
Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
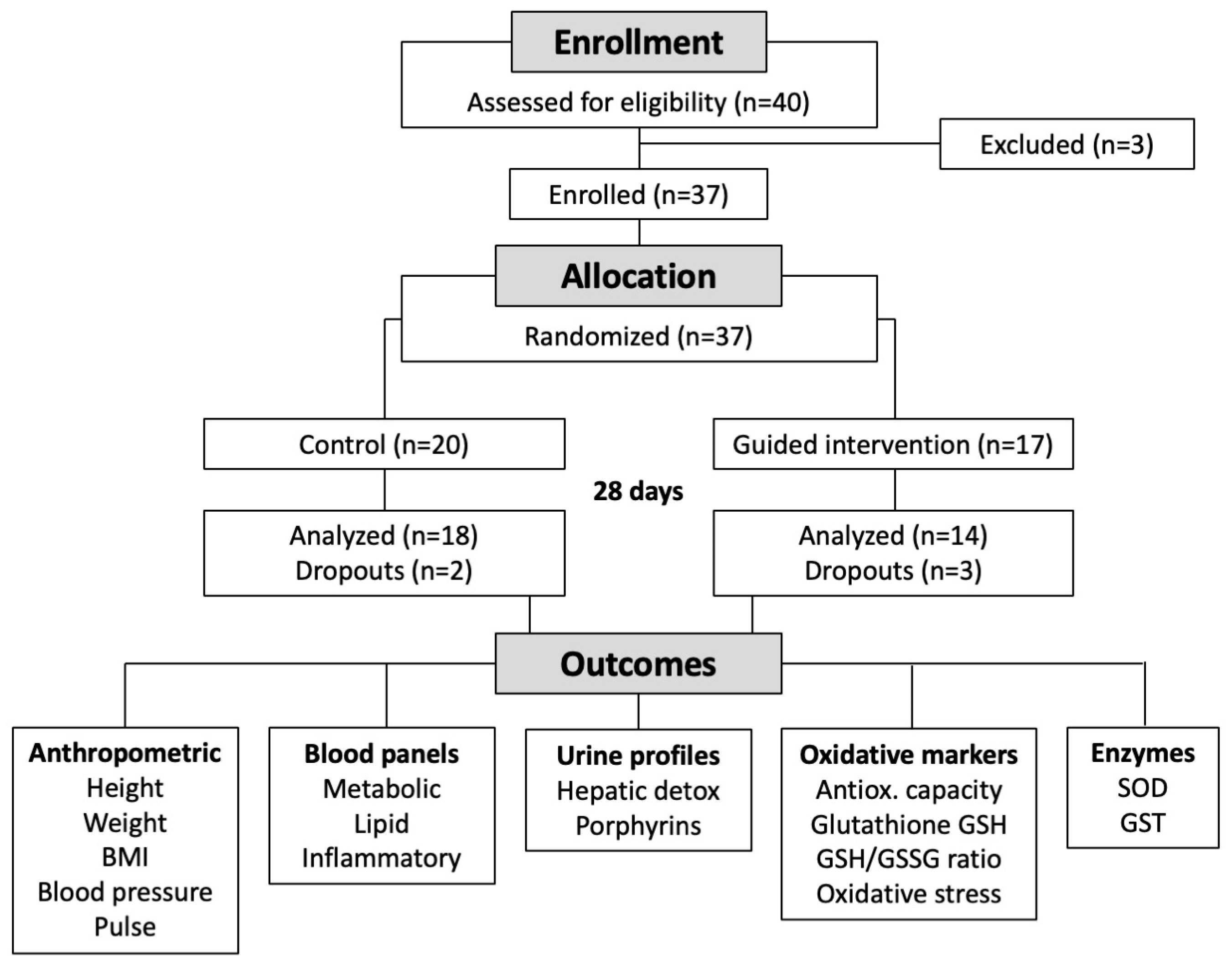
2.1. Participants
2.2. Study Investigational Product
2.3. Anthropometrics
2.4. Laboratory Testing
2.5. Antioxidant and Redox Status
2.6. Oxidative Stress
2.7. Detoxification Enzyme Activity
2.8. Self-Reported Wellness
2.9. Statistics
3. Results
3.1. Subjects’ Characteristics
3.2. Safety and Tolerance Assessment
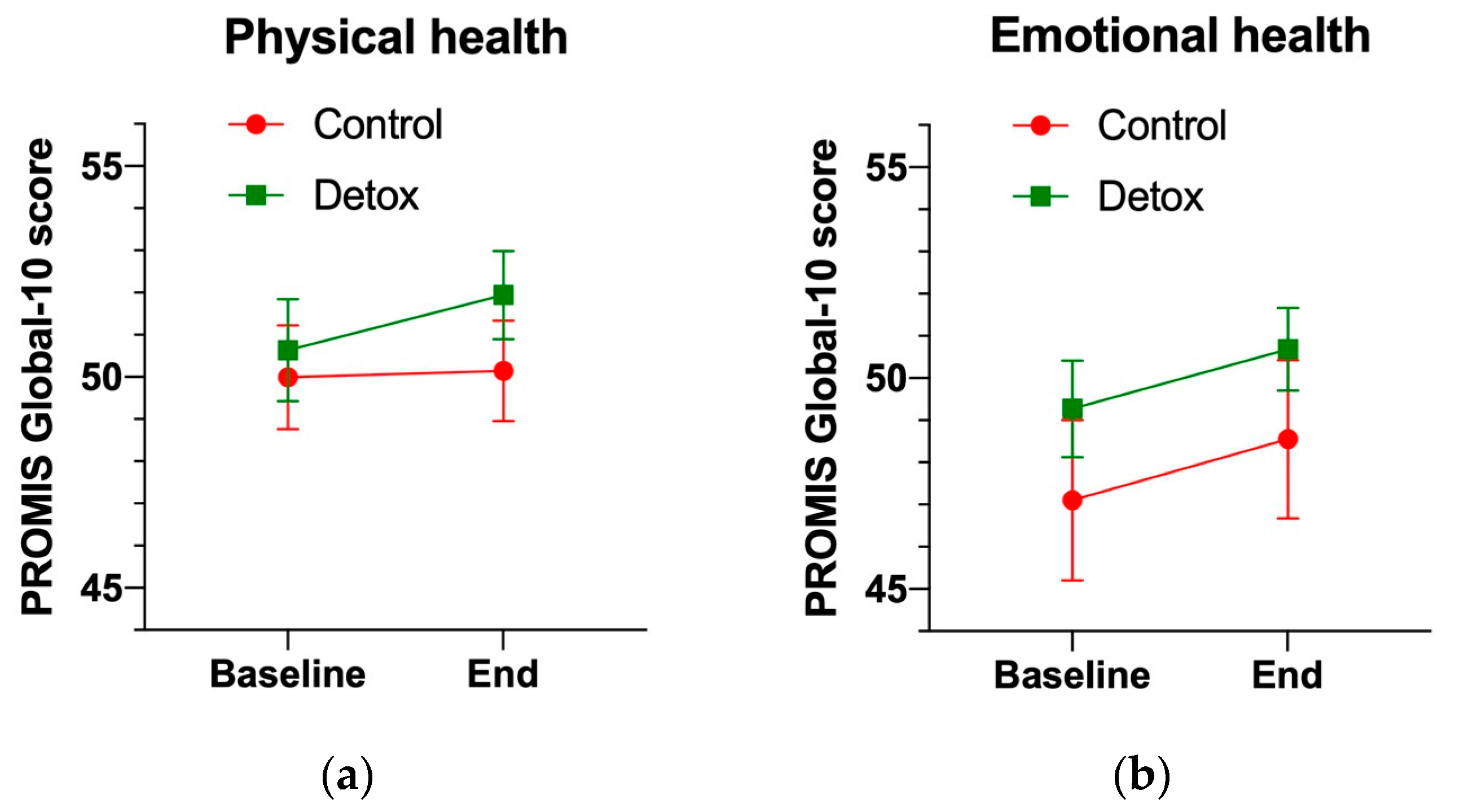
3.3. Wellness Status and Quality of Life
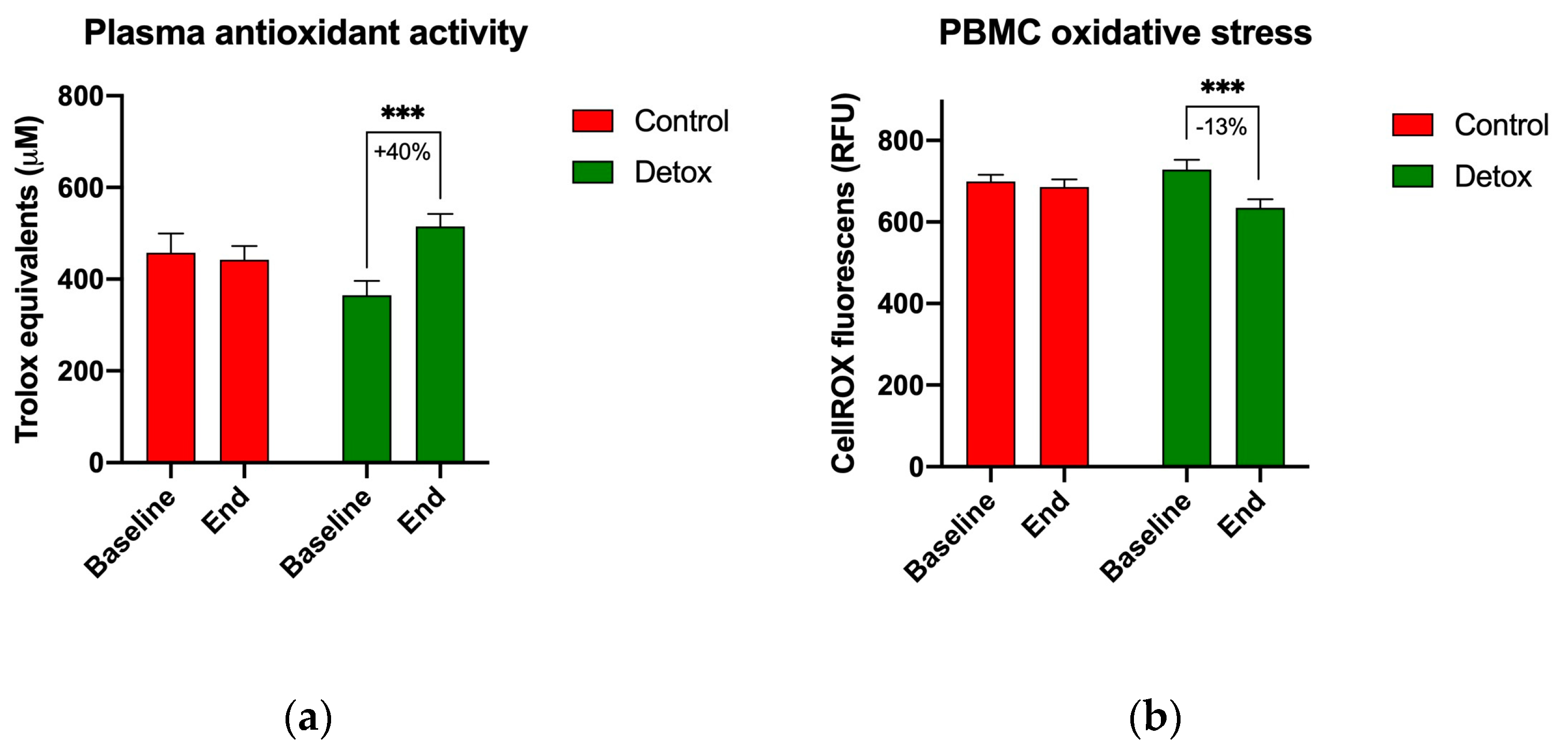
3.4. Biomarkers of Oxidative Stress
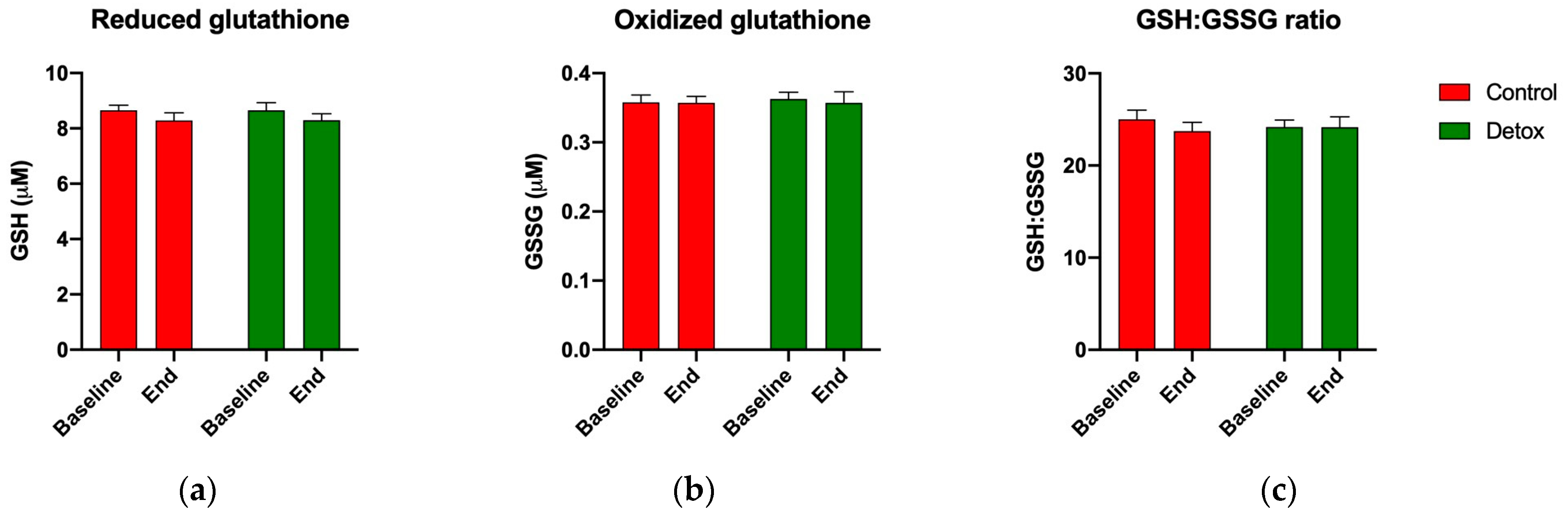
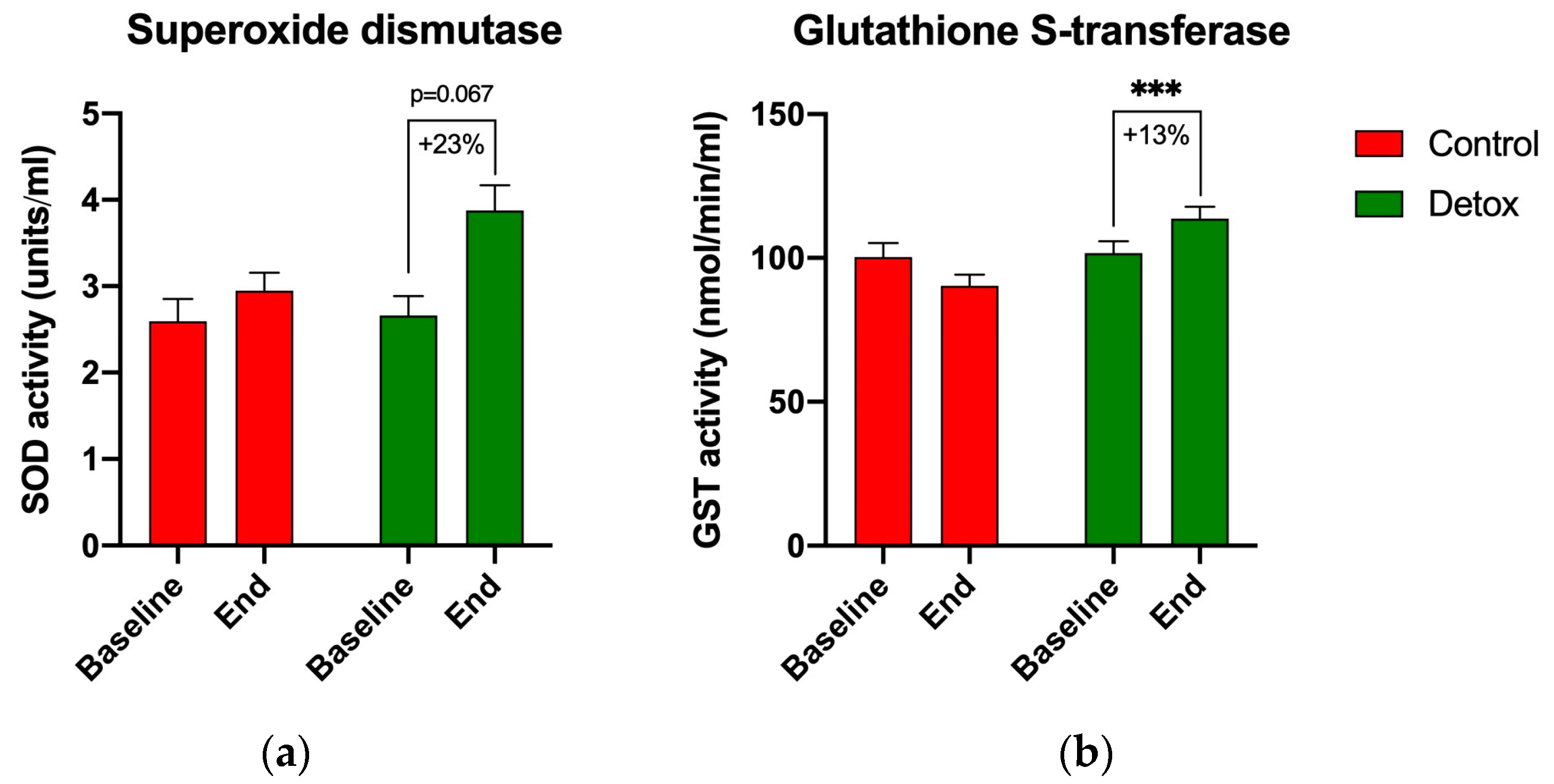
3.5. Redox Homeostasis
3.6. Detoxification Markers in Urine
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sears, M.E.; Genuis, S.J. Environmental Determinants of Chronic Disease and Medical Approaches: Recognition, Avoidance, Supportive Therapy, and Detoxification. J. Environ. Public Health 2012, 2012, 356798. [Google Scholar] [CrossRef]
- Guo, W.; Pan, B.; Sakkiah, S.; Yavas, G.; Ge, W.; Zou, W.; Tong, W.; Hong, H. Persistent Organic Pollutants in Food: Contamination Sources, Health Effects and Detection Methods. Int. J. Environ. Res. Public Health 2019, 16, 4361. [Google Scholar] [CrossRef]
- Komarnytsky, S.; Retchin, S.; Vong, C.I.; Lila, M.A. Gains and Losses of Agricultural Food Production: Implications for the Twenty-First Century. Annu. Rev. Food. Sci. Technol. 2022, 13, 239–261. [Google Scholar] [CrossRef]
- Kelishadi, R.; Mirghaffari, N.; Poursafa, P.; Gidding, S.S. Lifestyle and Environmental Factors Associated with Inflammation, Oxidative Stress and Insulin Resistance in Children. Atherosclerosis 2009, 203, 311–319. [Google Scholar] [CrossRef]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic Transgenerational Actions of Environmental Factors in Disease Etiology. Trends Endocrinol. Metab. 2010, 21, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Marroqui, L.; Tudurí, E.; Alonso-Magdalena, P.; Quesada, I.; Nadal, Á.; Santos, R.S. dos Mitochondria as Target of Endocrine-Disrupting Chemicals: Implications for Type 2 Diabetes. J. Endocrinol. 2018, 239, R27–R45. [Google Scholar] [CrossRef] [PubMed]
- Komarnytsky, S.; Rathinasabapathy, T.; Wagner, C.; Metzger, B.; Carlisle, C.; Panda, C.; Le Brun-Blashka, S.; Troup, J.P.; Varadharaj, S. Endocannabinoid System and Its Regulation by Polyunsaturated Fatty Acids and Full Spectrum Hemp Oils. Int. J. Mol. Sci. 2021, 22, 5479. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; He, H.; Qiu, W.; Zheng, Y.; Chen, Y.; Hu, S.; Zhao, X. Oxidative Stress, Endocrine Disturbance, and Immune Interference in Humans Showed Relationships to Serum Bisphenol Concentrations in a Dense Industrial Area. Environ. Sci. Technol. 2021, 55, 1953–1963. [Google Scholar] [CrossRef]
- Jia, P.-P.; Junaid, M.; Xin, G.-Y.; Wang, Y.; Ma, Y.-B.; Pei, D.-S. Disruption of Intestinal Homeostasis Through Altered Responses of the Microbial Community, Energy Metabolites, and Immune System in Zebrafish After Chronic Exposure to DEHP. Front. Microbiol. 2021, 12, 729530. [Google Scholar] [CrossRef]
- Bonakdar, R.A.; Sweeney, M.; Dalhoumi, S.; Adair, V.; Garvey, C.; Hodge, T.; Herrala, L.; Barbee, A.; Case, C.; Kearney, J.; et al. Detoxification Enhanced Lifestyle Intervention Targeting Endotoxemia (DELITE) in the Setting of Obesity and Pain: Results of a Pilot Group Intervention. Integr. Med. 2020, 19, 16–28. [Google Scholar]
- Mostafalou, S.; Abdollahi, M. Pesticides and Human Chronic Diseases: Evidences, Mechanisms, and Perspectives. Toxicol. Appl. Pharmacol. 2013, 268, 157–177. [Google Scholar] [CrossRef]
- Liu, J.; Lewis, G. Environmental Toxicity and Poor Cognitive Outcomes in Children and Adults. J. Environ. Health 2014, 76, 130–138. [Google Scholar]
- Pizzorno, J. Environmental Toxins and Infertility. Integr. Med. 2018, 17, 8–11. [Google Scholar]
- Grant, D.M. Detoxification Pathways in the Liver. J. Inherit. Metab. Dis. 1991, 14, 421–430. [Google Scholar] [CrossRef]
- Zamek-Gliszczynski, M.J.; Hoffmaster, K.A.; Nezasa, K.; Tallman, M.N.; Brouwer, K.L.R. Integration of Hepatic Drug Transporters and Phase II Metabolizing Enzymes: Mechanisms of Hepatic Excretion of Sulfate, Glucuronide, and Glutathione Metabolites. Eur. J. Pharm. Sci. 2006, 27, 447–486. [Google Scholar] [CrossRef]
- Hodges, R.E.; Minich, D.M. Modulation of Metabolic Detoxification Pathways Using Foods and Food-Derived Components: A Scientific Review with Clinical Application. J. Nutr. Metab. 2015, 2015, 760689. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T.; Bhatnagar, V.; Poloyac, S.M.; Momper, J.D. The Systems Biology of Drug Metabolizing Enzymes and Transporters: Relevance to Quantitative Systems Pharmacology. Clin. Pharmacol. Ther. 2020, 108, 40–53. [Google Scholar] [CrossRef]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.-R.; Harrison, D.G.; Bhatnagar, A. Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef]
- Labarrere, C.A.; Kassab, G.S. Glutathione: A Samsonian Life-Sustaining Small Molecule That Protects against Oxidative Stress, Ageing and Damaging Inflammation. Front. Nutr. 2022, 9, 1007816. [Google Scholar] [CrossRef]
- Zitka, O.; Skalickova, S.; Gumulec, J.; Masarik, M.; Adam, V.; Hubalek, J.; Trnkova, L.; Kruseova, J.; Eckschlager, T.; Kizek, R. Redox Status Expressed as GSH: GSSG Ratio as a Marker for Oxidative Stress in Paediatric Tumour Patients. Oncol. Lett. 2012, 4, 1247–1253. [Google Scholar] [CrossRef]
- Kalinina, E.V.; Chernov, N.N.; Novichkova, M.D. Role of Glutathione, Glutathione Transferase, and Glutaredoxin in Regulation of Redox-Dependent Processes. Biochemistry 2014, 79, 1562–1583. [Google Scholar] [CrossRef]
- Minich, D.M.; Brown, B.I. A Review of Dietary (Phyto)Nutrients for Glutathione Support. Nutrients 2019, 11, 2073. [Google Scholar] [CrossRef]
- Lampe, J.W.; Chen, C.; Li, S.; Prunty, J.; Grate, M.T.; Meehan, D.E.; Barale, K.V.; Dightman, D.A.; Feng, Z.; Potter, J.D. Modulation of Human Glutathione S-Transferases by Botanically Defined Vegetable Diets. Cancer Epidemiol. Biomark. Prev. 2000, 9, 787–793. [Google Scholar]
- Bettermann, E.L.; Hartman, T.J.; Easley, K.A.; Ferranti, E.P.; Jones, D.P.; Quyyumi, A.A.; Vaccarino, V.; Ziegler, T.R.; Alvarez, J.A. Higher Mediterranean Diet Quality Scores and Lower Body Mass Index Are Associated with a Less-Oxidized Plasma Glutathione and Cysteine Redox Status in Adults. J. Nutr. 2018, 148, 245–253. [Google Scholar] [CrossRef]
- Tinsley, G.; Urbina, S.; Santos, E.; Villa, K.; Foster, C.; Wilborn, C.; Taylor, L. A Purported Detoxification Supplement Does Not Improve Body Composition, Waist Circumference, Blood Markers, or Gastrointestinal Symptoms in Healthy Adult Females. J. Diet. Suppl. 2019, 16, 649–658. [Google Scholar] [CrossRef]
- Navarro, S.L.; Chang, J.-L.; Peterson, S.; Chen, C.; King, I.B.; Schwarz, Y.; Li, S.S.; Li, L.; Potter, J.D.; Lampe, J.W. Modulation of Human Serum Glutathione S-Transferase-A1/2 Concentration by Cruciferous Vegetables in a Controlled Feeding Study Is Influenced by GSTM1 and GSTT1 Genotypes. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2974–2978. [Google Scholar] [CrossRef]
- Gasper, A.V.; Al-Janobi, A.; Smith, J.A.; Bacon, J.R.; Fortun, P.; Atherton, C.; Taylor, M.A.; Hawkey, C.J.; Barrett, D.A.; Mithen, R.F. Glutathione S-Transferase M1 Polymorphism and Metabolism of Sulforaphane from Standard and High-Glucosinolate Broccoli. Am. J. Clin. Nutr. 2005, 82, 1283–1291. [Google Scholar] [CrossRef]
- Evans, M.; Paterson, E.; Barnes, D.M. An Open Label Pilot Study to Evaluate the Efficacy of Spanish Black Radish on the Induction of Phase I and Phase II Enzymes in Healthy Male Subjects. BMC Complement. Altern. Med. 2014, 14, 475. [Google Scholar] [CrossRef]
- Kadam, P.; Bhalerao, S. Sample size calculation. Int. J. Ayurveda Res. 2010, 1, 55–57. [Google Scholar] [CrossRef]
- Hays, R.D.; Bjorner, J.B.; Revicki, D.A.; Spritzer, K.L.; Cella, D. Development of Physical and Mental Health Summary Scores from the Patient-Reported Outcomes Measurement Information System (PROMIS) Global Items. Qual. Life Res. 2009, 18, 873–880. [Google Scholar] [CrossRef]
- Mega, A.; Marzi, L.; Kob, M.; Piccin, A.; Floreani, A. Food and Nutrition in the Pathogenesis of Liver Damage. Nutrients 2021, 13, 1326. [Google Scholar] [CrossRef]
- Liska, D.J. The Detoxification Enzyme Systems. Altern. Med. Rev. 1998, 3, 187–198. [Google Scholar]
- Bailey, D.G.; Dresser, G.K. Natural Products and Adverse Drug Interactions. Can. Med. Assoc. J. 2004, 170, 1531–1532. [Google Scholar] [CrossRef]
- Forde, C.G.; Bolhuis, D. Interrelations Between Food Form, Texture, and Matrix Influence Energy Intake and Metabolic Responses. Curr. Nutr. Rep. 2022, 11, 124–132. [Google Scholar] [CrossRef]
- Capuano, E.; Oliviero, T.; Fogliano, V.; Pellegrini, N. Role of the Food Matrix and Digestion on Calculation of the Actual Energy Content of Food. Nutr. Rev. 2018, 76, 274–289. [Google Scholar] [CrossRef]
- Aguilera, J.M. The Food Matrix: Implications in Processing, Nutrition and Health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef]
- Anania, C.; Perla, F.M.; Olivero, F.; Pacifico, L.; Chiesa, C. Mediterranean Diet and Nonalcoholic Fatty Liver Disease. World J. Gastroenterol. 2018, 24, 2083–2094. [Google Scholar] [CrossRef]
- Guo, X.; Shao, X.; Li, J.; Li, S.; Li, K.; Li, D. Fruit and Vegetable Intake and Liver Cancer Risk: A Meta-Analysis of Prospective Cohort Studies. Food Funct. 2019, 10, 4478–4485. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schwedhelm, C.; Hoffmann, G.; Lampousi, A.-M.; Knüppel, S.; Iqbal, K.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food Groups and Risk of All-Cause Mortality: A Systematic Review and Meta-Analysis of Prospective Studies 1, 2. Am. J. Clin. Nutr. 2017, 105, 1462–1473. [Google Scholar] [CrossRef]
- Clifford, T.; Howatson, G.; West, D.J.; Stevenson, E.J. The Potential Benefits of Red Beetroot Supplementation in Health and Disease. Nutrients 2015, 7, 2801–2822. [Google Scholar] [CrossRef]
- El-Kott, A.F.; Bin-Meferij, M.M. Use of Arctium Lappa Extract Against Acetaminophen-Induced Hepatotoxicity in Rats. Curr. Ther. Res. Clin. Exp. 2015, 77, 73–78. [Google Scholar] [CrossRef]
- Lara-Espinoza, C.; Carvajal-Millán, E.; Balandrán-Quintana, R.; López-Franco, Y.; Rascón-Chu, A. Pectin and Pectin-Based Composite Materials: Beyond Food Texture. Molecules 2018, 23, 942. [Google Scholar] [CrossRef] [PubMed]
- Höferl, M.; Stoilova, I.; Schmidt, E.; Wanner, J.; Jirovetz, L.; Trifonova, D.; Krastev, L.; Krastanov, A. Chemical Composition and Antioxidant Properties of Juniper Berry (Juniperus communis L.) Essential Oil. Action of the Essential Oil on the Antioxidant Protection of Saccharomyces cerevisiae Model Organism. Antioxidants 2014, 3, 81–98. [Google Scholar] [CrossRef]
- Pfingstgraf, I.O.; Taulescu, M.; Pop, R.M.; Orăsan, R.; Vlase, L.; Uifalean, A.; Todea, D.; Alexescu, T.; Toma, C.; Pârvu, A.E. Protective Effects of Taraxacum officinale L. (Dandelion) Root Extract in Experimental Acute on Chronic Liver Failure. Antioxidants 2021, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Leoncini, E.; Malaguti, M.; Angelini, S.; Hrelia, P.; Hrelia, S. Modulation of Phase II Enzymes by Sulforaphane: Implications for Its Cardioprotective Potential. J. Agric. Food Chem. 2009, 57, 5615–5622. [Google Scholar] [CrossRef]
- Rangboo, V.; Noroozi, M.; Zavoshy, R.; Rezadoost, S.A.; Mohammadpoorasl, A. The Effect of Artichoke Leaf Extract on Alanine Aminotransferase and Aspartate Aminotransferase in the Patients with Nonalcoholic Steatohepatitis. Int. J. Hepatol. 2016, 2016, 4030476. [Google Scholar] [CrossRef]
- Bijak, M. Silybin, a Major Bioactive Component of Milk Thistle (Silybum marianum L. Gaernt.)-Chemistry, Bioavailability, and Metabolism. Molecules 2017, 22, 1942. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Medicinal Fungus Cordyceps with Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef] [PubMed]
- Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi, S.; Tieu, K.; Hammond, C.L. Glutathione Dysregulation and the Etiology and Progression of Human Diseases. Biol. Chem. 2009, 390, 191–214. [Google Scholar] [CrossRef]
- Pizzorno, J. Glutathione! Integr. Med. 2014, 13, 8–12. [Google Scholar]
- Guerrero-Beltrán, C.E.; Calderón-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective Effect of Sulforaphane against Oxidative Stress: Recent Advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.; Schwarz, Y.; Li, S.S.; Li, L.; King, I.B.; Chen, C.; Eaton, D.L.; Potter, J.D.; Lampe, J.W. CYP1A2, GSTM1, and GSTT1 Polymorphisms and Diet Effects on CYP1A2 Activity in a Crossover Feeding Trial. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3118–3125. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
|
|
| Control (n = 18) | Detox (n = 14) | |||
|---|---|---|---|---|
| Baseline | End of Study | Baseline | End of Study | |
| Age (years) | 33.6 ± 3.2 | 36.9 ± 3.2 | ||
| Gender (male/female, %) | 5/13 (27.8%) | 2/12 (14.3%) | ||
| Height, cm | 167.75 ± 3.02 | 167.75 ± 3.02 | 163.78 ± 2.90 | 163.79 ± 2.90 |
| Weight, kg | 80.69 ± 4.66 | 80.33 ± 4.70 ** (p = 0.002) | 70.59 ± 5.29 | 70.52 ± 5.33 |
| BMI, kg/m2 | 27.04 ± 1.52 | 26.58 ± 1.48 *** (p = 0.001) | 26.72 ± 1.66 | 26.63 ± 1.64 |
| Systolic BP, mmHg | 123.94 ± 3.03 | 120.33 ± 3.90 | 117.21 ± 2.43 | 118.14 ± 5.15 |
| Diastolic BP, mmHg | 79.67 ± 2.65 | 78.72 ± 3.13 | 74.07 ± 1.45 | 71.93 ± 1.73 |
| Pulse, bpm | 69.75 ± 2.77 | 69.75 ± 2.70 | 66.43 ± 2.17 | 66.00 ± 1.55 |
| Reference Range | Control | Detox | |||||
|---|---|---|---|---|---|---|---|
| Baseline | End of Study | p | Baseline | End of Study | p | ||
| D-glucaric acid, phase I (nM/mg) | 40–400 | 104.67 ± 12.62 | 122.89 ± 20.23 | 0.35 | 86.14 ± 8.03 | 106.07 ± 3.82 | 0.09 |
| Mercapturic acids, phase II (μM/mM) | 40–95 | 48.56 ± 2.34 | 48.61 ± 2.34 | 0.98 | 55.14 ± 3.82 | 51.07 ± 3.28 | 0.38 |
| Creatinine, mg/dL | 40–325 | 101.00 ± 10.12 | 119.33 ± 17.80 | 0.33 | 103.50 ± 11.48 | 91.50 ± 9.30 | 0.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, C.; Komarnytsky, S.; Fleming, M.N.; Marsh, C.; Barron, K.; Le Brun-Blashka, S.; Metzger, B. Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants. Nutrients 2023, 15, 2209. https://doi.org/10.3390/nu15092209
Panda C, Komarnytsky S, Fleming MN, Marsh C, Barron K, Le Brun-Blashka S, Metzger B. Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants. Nutrients. 2023; 15(9):2209. https://doi.org/10.3390/nu15092209
Chicago/Turabian StylePanda, Chinmayee, Slavko Komarnytsky, Michelle Norton Fleming, Carissa Marsh, Keri Barron, Sara Le Brun-Blashka, and Brandon Metzger. 2023. "Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants" Nutrients 15, no. 9: 2209. https://doi.org/10.3390/nu15092209
APA StylePanda, C., Komarnytsky, S., Fleming, M. N., Marsh, C., Barron, K., Le Brun-Blashka, S., & Metzger, B. (2023). Guided Metabolic Detoxification Program Supports Phase II Detoxification Enzymes and Antioxidant Balance in Healthy Participants. Nutrients, 15(9), 2209. https://doi.org/10.3390/nu15092209