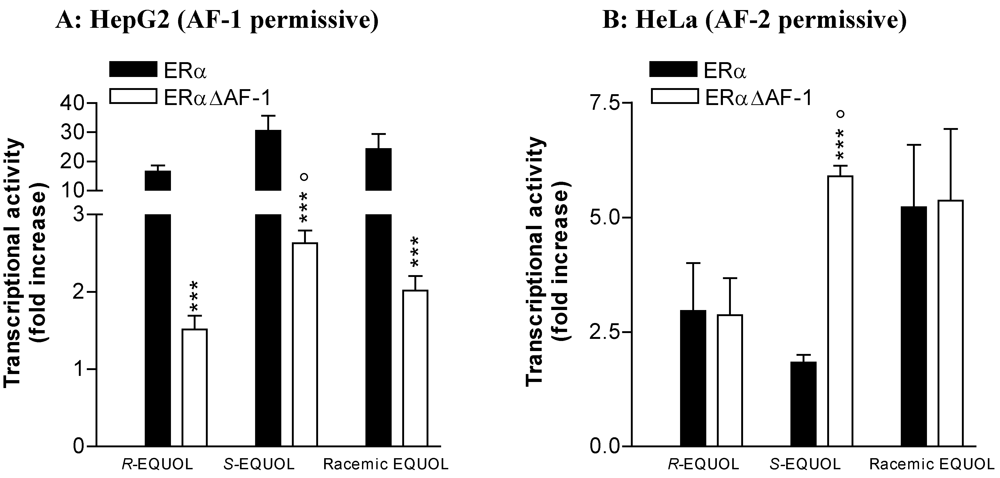
Comparative Effects of R- and S-equol and Implication of Transactivation Functions (AF-1 and AF-2) in Estrogen Receptor-Induced Transcriptional Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Shinkaruk, S.; Carreau, C.; Flouriot, G.; Bennetau-Pelissero, C.; Potier, M. Comparative Effects of R- and S-equol and Implication of Transactivation Functions (AF-1 and AF-2) in Estrogen Receptor-Induced Transcriptional Activity. Nutrients 2010, 2, 340-354. https://doi.org/10.3390/nu2030340
Shinkaruk S, Carreau C, Flouriot G, Bennetau-Pelissero C, Potier M. Comparative Effects of R- and S-equol and Implication of Transactivation Functions (AF-1 and AF-2) in Estrogen Receptor-Induced Transcriptional Activity. Nutrients. 2010; 2(3):340-354. https://doi.org/10.3390/nu2030340
Chicago/Turabian StyleShinkaruk, Svitlana, Charlotte Carreau, Gilles Flouriot, Catherine Bennetau-Pelissero, and Mylène Potier. 2010. "Comparative Effects of R- and S-equol and Implication of Transactivation Functions (AF-1 and AF-2) in Estrogen Receptor-Induced Transcriptional Activity" Nutrients 2, no. 3: 340-354. https://doi.org/10.3390/nu2030340
APA StyleShinkaruk, S., Carreau, C., Flouriot, G., Bennetau-Pelissero, C., & Potier, M. (2010). Comparative Effects of R- and S-equol and Implication of Transactivation Functions (AF-1 and AF-2) in Estrogen Receptor-Induced Transcriptional Activity. Nutrients, 2(3), 340-354. https://doi.org/10.3390/nu2030340