Long-Chain Omega-3 Polyunsaturated Fatty Acids May Be Beneficial for Reducing Obesity—A Review



{kind=link}
Abstract
:1. Introduction
2. LC n-3 PUFA and Body Fat Reduction in Animals
2.1. Protection against Body Fat Gain
2.2. LC n-3 PUFA for Reducing Existing Obesity
3. LC n-3 PUFA and Body Fat Reduction in Humans
3.1. Observational Studies—Relationships between LC n-3 PUFA Intake and Body Weight
3.2. Randomized Controlled Trials
3.2.1. Effect of LC n-3 PUFA Intake Alone on Body Composition
3.2.2. Effect of LC n-3 PUFA Intake Combined with Dietary Energy Restriction on Body Composition
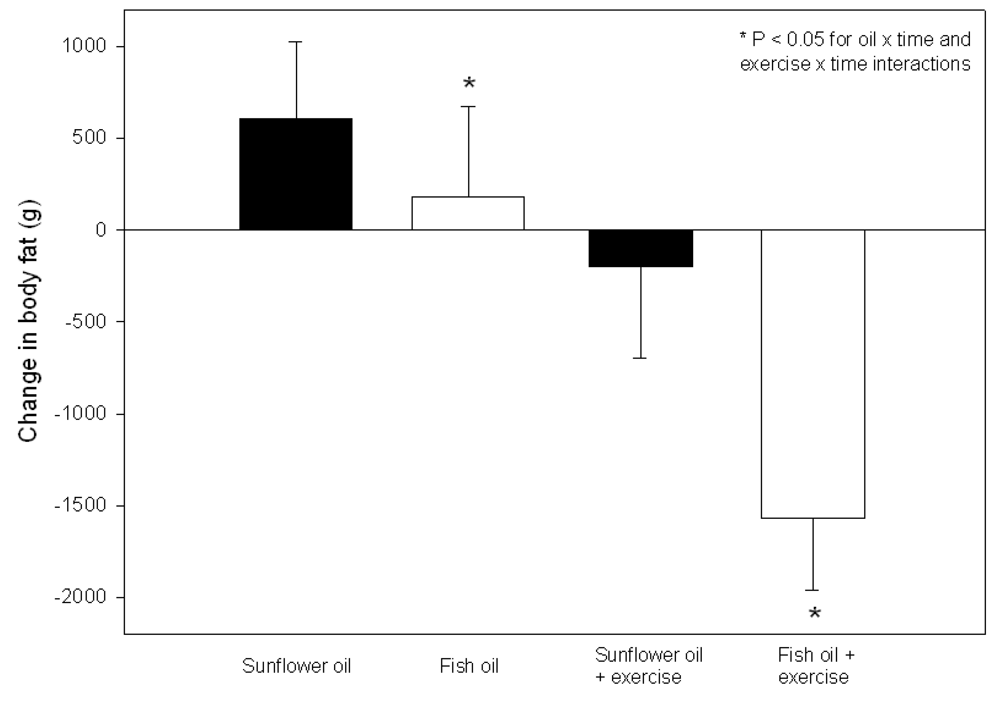
3.2.3. Effect of LC n-3 PUFA Intake Combined with Regular Exercise on Body Composition
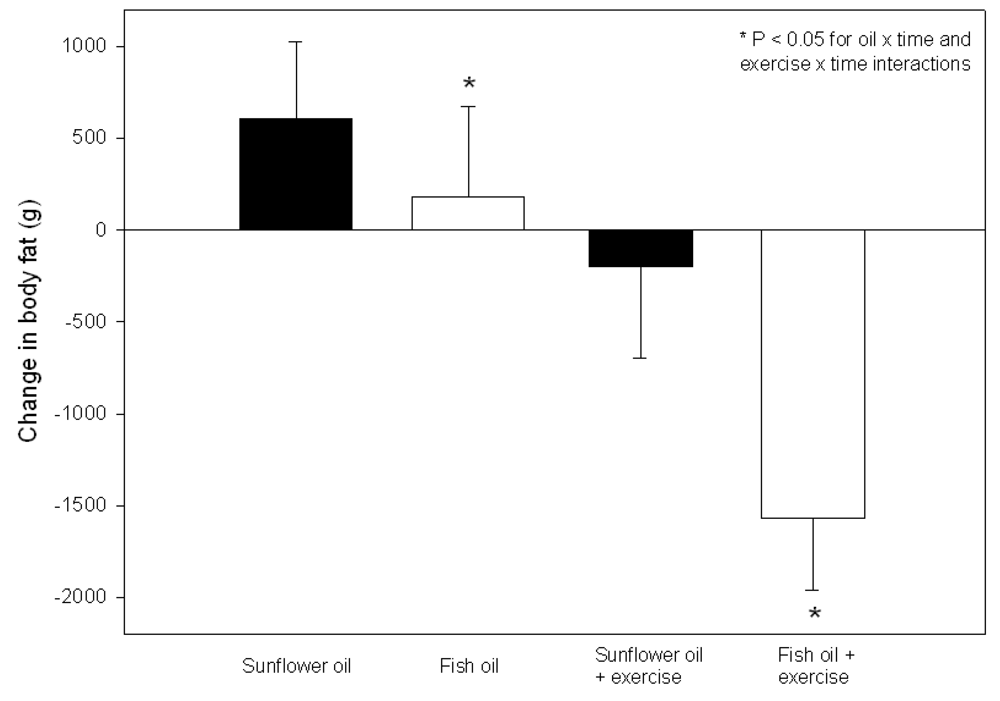
3.2.4. Effect of LC n-3 PUFA Intake Combined with Dietary Energy Restriction and Regular Exercise on Body Composition
3.3. Summary of Effects of LC n-3 PUFA Intake on Body Composition in Humans
4. Mechanisms of Body Fat Reduction with LC n-3 PUFA
4.1. Appetite Effects
4.2. Effects on Expression of Genes Regulating Metabolic Pathways
4.3. Effects on Tissue Anabolism and Maintenance of Lean Tissue Mass
4.4. Effects on Muscle Blood Flow
5. Conclusions
References
- Dunstan, D.W.; Zimmet, P.Z.; Welborn, T.A.; De Courten, M.P.; Cameron, A.J.; Sicree, R.A.; Dwyer, T.; Colagiuri, S.; Jolley, D.; Knuiman, M.; Atkins, R.; Shaw, J.E. The rising prevalence of diabetes and impaired glucose tolerance: The Australian Diabetes, Obesity and Lifestyle Study. Diabetes Care 2002, 25, 829–834. [Google Scholar]
- Flegel, K.; Carroll, M.; Kuczmarski, R.; Johnson, C. Overweight and obesity in the United States: Prevalence and trends, 1960–1994. Int. J. Obes. 1998, 22, 39–47. [Google Scholar]
- Must, A.; Spadano, J.; Coakley, E.H.; Field, A.E.; Colditz, G.; Dietz, W.H. The disease burden associated with overweight and obesity. JAMA 1999, 282, 1523–1529. [Google Scholar]
- Visscher, T.L.; Seidell, J.C. The public health impact of obesity. Annu. Rev. Public Health 2001, 22, 355–375. [Google Scholar]
- Department of Health and Ageing, National Physical Activity Guidelines for Adults; Commonwealth of Australia: Canberra, Australia, 2005.
- National Health and Medical Research Council, Dietary Guidelines for Australian Adults; Commonwealth of Australia: Canberra, Australia, 2003.
- Curioni, C.C.; Lourenco, P.M. Long-term weight loss after diet and exercise: A systematic review. Int. J. Obes. 2005, 29, 1168–1174. [Google Scholar]
- Buckley, J.; Howe, P. Anti-obesity effects of omega-3 long-chain polyunsaturated fatty acids. Obes. Rev. 2009, 10, 648–659. [Google Scholar]
- Baillie, R.; Takada, R.; Nakamura, M.; Clarke, S. Coordinate induction of peroxisomal acyl-CoA oxidase and UCP-3 by dietary fishoil: A mechanism for decreased body fat deposition. Prostaglandins Leukot. Essent. Fatty Acids 1999, 60, 351–356. [Google Scholar]
- Belzung, F.; Raclot, T.; Groscolas, R. Fish oil n-3 fatty acids selectively limit the hypertrophy of abdominal fat depots in growing rats fed high-fat diets. Am. J. Physiol. 1993, 264, R1111–R1118. [Google Scholar]
- Cunnane, S.; McAdoo, K.; Horrobin, D. n-3 essential fatty acids decrease weight gain in genetically obese mice. Br. J. Nutr. 1986, 56, 87–95. [Google Scholar]
- Hainault, I.; Carlotti, M.; Hajduch, E.; Guichard, C.; Lavau, M. Fish oil in a high lard diet prevents obesity, hyperlipidemia, and adipocyte insulin resistance in rats. Ann. N. Y. Acad. Sci. 1993, 683, 98–101. [Google Scholar]
- Ruzickova, J.; Rossmeisl, M.; Prazak, T.; Flachs, P.; Sponarova, J.; Veck, M.; Tvrzicka, E.; Bryhn, M.; Kopecky, J. Omega-3 PUFA of marine origin limit diet-induced obesity in mice by reducing cellularity of adipose tissue. Lipids 2004, 39, 1177–1185. [Google Scholar]
- Todoric, J.; Löffler, M.; Huber, J.; Bilban, M.; Reimers, M.; Kadl, A.; Zeyda, M.; Waldhäusl, W.; Stulnig, T. Adipose tissue inflammation induced by high-fat diet in obese diabetic mice is prevented by n-3 polyunsaturated fatty acids. Diabetologia 2006, 49, 2109–2119. [Google Scholar]
- Clarke, S. Polyunsaturated fatty acid regulation of gene transcription: A mechanism to improve energy balance and insulin resistance. Br. J. Nutr. 2000, 83, S59–S66. [Google Scholar]
- Parrish, C.; Pathy, D.; Parkes, J.; Angel, A. Dietary fish oils limit adipose tissue hypertrophy in rats. Metabolism 1990, 39, 217–219. [Google Scholar]
- Huang, X.F.; Xin, X.; McLennan, P.; Storlien, L. Role of fat amount and type in ameliorating diet-induced obesity: Insights at the level of hypothalamic arcuate nucleus leptin receptor, neuropeptide Y and pro-opiomelanocortin mRNA expression. Diabetes Obes. Metab. 2004, 6, 35–44. [Google Scholar]
- He, K.; Rimm, E.; Merchant, A.; Rosner, B.; Stampfer, M.; Willett, W.; Ascherio, A. Fish consumption and risk of stroke in men. JAMA 2002, 288, 3130–3136. [Google Scholar]
- Iso, H.; Rexrode, K.M.; Stampfer, M.J.; Manson, J.E.; Colditz, G.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Intake of fish and omega-3 fatty acids and risk of stroke in women. JAMA 2001, 285, 304–312. [Google Scholar]
- Garaulet, M.; Pérez-Llamas, F.; Pérez-Ayala, M.; Martínez, P.; de Medina, F.; Tebar, F.; Zamora, S. Site-specific differences in the fatty acid composition of abdominal adipose tissue in an obese population from a Mediterranean area: Relation with dietary fatty acids, plasma lipid profile, serum insulin, and central obesity. Am. J. Clin. Nutr. 2001, 74, 585–591. [Google Scholar] [PubMed]
- Garaulet, M.; Hernandez-Morante, J.; Lujan, J.; Tebar, F.; Zamora, S. Relationship between fat cell size and number and fatty acid composition in adipose tissue from different fat depots in overweight/obese humans. Int. J. Obes. 2006, 30, 899–905. [Google Scholar]
- Micaleff, M.; Munro, I.; Phang, M.; Garg, M. Plasma n-3 polyunsaturated fatty acids are negatively associated with obesity. Br. J. Nutr. 2009, 102, 1370–1374. [Google Scholar]
- Couet, C.; Delarue, J.; Ritz, P.; Antoine, J.-M.; Lamisse, F. Effect of dietary fish oil on body fat mass and basal fat oxidation in healthy adults. Int. J. Obes. 1997, 21, 637–643. [Google Scholar]
- Kabir, M.; Skurnik, G.; Naour, N.; Pechtner, V.; Meugnier, E.; Rome, S.; Quignard-Boulange, A.; Vidal, H.; Slama, G.; Clement, K.; Guerre-Millo, M.; Rizkalla, S. Treatment for 2 mo with n-3 polyunsaturated fatty acids reduces adiposity and some atherogenic factors but does not improve insulin sensitivity in women with type 2 diabetes: A randomized controlled study. Am. J. Clin. Nutr. 2007, 86, 1670–1679. [Google Scholar]
- Fontani, G.; Corradeschi, F.; Felici, A.; Alfatti, F.; Bugarini, R.; Fiaschi, A.I.; Cerretani, D.; Montorfano, G.; Rizzo, A.M.; Berra, B. Blood profiles, body fat and mood state in healthy subjects on different diets supplemented with Omega-3 polyunsaturated fatty acids. Eur. J. Clin. Invest. 2005, 35, 499–507. [Google Scholar]
- Buckley, J.; Burgess, S.; Murphy, K.; Howe, P. DHA-rich fish oil lowers heart rate during submaximal exercise in elite Australian Rules footballers. J. Sci. Med. Sport 2009, 12, 503–507. [Google Scholar]
- Brown, A.; Pang, E.; Roberts, D. Persistent changes in the fatty acid composition of erythrocyte membranes after moderate intake of n-3 polyunsaturated fatty acids: Study design implications. Am. J. Clin. Nutr. 1991, 54, 668–673. [Google Scholar]
- Krebs, J.; Browning, L.; McLean, N.; Rothwell, J.; Mishra, G.; Moore, C.; Jebb, S. Additive benefits of long-chain n-3 polyunsaturated fatty acids and weight-loss in the management of cardiovascular disease risk in overweight hyperinsulinaemic women. Int. J. Obes. 2006, 30, 1535–1544. [Google Scholar]
- Thorsdottir, I.; Tomasson, H.; Gunnarsdottir, I.; Gisladottir, E.; Kiely, M.; Parra, M.; Bandarra, N.; Schaafsma, G.; Martine, J. Randomized trial of weight-loss-diets for young adults varying in fish and fish oil content. Int. J. Obes. 2007, 31, 1560–1566. [Google Scholar] [Green Version]
- Fujihira, E.; Takahashi, H.; Nakazawa, M. Effect of long-term feeding of taurine in hereditary hyperglycemic obese mice. Chem. Pharm. Bull. (Tokyo) 1970, 18, 1636–1642. [Google Scholar] [PubMed]
- Warner, J.; Ullrich, I.; Albrink, M.; Yeater, R. Combined effects of aerobic exercise and omega-3 fatty acids in hyperlipidemic persons. Med. Sci. Sports Exerc. 1989, 21, 498–505. [Google Scholar]
- Brilla, L.; Landerholm, T. Effect of fish oil supplementation and exercise on serum lipids and aerobic fitness. J. Sports Med. Phys. Fitness 1990, 30, 173–180. [Google Scholar]
- Hill, A.; Buckley, J.; Murphy, K.; Howe, P. Combining fish oil supplementation with regular aerobic exercise improves body composition and cardiovascular risk factors. Am. J. Clin. Nutr. 2007, 85, 1267–1274. [Google Scholar]
- Kunešová, M.; Braunerová, R.; Hlavatý, P.; Tvrzická, E.; Staňková, B.; Škrha, J.; Hilgertová, J.; Hill, M.; Kopecký, J.; Wagenchnecht, M.; Hainer, V.; Matoulek, M.; Pařízková, J.; Žák, A.; Svačina, Š. The influence of n-3 polyunsaturated fatty acids and very low calorie diet during a short-term weight reducing regimen on weight loss and serum fatty acid composition in severely obese women. Physiol. Res. 2006, 55, 63–72. [Google Scholar] [PubMed]
- Parra, D.; Ramel, A.; Bandarra, N.; Kiely, M.; Martinez, J.; Thorsdottir, I. A diet rich in long chain omega 3 fatty acids modulates satiety in overweight and obese volunteers during weight loss. Appetite 2008, 51, 676–680. [Google Scholar]
- Mascaro, C.; Acosta, E.; Ortiz, J.; Marrero, P.; Hegardt, F.; Haro, D. Control of human muscle–type carnitine palmitoyltransferase I gene transcription by peroxisome proliferator–activated receptor. J. Biol. Chem. 1998, 273, 8560–8563. [Google Scholar]
- Laurente-Cebrián, S.; Bustos, M.; Marti, A.; Martinez, J.; Moreno-Aliaga, M. Eicosapentaenoic acid stimulates AMP-activated protein kinase and increases visfatin secretion in cultured murine adipocytes. Clin. Sci. 2009, 117, 243–249. [Google Scholar]
- Motawi, T.; Hashem, R.; Rashed, L.; El-Razek, S. Comparative study between the effect of the peroxisome proliferator activated receptor-alpha ligands fenofibrate and n-3 polyunsaturated fatty acids on activation of 5'-AMP-activated protein kinase-alpha1 in high-fat fed rats. J. Pharm. Pharmacol. 2009, 61, 1339–1346. [Google Scholar]
- Power, G.; Newsholme, E. Dietary fatty acids influence the activity and metabolic control of mitochondrial carnitine palmitoyltransferase I in rat heart and skeletal muscle. J. Nutr. 1997, 127, 2142–2150. [Google Scholar]
- Elayan, I.; Winder, W. Effect of glucose infusion on muscle malonyl-CoA during exercise. J. Appl. Physiol. 1991, 70, 1495–1499. [Google Scholar]
- Reddy, J.; Mannaerts, G. Peroxisomal lipid metabolism. Annu. Rev. Nutr. 1994, 14, 343–370. [Google Scholar]
- Mori, T.; Kondo, H.; Hase, T.; Tokimitsu, I.; Murase, T. Dietary Fish Oil Upregulates Intestinal Lipid Metabolism and Reduces Body Weight Gain in C57BL/6J Mice. J. Nutr. 2007, 137, 2629–2634. [Google Scholar]
- Dagenais, G.; Tancredi, R.; Zierler, K. Free fatty acid oxidation by forearm muscle at rest, and evidence for an intramuscular lipid pool in the human forearm. J. Clin. Invest. 1976, 58, 421–431. [Google Scholar]
- Flachs, P.; Horakova, O.; Brauner, P.; Rossmeisl, M.; Pecina, P.; Franssen-van Hal, N.; Ruzickova, J.; Sponarova, J.; Drahota, Z.; Vlcek, C.; Keijer, J.; Houstek, J.; Kopecky, J. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce β–oxidation in white fat. Diabetologia 2005, 48, 2365–2375. [Google Scholar]
- Kelly, D.P.; Scarpulla, R.C. Transcriptional regulatory circuits controlling mitochondrial biogenesis and function. Genes Dev. 2004, 18, 357–368. [Google Scholar]
- Vanden Heuvel, J.P. Fatty Acid Transcriptomics. In Proceedings of the 8th Meeting of the International Society for the Study of Fatty Acids and Lipids, Kansas City, MO, USA, 17–22 May 2008; p. 115.
- Whitehouse, A.; Tisdale, M. Downregulation of ubiquitin-dependent proteolysis by eicosapentaenoic acid in acute starvation. Biochem. Biophys. Res. Commun. 2001, 285, 598–602. [Google Scholar]
- Wing, S.; Goldberg, A. Glucocorticoids activate the ATP-ubiquitin-dependent proteolytic system in skeletal muscle during fasting. Am. J. Physiol. 1993, 264, E668–E676. [Google Scholar]
- Wyke, S.; Tisdale, M. NF-kB mediates proteolysis-inducing factor induced protein degradation and expression of the ubiquitin-proteasome system in skeletal muscle. Br. J. Cancer 2005, 92, 711–721. [Google Scholar]
- Gingras, A.; White, P.; Chouinard, P.; Julien, P.; Davis, T.; Dombrowski, L.; Couture, Y.; Dubreuil, P.; Myre, A.; Bergeron, K.; Marette, A.; Thivierge, M. Long-chain omega-3 fatty acids regulate bovine whole-body protein metabolism by promoting muscle insulin signalling to the Akt-mTOR-S6K1 pathway and insulin sensitivity. J. Physiol. 2007, 579, 269–284. [Google Scholar]
- Hamdy, O.; Ledbury, S.; Mullooly, C.; Jarema, C.; Porter, S.; Ovalle, K.; Moussa, A.; Caselli, A.; Caballero, A.E.; Economides, P.A.; Veves, A.; Horton, E.S. Lifestyle modification improves endothelial function in obese subjects with the insulin resistance syndrome. Diabetes Care 2003, 26, 2119–2125. [Google Scholar]
- Parikh, N.; Keyes, M.; Larson, M.; Pou, K.; Hamburg, N.; VIta, J.; O'Donnell, C.; Vasan, R.; Mitchell, G.; Hoffman, U.; Fox, C.; Benjamin, E. Visceral and subcutaneous adiposity and brachial artery vasodilator function. Obesity 2009, 17, 2054–2059. [Google Scholar]
- Hodnett, B.; Hester, R. Regulation of muscle blood flow in obesity. Microcirculation 2007, 14, 273–288. [Google Scholar]
- Clerk, L.; Vincent, M.; Jahn, L.; Liu, Z.; Lindner, J.; Barrett, E. Obesity blunts insulin-mediated microvascular recruitment in human forearm muscle. Diabetes 2006, 55, 1436–1442. [Google Scholar]
- Walser, B.; Giordano, R.; Stebbins, C. Supplementation with omega-3 polyunsaturated fatty acids augments brachial artery dilation and blood flow during forearm contraction. Eur. J. Appl. Physiol. 2006, 97, 347–354. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Buckley, J.D.; Howe, P.R.C. Long-Chain Omega-3 Polyunsaturated Fatty Acids May Be Beneficial for Reducing Obesity—A Review. Nutrients 2010, 2, 1212-1230. https://doi.org/10.3390/nu2121212
Buckley JD, Howe PRC. Long-Chain Omega-3 Polyunsaturated Fatty Acids May Be Beneficial for Reducing Obesity—A Review. Nutrients. 2010; 2(12):1212-1230. https://doi.org/10.3390/nu2121212
Chicago/Turabian StyleBuckley, Jonathan D., and Peter R. C. Howe. 2010. "Long-Chain Omega-3 Polyunsaturated Fatty Acids May Be Beneficial for Reducing Obesity—A Review" Nutrients 2, no. 12: 1212-1230. https://doi.org/10.3390/nu2121212