
Microencapsulated Limosilactobacillus reuteri Encoding Lactoferricin-Lactoferrampin Targeted Intestine against Salmonella typhimurium Infection
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Biological Characterization of LR-LFCA
2.3. LR-LFCA Antibacterial Activity Assays In Vitro
2.4. Preparation of Microencapsulated LR-LFCA
2.5. Artificial Gastrointestinal Fluid and Bile Salt Resistance of Microencapsulated LR-LFCA
2.6. Animals and Experimental Design
2.7. Sample Collection and Organ Morphology Analysis
2.8. Determination of the Immune Organ Index, Cytokines Levels and Intestinal Antioxidant Parameters
2.9. Microbiome Analysis
2.10. Statistical Analyses
3. Results
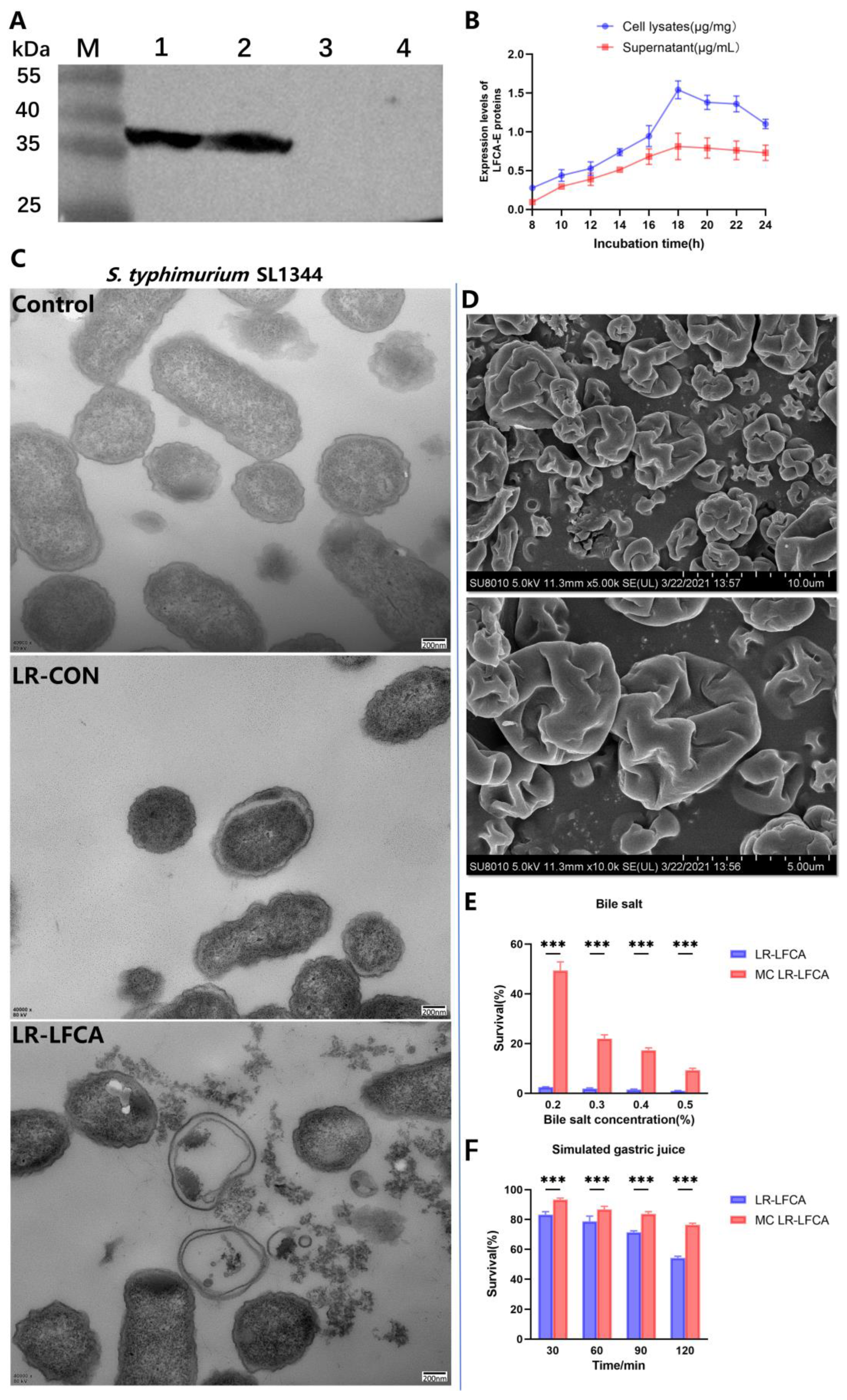
3.1. Bioactivity of LR-LFCA and Stress Resistance of Microencapsulated LR-LFCA
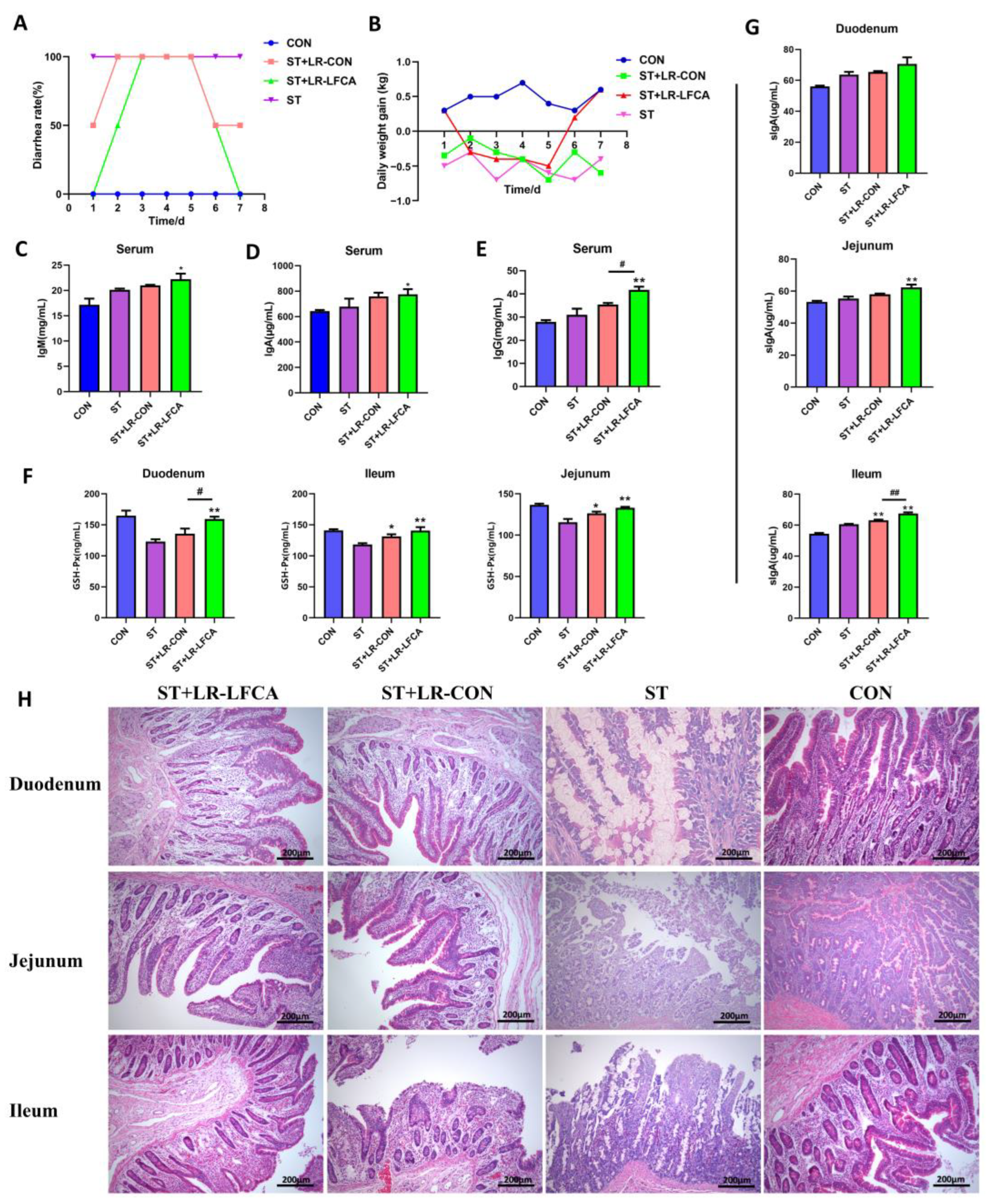
3.2. Oral Administration of Microencapsulated LR-LFCA to Weaned Piglets Protected agsainst S. typhimurium Infection
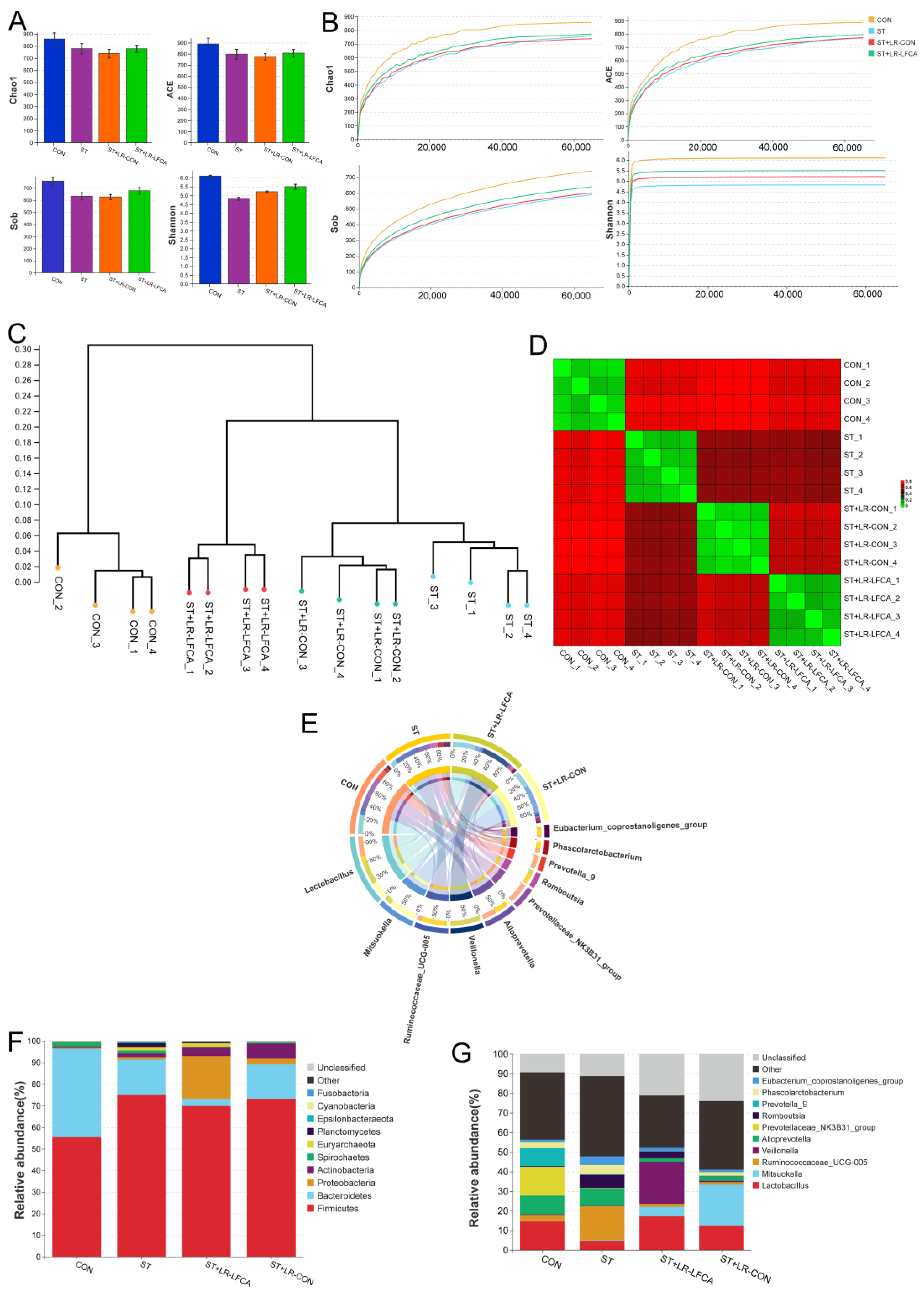
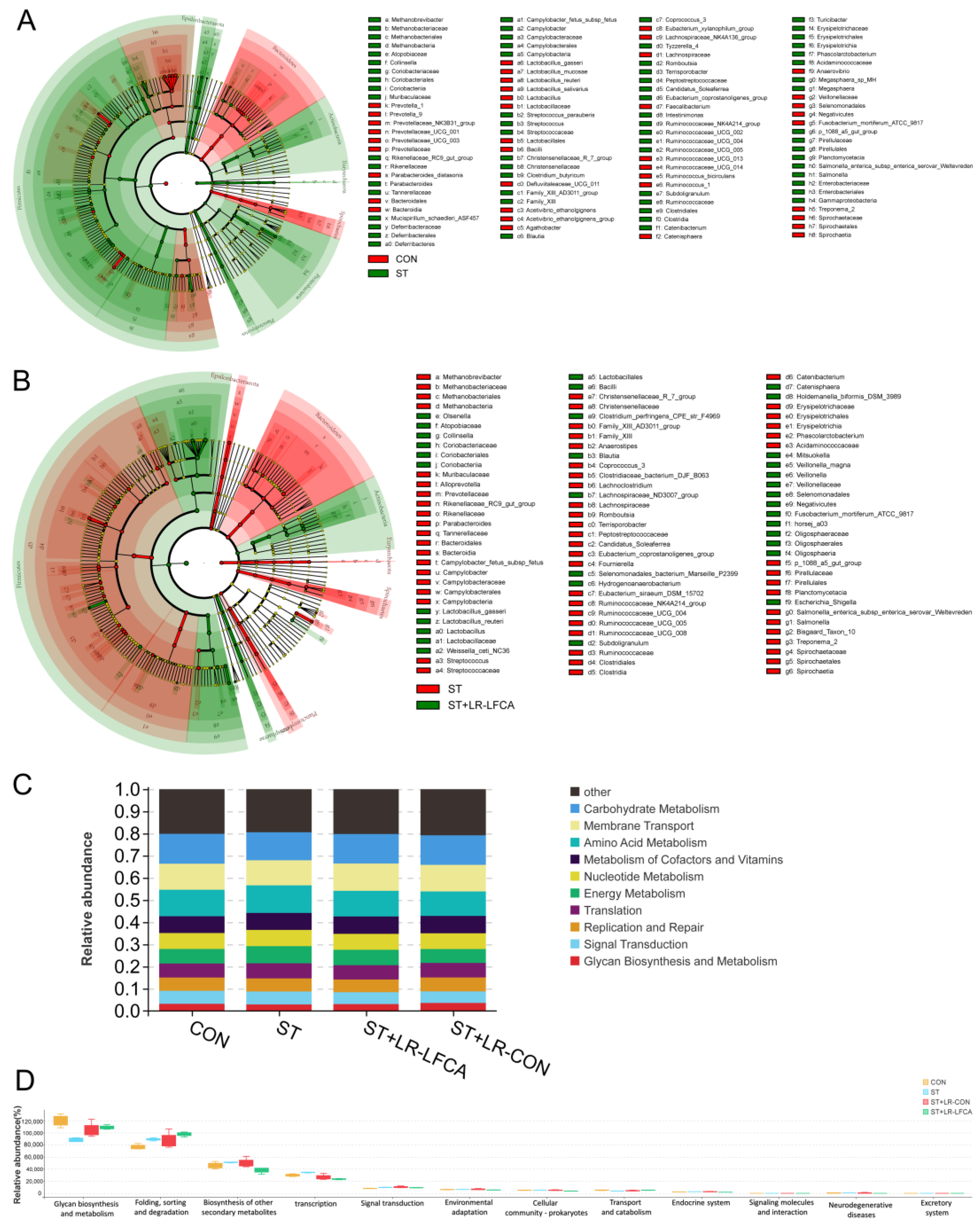
3.3. Oral Administration of Microencapsulated LR-LFCA to S. typhimurium-Infected Weaned Piglets Improved Gut Microbiota Composition
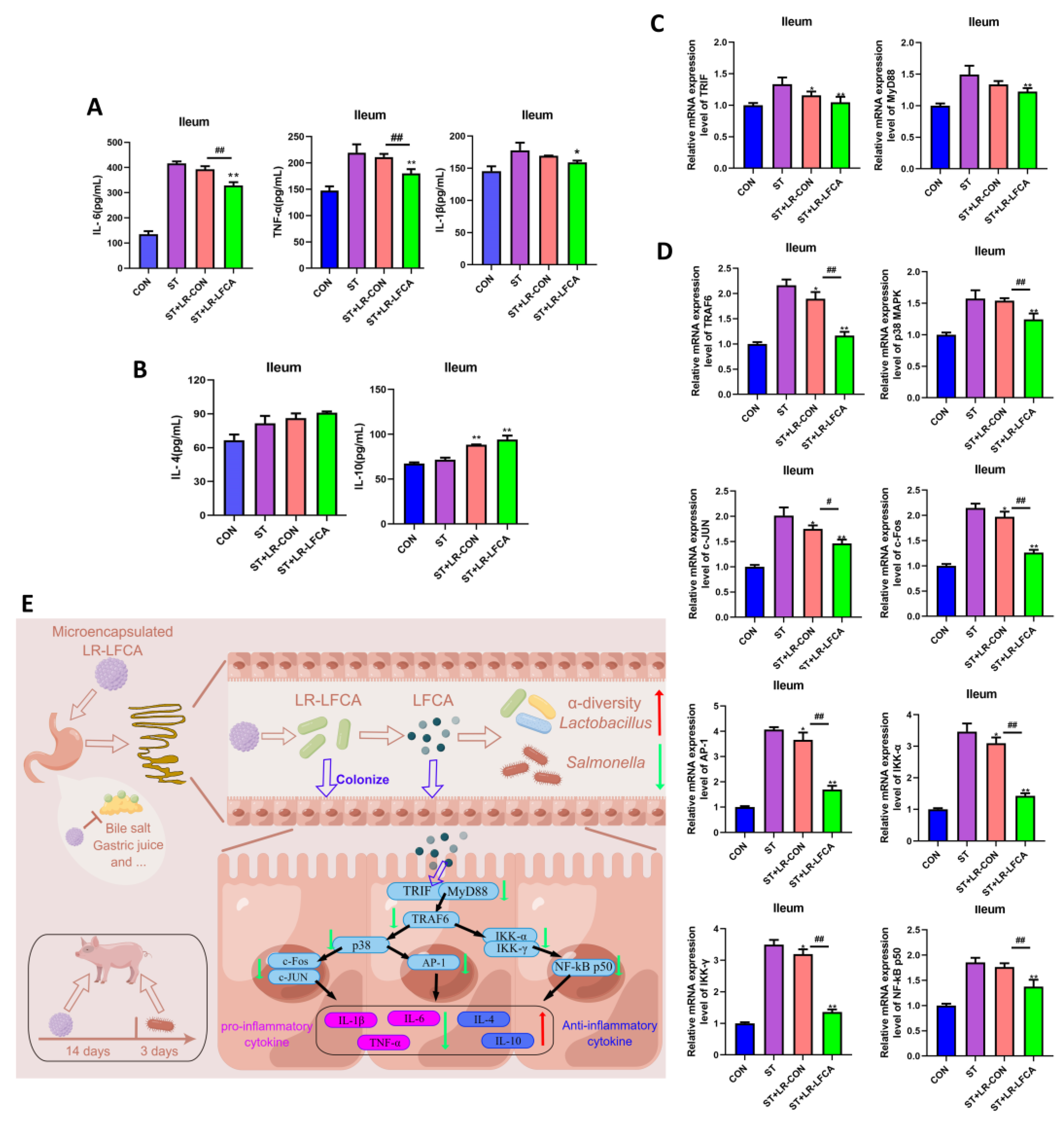
3.4. Oral Administration of Microencapsulated LR-LFCA to Weaned Piglets Protected against Changes in S. typhimurium-Induced Inflammatory Factor Levels in the Intestinal Mucosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Jasim, I.; Abdullah, A.; Shen, Z.; Zhao, L.; El-Dweik, M.; Zhang, S.; Almasri, M. An integrated impedance biosensor platform for detection of pathogens in poultry products. Sci. Rep. 2018, 8, 16109. [Google Scholar] [CrossRef] [PubMed]
- Teafatiller, T.; Subramanya, S.B.; Lambrecht, N.; Subramanian, V.S. Salmonella typhimurium infection reduces the ascorbic acid uptake in the intestine. Mediat. Inflamm. 2023, 2023, 2629262. [Google Scholar] [CrossRef]
- Kirthika, P.; Jawalagatti, V.; Senevirathne, A.; Lee, J.H. Coordinated interaction between Lon protease and catalase-peroxidase regulates virulence and oxidative stress management during Salmonellosis. Gut Microbes 2022, 14, 2064705. [Google Scholar] [CrossRef]
- Yeo, S.; Park, H.; Seo, E.; Kim, J.; Kim, B.K.; Choi, I.S.; Huh, C.S. Anti-inflammatory and gut microbiota modulatory effect of Lactobacillus rhamnosus strain LDTM 7511 in a dextran sulfate sodium-induced colitis murine model. Microorganisms 2020, 8, 845. [Google Scholar] [CrossRef]
- Jeon, M.S.; Choi, Y.Y.; Mo, S.J.; Ha, J.H.; Lee, Y.S.; Lee, H.U.; Park, S.D.; Shim, J.J.; Lee, J.L.; Chung, B.G. Contributions of the microbiome to intestinal inflammation in a gut-on-a-chip. Nano Converg. 2022, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Giordano, I.; Mauriello, G. Ultrasound Attenuation Improves some Surface Properties of the Probiotic Strain Lacticaseibacillus casei ATCC 393. Microorganisms 2023, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, Z.; Zhang, M.; Meng, L.; Ming, Z.; Liu, J. Bioinspired oral delivery of gut microbiota by self-coating with biofilms. Sci. Adv. 2020, 6, eabb1952. [Google Scholar] [CrossRef] [PubMed]
- Da, S.S.; Batista, L.; Diniz, D.S.; Nascimento, S.; Morais, N.S.; de Assis, C.F.; Passos, T.S.; de Sousa, J.F. Microencapsulation of probiotics by Oil-in-Water emulsification technique improves cell viability under different storage conditions. Foods 2023, 12, 252. [Google Scholar] [CrossRef]
- Huang, S.; Méjean, S.; Rabah, H.; Dolivet, A.; Le Loir, Y.; Chen, X.D.; Jan, G.; Jeantet, R.; Schuck, P. Double use of concentrated sweet whey for growth and spray drying of probiotics: Towards maximal viability in pilot scale spray dryer. J. Food Eng. 2017, 196, 11–17. [Google Scholar] [CrossRef]
- Ali, A.; Javaid, M.T.; Tazeddinova, D.; Khan, A.; Mehany, T.; Djabarovich, T.A.; Siddique, R.; Khalid, W.; Tariq, T.; Lai, W.F. Optimization of spray dried yogurt and its application to prepare functional cookies. Front. Nutr. 2023, 10, 1186469. [Google Scholar] [CrossRef]
- Tai, Z.; Zheng, M.; Yang, Y.; Xie, C.; Li, Z.; Xu, C. Temperature controlled microcapsule loaded with Perilla essential oil and its application in preservation of peaches. Front. Nutr. 2023, 10, 1087605. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.; Martins, E.; Perrone, I.T.; de Freitas, R.; Queiroz, L.S.; de Carvalho, A.F. Challenges associated with spray drying of lactic acid bacteria: Understanding cell viability loss. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3267–3283. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.; Cnossen, D.C.; Silva, C.; Cezarino, J.J.; Nero, L.A.; Perrone, I.T.; Carvalho, A.F. Determination of ideal water activity and powder temperature after spray drying to reduce Lactococcus lactis cell viability loss. J. Dairy Sci. 2019, 102, 6013–6022. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, S.; Sun, X.; Jie, Y.; Zhao, H.; Zhu, B.; Dziugan, P.; Zhang, B. A novel insight to screen the optimal spray-drying protectants and parameters for manufacturing lactic acid bacteria preparations. Dry. Technol. 2020, 38, 1843–1856. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L. Impact of a novel nano-protectant on the viability of probiotic bacterium Lactobacillus casei K17. Foods 2021, 10, 529. [Google Scholar] [CrossRef]
- Yi, H.; Hu, W.; Chen, S.; Lu, Z.; Wang, Y. Cathelicidin-WA improves intestinal epithelial barrier function and enhances host defense against enterohemorrhagic Escherichia coli O157:H7 infection. J. Immunol. 2017, 198, 1696–1705. [Google Scholar] [CrossRef]
- Sinha, M.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Antimicrobial lactoferrin peptides: The hidden players in the protective function of a multifunctional protein. Int. J. Pept. 2013, 2013, 390230. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Huang, K.; Shi, W.; Yang, B.; Wang, J. The probiotic and immunomodulation effects of Limosilactobacillus reuteri RGW1 isolated from calf feces. Front. Cell Infect. Microbiol. 2022, 12, 1086861. [Google Scholar] [CrossRef]
- Song, L.; Qiao, X.; Zhao, D.; Xie, W.; Bukhari, S.M.; Meng, Q.; Wang, L.; Cui, W.; Jiang, Y.; Zhou, H.; et al. Effects of Lactococcus lactis MG1363 producing fusion proteins of bovine lactoferricin-lactoferrampin on growth, intestinal morphology and immune function in weaned piglet. J. Appl. Microbiol. 2019, 127, 856–866. [Google Scholar] [CrossRef]
- Song, L.; Xie, W.; Liu, Z.; Guo, D.; Zhao, D.; Qiao, X.; Wang, L.; Zhou, H.; Cui, W.; Jiang, Y.; et al. Oral delivery of a Lactococcus lactis strain secreting bovine lactoferricin-lactoferrampin alleviates the development of acute colitis in mice. Appl. Microbiol. Biotechnol. 2019, 103, 6169–6186. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Song, L.; Wang, X.; Xu, Y.; Liu, Z.; Zhao, D.; Wang, S.; Fan, X.; Wang, Z.; Gao, C.; et al. A bovine lactoferricin-lactoferrampin-encoding Lactobacillus reuteri CO21 regulates the intestinal mucosal immunity and enhances the protection of piglets against enterotoxigenic Escherichia coli K88 challenge. Gut Microbes 2021, 13, 1956281. [Google Scholar] [CrossRef]
- Zhang, Q.; Widmer, G.; Tzipori, S. A pig model of the human gastrointestinal tract. Gut Microbes 2013, 4, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Morselli, M.; Su, T.; Million, M.; Larauche, M.; Pellegrini, M.; Tache, Y.; Yuan, P.Q. Comparative transcriptomics reveals highly conserved regional programs between porcine and human colonic enteric nervous system. Commun. Biol. 2023, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Wylensek, D.; Hitch, T.; Riedel, T.; Afrizal, A.; Kumar, N.; Wortmann, E.; Liu, T.; Devendran, S.; Lesker, T.R.; Hernandez, S.B.; et al. A collection of bacterial isolates from the pig intestine reveals functional and taxonomic diversity. Nat. Commun. 2020, 11, 6389. [Google Scholar] [CrossRef]
- Tilocca, B.; Burbach, K.; Heyer, C.; Hoelzle, L.E.; Mosenthin, R.; Stefanski, V.; Camarinha-Silva, A.; Seifert, J. Dietary changes in nutritional studies shape the structural and functional composition of the pigs’ fecal microbiome-from days to weeks. Microbiome 2017, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, S.J.; Tu, Z.C.; Wang, H.; Li, R.Y.; Zhang, L.; Huang, T.; Su, T.; Li, C. Quality evaluation of peony seed oil spray-dried in different combinations of wall materials during encapsulation and storage. J. Food Sci. Technol. 2016, 53, 2597–2605. [Google Scholar] [CrossRef]
- Pastorelli, G.; Serra, V.; Turin, L.; Redaelli, V.; Luzi, F.; Barbieri, S. Tranquillizing Effect of Passiflora incarnata Extract: Outcome on Behavioral and Physiological Indicators in Weaning Pigs with Intact Tails. Animals 2022, 12, 203. [Google Scholar] [CrossRef]
- Gao, K.; Wang, C.; Liu, L.; Dou, X.; Liu, J.; Yuan, L.; Zhang, W.; Wang, H. Immunomodulation and signaling mechanism of Lactobacillus rhamnosus GG and its components on porcine intestinal epithelial cells stimulated by lipopolysaccharide. J. Microbiol. Immunol. Infect. 2017, 50, 700–713. [Google Scholar] [CrossRef]
- Fan, Y.F.; Yang, Y.Y.; Yang, P.; Xia, T.; Ma, Y.X. Available energy content, nutrients digestibility of chili meal and effects on performance of growing pigs. Anim. Feed Sci. Technol. 2017, 229, 97–105. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, Y.; Xiong, Y.H.; Zhao, Y.Q.; Xiu, Z.; Ren, H.M.; Zhang, K.; Duan, S.; Chen, Y.; Xu, F.J. Infection-responsive long-term antibacterial bone plates for open fracture therapy. Bioact. Mater. 2023, 25, 1–12. [Google Scholar] [CrossRef]
- Antigo, J.L.D.; Bergamasco, R.D.C.; Madrona, G.S. Effect of ph on the stability of red beet extract (Beta vulgaris l.) microcapsules produced by spray drying or freeze drying. Food Sci. Technol. 2018, 38, 72–77. [Google Scholar] [CrossRef]
- Medina-Torres, L.; Calderas, F.; Nunez, R.D.; Herrera-Valencia, E.E.; Bernad, B.M.; Manero, O. Spray drying egg using either maltodextrin or nopal mucilage as stabilizer agents. J. Food Sci. Technol. 2017, 54, 4427–4435. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.; Oomah, B.D.; Rubilar, M.; Shene, C. Effective Lactobacillus plantarum and Bifidobacterium infantis encapsulation with chia seed (Salvia hispanica L.) and flaxseed (Linum usitatissimum L.) mucilage and soluble protein by spray drying. Food Chem. 2017, 216, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wu, H.; Lu, L.; Jiang, L.; Yu, Q. Lactobacillus reuteri promotes intestinal development and regulates mucosal immune function in newborn piglets. Front. Vet. Sci. 2020, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, C.; Liu, L.; Zhang, M. Lactobacillus reuteri KT260178 supplementation reduced morbidity of piglets through its targeted colonization, improvement of cecal microbiota profile, and immune functions. Probiotics Antimicrob. Proteins 2020, 12, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Li, W.; Li, Y.; Zhai, B.; Guo, Y.; Chen, Y.; Han, R.; Sun, G.; Jiang, R.; Li, Z.; et al. Genome-wide association study of 17 serum biochemical indicators in a chicken F2 resource population. BMC Genom. 2023, 24, 98. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, Y.; Lu, M.; Sun, W.; Sun, X.; Chen, X.; Li, L.; Chandoo, A.; Li, L. Controlling Nutritional Status (CONUT) score is a predictor of post-operative outcomes in elderly gastric cancer patients undergoing curative gastrectomy: A prospective study. Cancer Manag. Res. 2019, 11, 9793–9800. [Google Scholar] [CrossRef] [PubMed]
- Owczarzy, A.; Rogoz, W.; Kulig, K.; Pozycka, J.; Zieba, A.; Maciazek-Jurczyk, M. Spectroscopic studies of quinobenzothiazine derivative in terms of the in vitro interaction with selected human plasma proteins: Part 2. Molecules 2023, 28, 698. [Google Scholar] [CrossRef]
- Bogers, L.; Engelenburg, H.J.; Janssen, M.; Unger, P.A.; Melief, M.J.; Wierenga-Wolf, A.F.; Hsiao, C.C.; Mason, M.; Hamann, J.; van Langelaar, J.; et al. Selective emergence of antibody-secreting cells in the multiple sclerosis brain. EBioMedicine 2023, 89, 104465. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Zheng, N.; Guo, L.; Song, X.; Zhao, S.; Wang, J. Biological system responses of dairy cows to aflatoxin B1 exposure revealed with metabolomic changes in multiple biofluids. Toxins 2019, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lin, J.; Pan, T.; Li, T.; Jiang, H.; Fang, Y.; Wang, Y.; Wu, F.; Huang, J.; Zhang, H.; et al. Polymeric immunoglobulin receptor deficiency exacerbates autoimmune hepatitis by inducing intestinal dysbiosis and barrier dysfunction. Cell Death Dis. 2023, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, C.; Peng, M.; Wang, L.; Zhao, D.; Wu, T.; Yi, D.; Hou, Y.; Wu, G. N-Acetylcysteine improves intestinal function and attenuates intestinal autophagy in piglets challenged with beta-conglycinin. Sci. Rep. 2021, 11, 1261. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Yu, H.; Qiao, S. Lasso peptide microcin J25 effectively enhances gut barrier function and modulates inflammatory response in an enterotoxigenic Escherichia coli-challenged mouse model. Int. J. Mol. Sci. 2020, 21, 6500. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, Y.; Hu, Y.; Zhang, Y.; Wang, S. Lactobacillus rhamnosus GG reduces beta-conglycinin-allergy-induced apoptotic cells by regulating Bacteroides and bile secretion pathway in intestinal contents of BALB/c Mice. Nutrients 2020, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Bescucci, D.M.; Moote, P.E.; Ortega, P.R.; Uwiera, R.; Inglis, G.D. Salmonella enterica serovar Typhimurium temporally modulates the enteric microbiota and host responses to overcome colonization resistance in swine. Appl. Environ. Microbiol. 2020, 86, e01569-20. [Google Scholar] [CrossRef] [PubMed]
- Lapthorne, S.; Pereira-Fantini, P.M.; Fouhy, F.; Wilson, G.; Thomas, S.L.; Dellios, N.L.; Scurr, M.; O’Sullivan, O.; Ross, R.P.; Stanton, C.; et al. Gut microbial diversity is reduced and is associated with colonic inflammation in a piglet model of short bowel syndrome. Gut Microbes 2013, 4, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Chirullo, B.; Pesciaroli, M.; Drumo, R.; Ruggeri, J.; Razzuoli, E.; Pistoia, C.; Petrucci, P.; Martinelli, N.; Cucco, L.; Moscati, L.; et al. Salmonella Typhimurium exploits inflammation to its own advantage in piglets. Front. Microbiol. 2015, 6, 985. [Google Scholar] [CrossRef]
- Duan, B.; Shao, L.; Liu, R.; Msuthwana, P.; Hu, J.; Wang, C. Lactobacillus rhamnosus GG defense against Salmonella enterica serovar Typhimurium infection through modulation of M1 macrophage polarization. Microb. Pathog. 2021, 156, 104939. [Google Scholar] [CrossRef]
- Wang, R.; Li, S.; Jia, H.; Si, X.; Lei, Y.; Lyu, J.; Dai, Z.; Wu, Z. Protective Effects of Cinnamaldehyde on the Inflammatory Response, Oxidative Stress, and Apoptosis in Liver of Salmonella Typhimurium-Challenged Mice. Molecules 2021, 26, 2309. [Google Scholar] [CrossRef]
- Hu, J.L.; Yu, H.; Kulkarni, R.R.; Sharif, S.; Cui, S.W.; Xie, M.Y.; Nie, S.P.; Gong, J. Modulation of cytokine gene expression by selected Lactobacillus isolates in the ileum, caecal tonsils and spleen of Salmonella-challenged broilers. Avian Pathol. 2015, 44, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, X.; Xia, X.; Zhu, C.; Zhang, H.; Qin, W.; Xu, Y.; Hang, B.; Sun, Y.; Chen, S.; et al. Inhibitory Effects of antimicrobial peptide JH-3 on Salmonella enterica serovar Typhimurium strain CVCC541 infection-induced inflammatory cytokine release and apoptosis in RAW264.7 Cells. Molecules 2019, 24, 596. [Google Scholar] [CrossRef] [PubMed]
- Krypotou, E.; Townsend, G.E.; Gao, X.; Tachiyama, S.; Liu, J.; Pokorzynski, N.D.; Goodman, A.L.; Groisman, E.A. Bacteria require phase separation for fitness in the mammalian gut. Science 2023, 379, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ji, H.; Liu, H.; Wang, S.; Wang, J.; Wang, Y. Changes in the diversity and composition of gut microbiota of weaned piglets after oral administration of Lactobacillus or an antibiotic. Appl. Microbiol. Biotechnol. 2016, 100, 10081–10093. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Malik, A.; Steury, M.D.; Lucas, P.C.; Parameswaran, N. Protective role of beta-arrestin2 in colitis through modulation of T-cell activation. Inflamm. Bowel Dis. 2015, 21, 2766–2777. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Xie, W.; Cai, L.; Han, C.; Kuang, H.; Shao, Y.; Zhang, S.; Zhang, Q.; Li, J.; Cui, W.; et al. Microencapsulated Limosilactobacillus reuteri Encoding Lactoferricin-Lactoferrampin Targeted Intestine against Salmonella typhimurium Infection. Nutrients 2023, 15, 5141. https://doi.org/10.3390/nu15245141
Wang X, Xie W, Cai L, Han C, Kuang H, Shao Y, Zhang S, Zhang Q, Li J, Cui W, et al. Microencapsulated Limosilactobacillus reuteri Encoding Lactoferricin-Lactoferrampin Targeted Intestine against Salmonella typhimurium Infection. Nutrients. 2023; 15(24):5141. https://doi.org/10.3390/nu15245141
Chicago/Turabian StyleWang, Xueying, Weichun Xie, Limeng Cai, Chuang Han, Hongdi Kuang, Yilan Shao, Senhao Zhang, Qi Zhang, Jiaxuan Li, Wen Cui, and et al. 2023. "Microencapsulated Limosilactobacillus reuteri Encoding Lactoferricin-Lactoferrampin Targeted Intestine against Salmonella typhimurium Infection" Nutrients 15, no. 24: 5141. https://doi.org/10.3390/nu15245141
APA StyleWang, X., Xie, W., Cai, L., Han, C., Kuang, H., Shao, Y., Zhang, S., Zhang, Q., Li, J., Cui, W., Jiang, Y., & Tang, L. (2023). Microencapsulated Limosilactobacillus reuteri Encoding Lactoferricin-Lactoferrampin Targeted Intestine against Salmonella typhimurium Infection. Nutrients, 15(24), 5141. https://doi.org/10.3390/nu15245141