Effects of Cinnamon (Cinnamomum zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet



Abstract
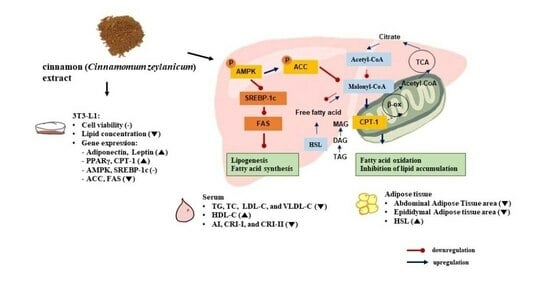
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Cinnamon Extract
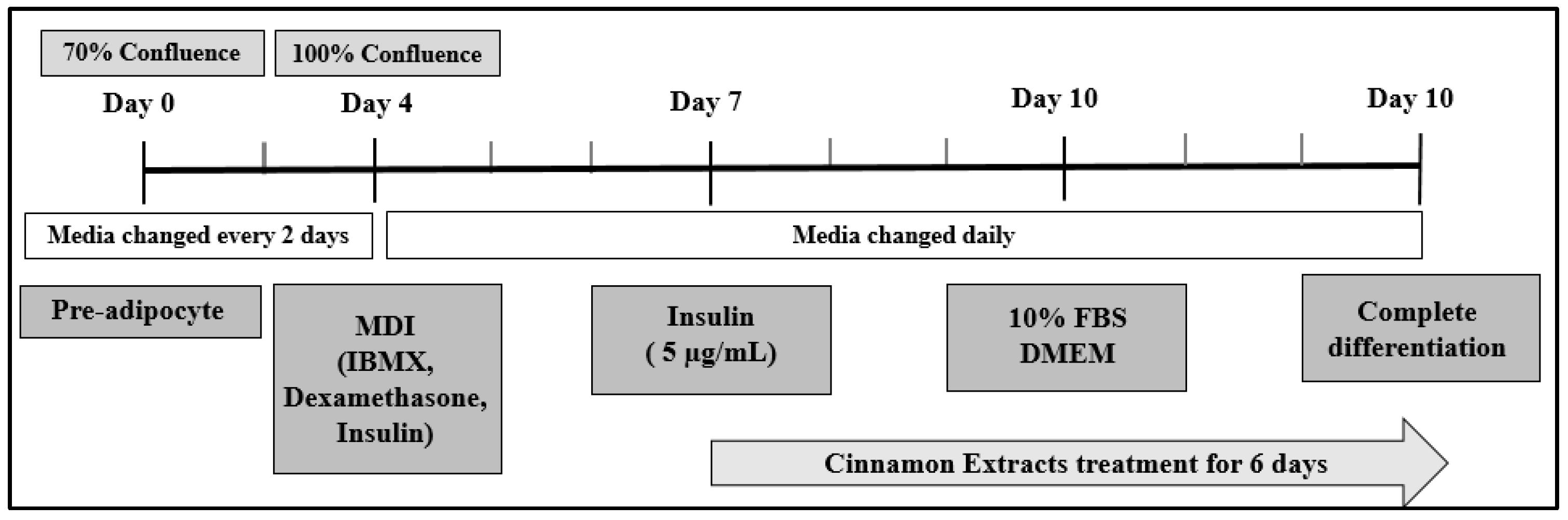
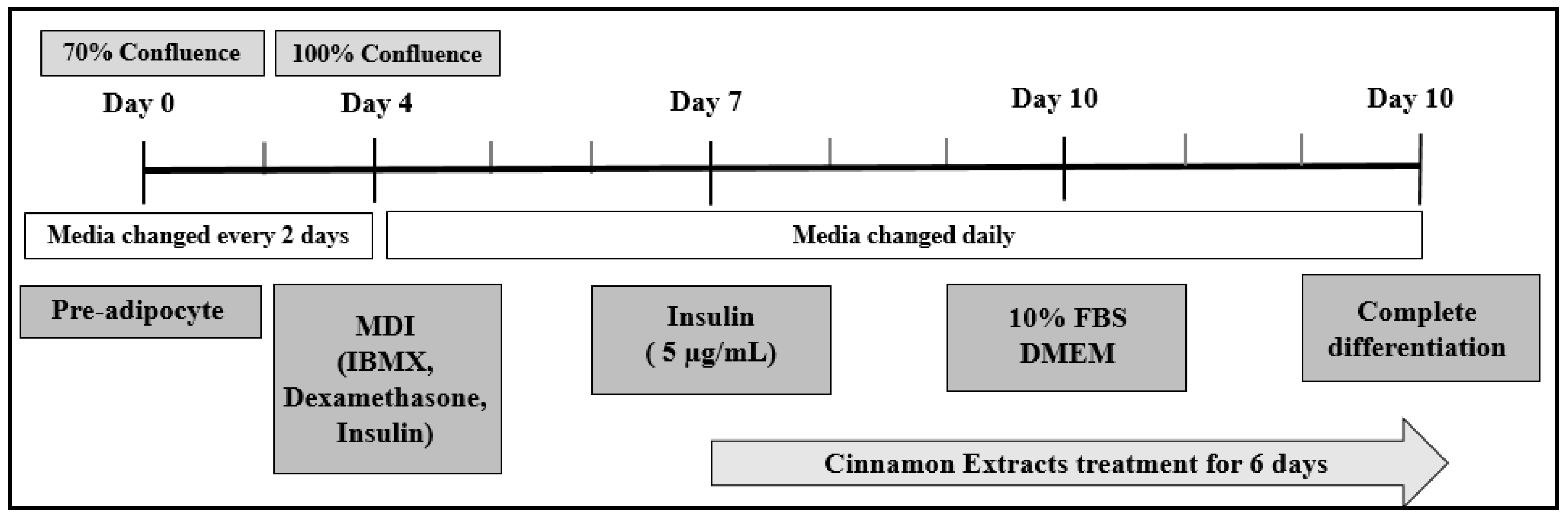
2.3. Cell Culture
2.4. MTT Assay
2.5. Oil Red O Staining
2.6. mRNA Expression Analysis
2.7. Preparation of Diets
2.8. Animals and Housing Conditions
2.9. Experimental Design of Animal Groups
2.10. Measurement of Body Weight and Food Intake
2.11. Blood and Tissue Collection
2.12. Lipid Profiles
2.13. Atherogenic Index (AI), Cardiac Risk Index (CRI) I, and II
2.14. Hepatic TG and TC Levels
2.15. Hormone-Sensitive Lipase in the Liver and WAT
2.16. Histological Analysis
2.17. Western Blotting
2.18. Statistical Analysis
3. Results
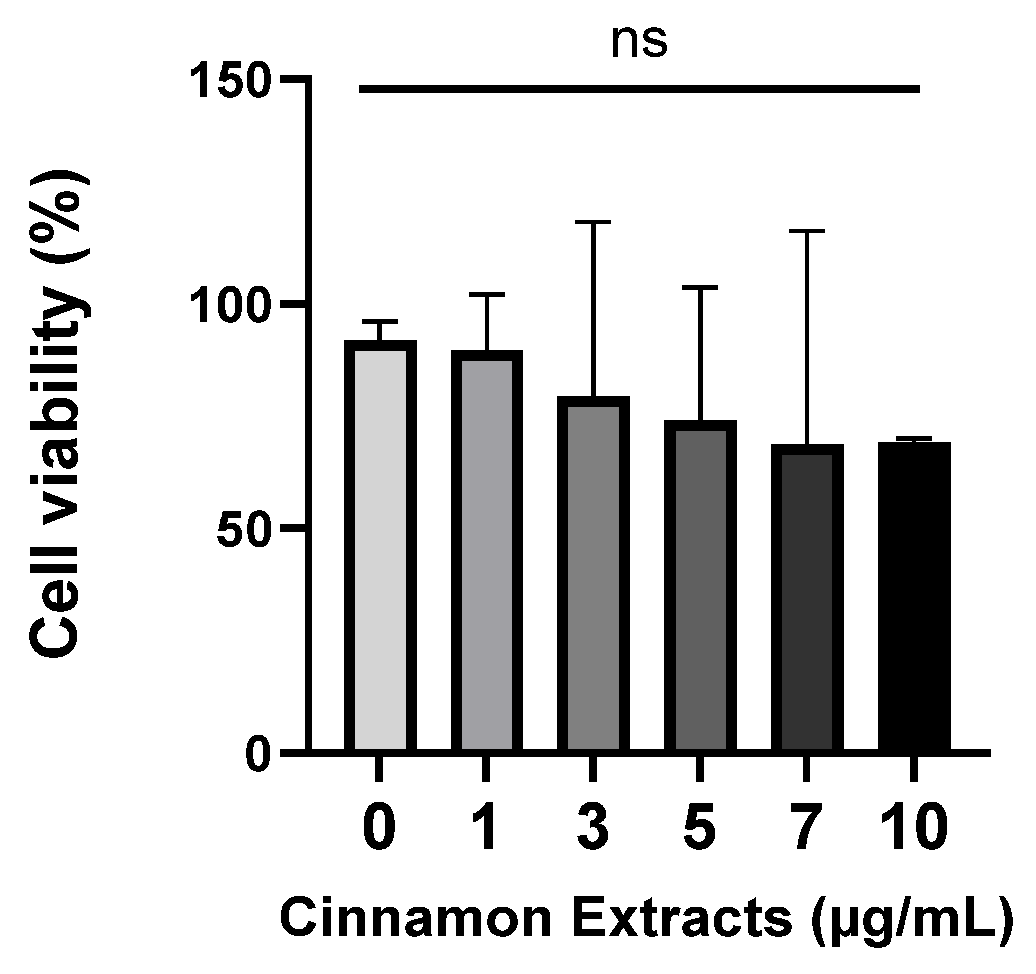
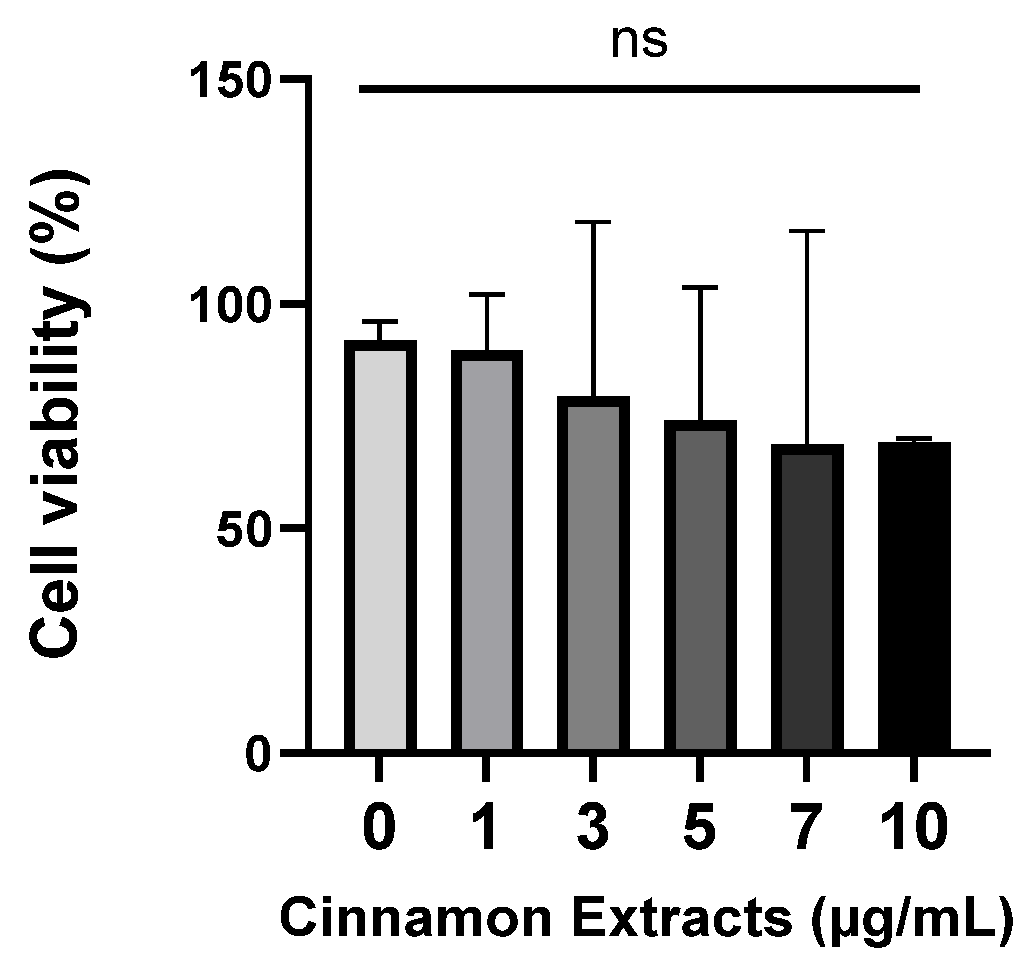
3.1. Effects of Cinnamon Extract on Cell Viability in Pre-Adipocytes
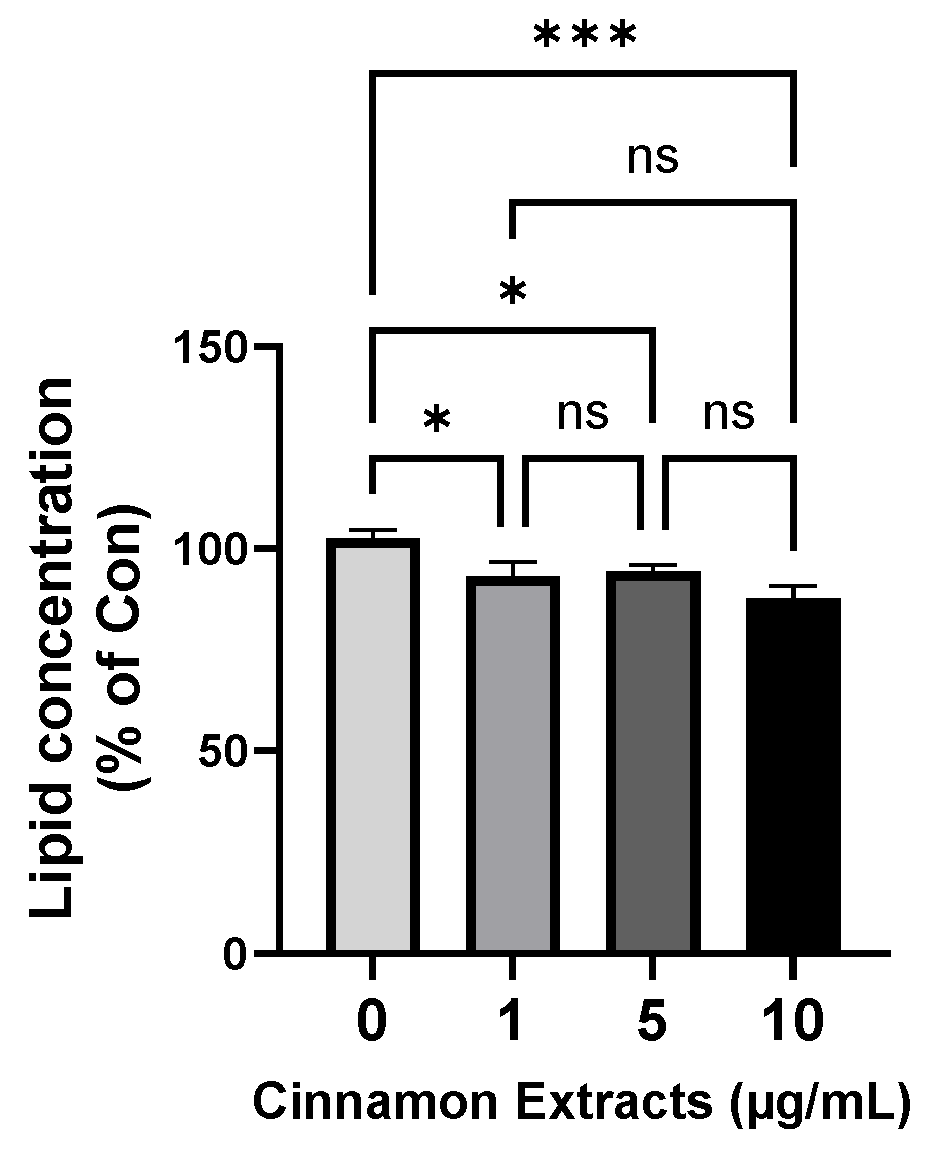
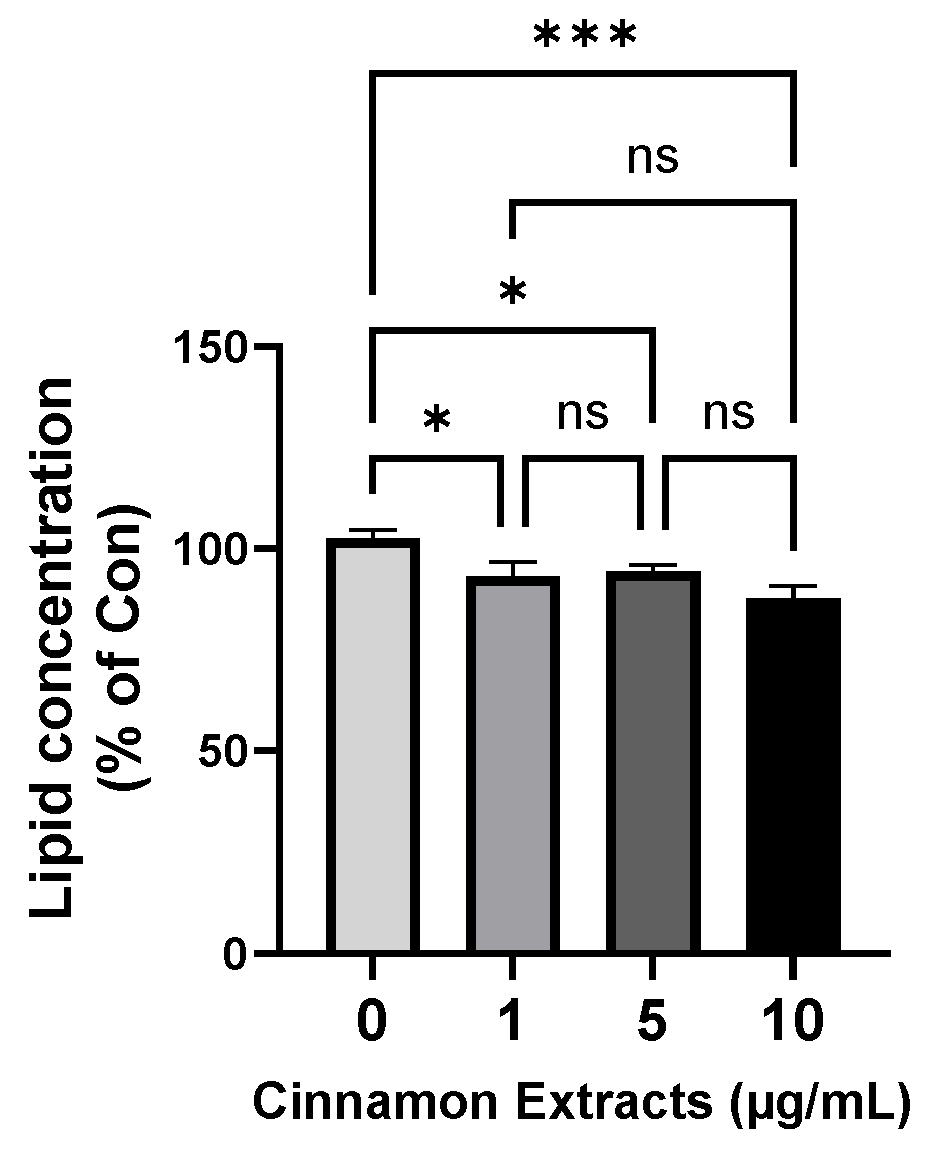
3.2. Effects of Cinnamon Extract on Lipid Concentration
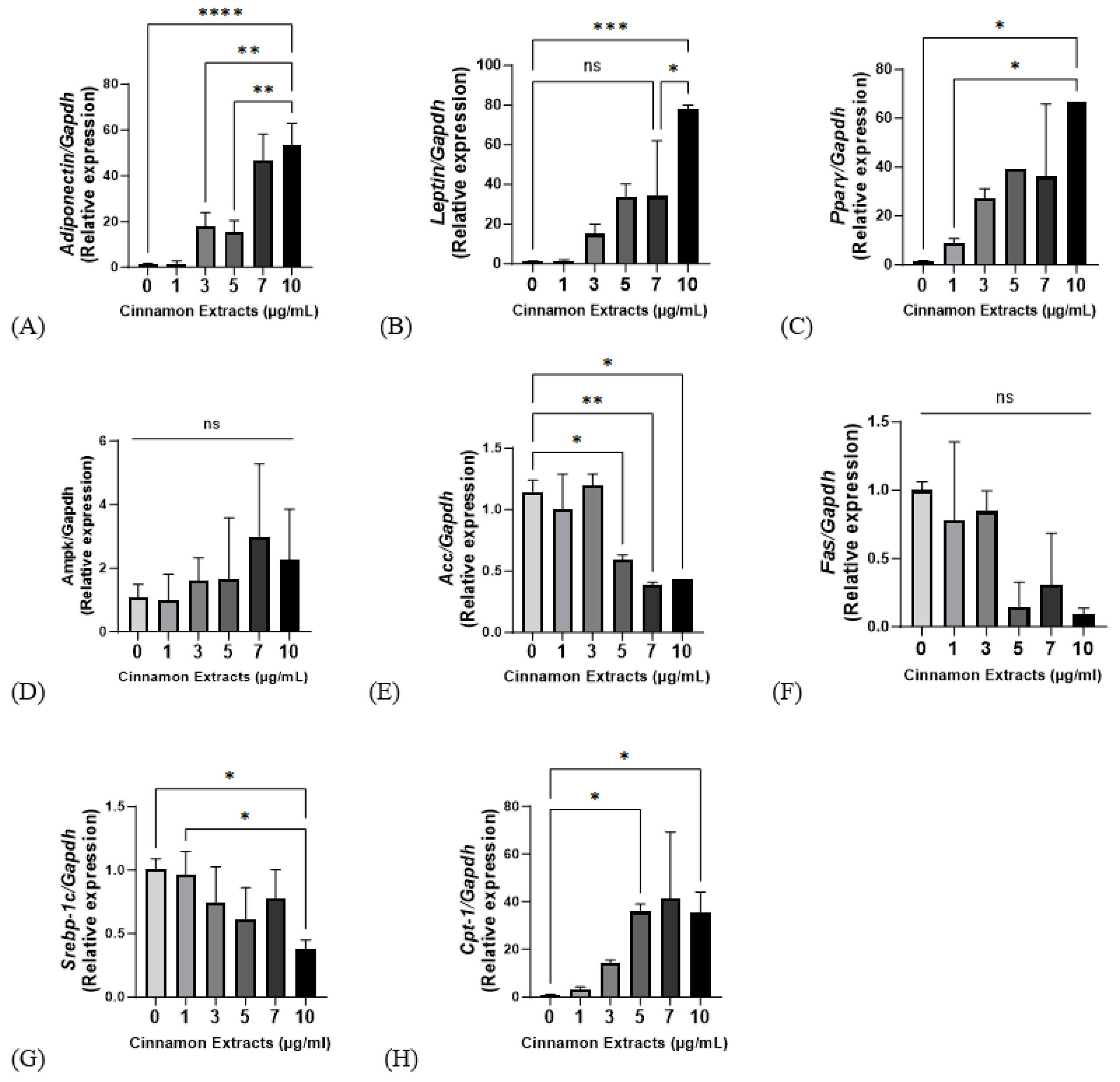
3.3. Effects of Cinnamon Extract on the mRNA Expression Levels of Genes Related to Lipid Metabolism: Antiadipogenic Effect, Lipolysis, and Lipid Synthesis in 3T3-L1 Cells
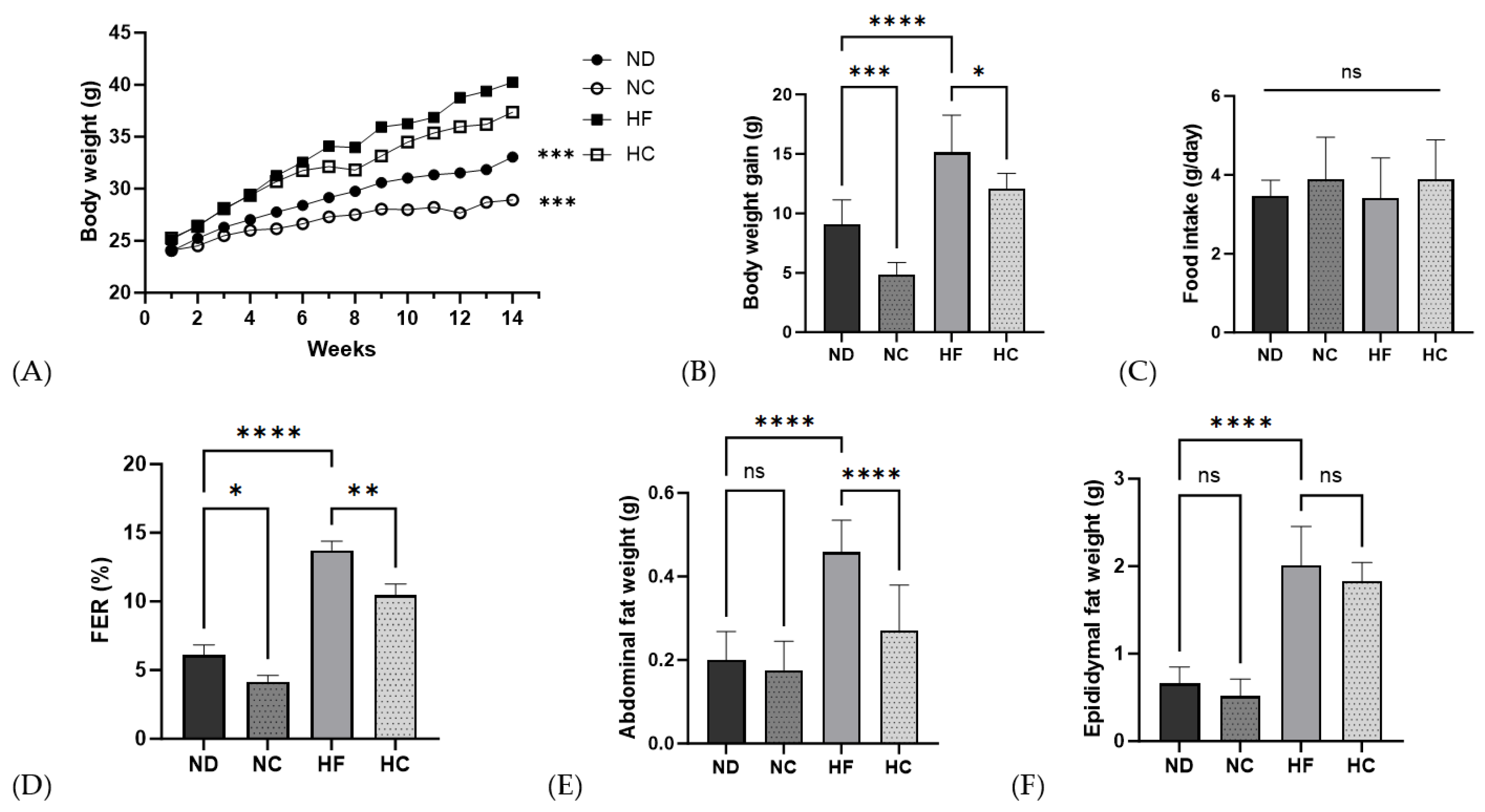
3.4. Effects of Cinnamon Extract on Body Weight, Body Weight Gain, Food Intake, FER, and Organ Weight of Fat
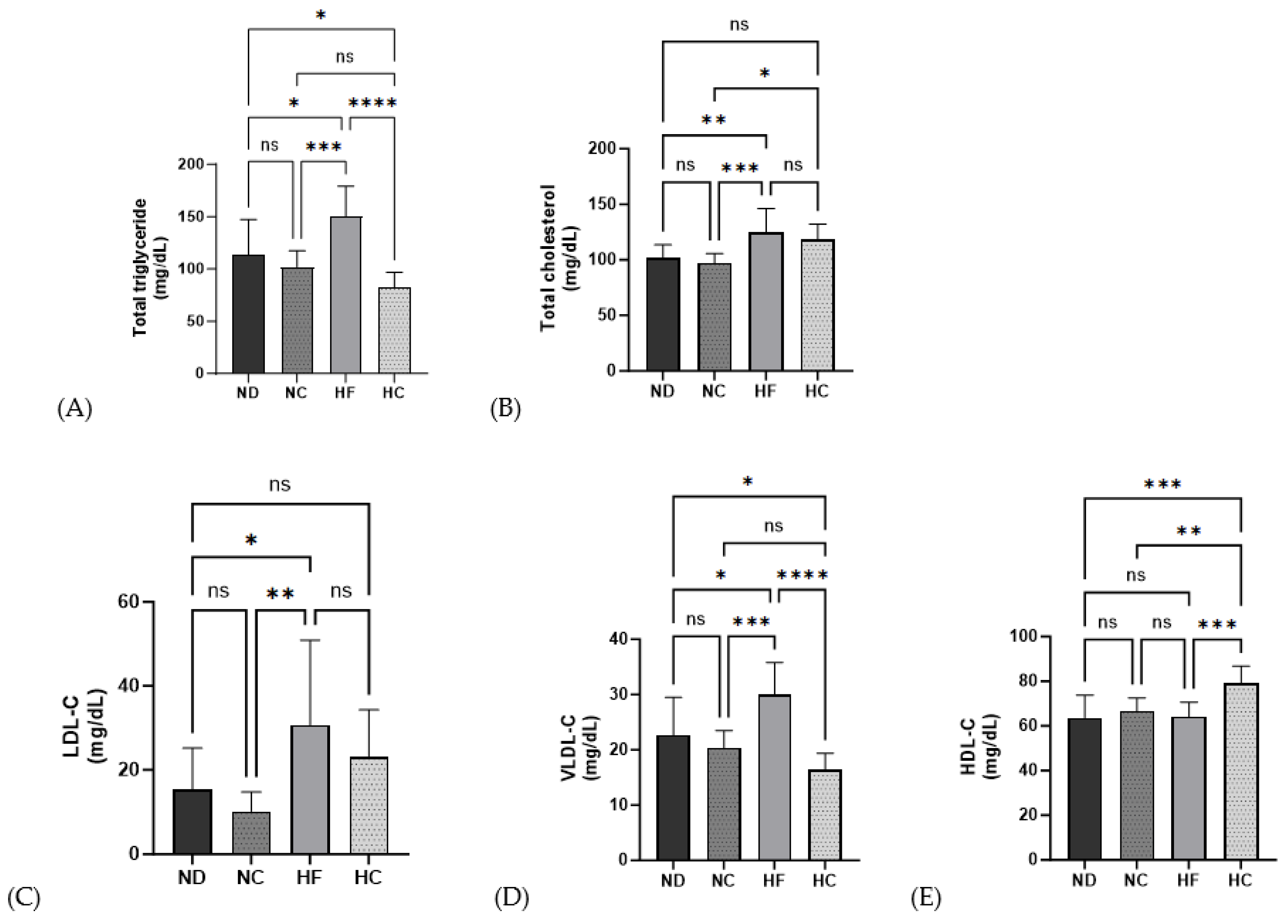
3.5. Effects of Cinnamon Extract on Serum Lipid Profiles
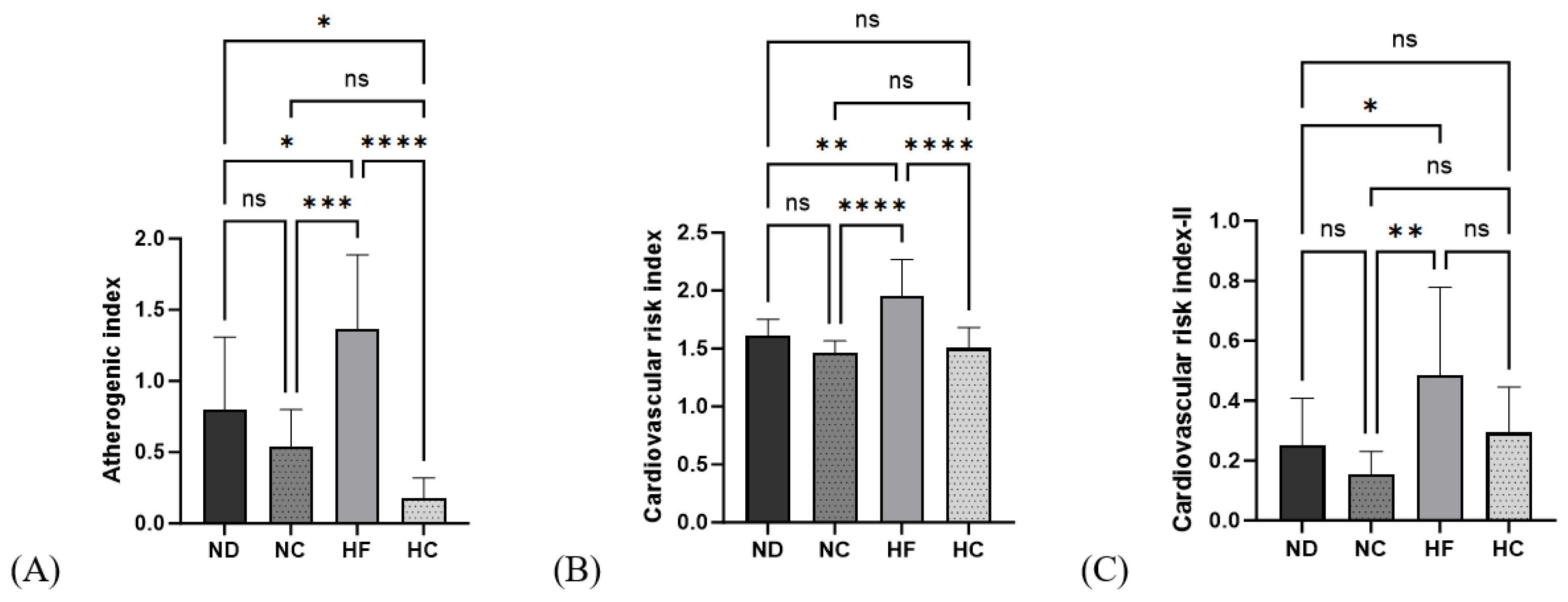
3.6. Effects of Cinnamon Extract on the Atherogenic Index (AI) and Cardiac Risk Index (CRI-I, CRI-II)
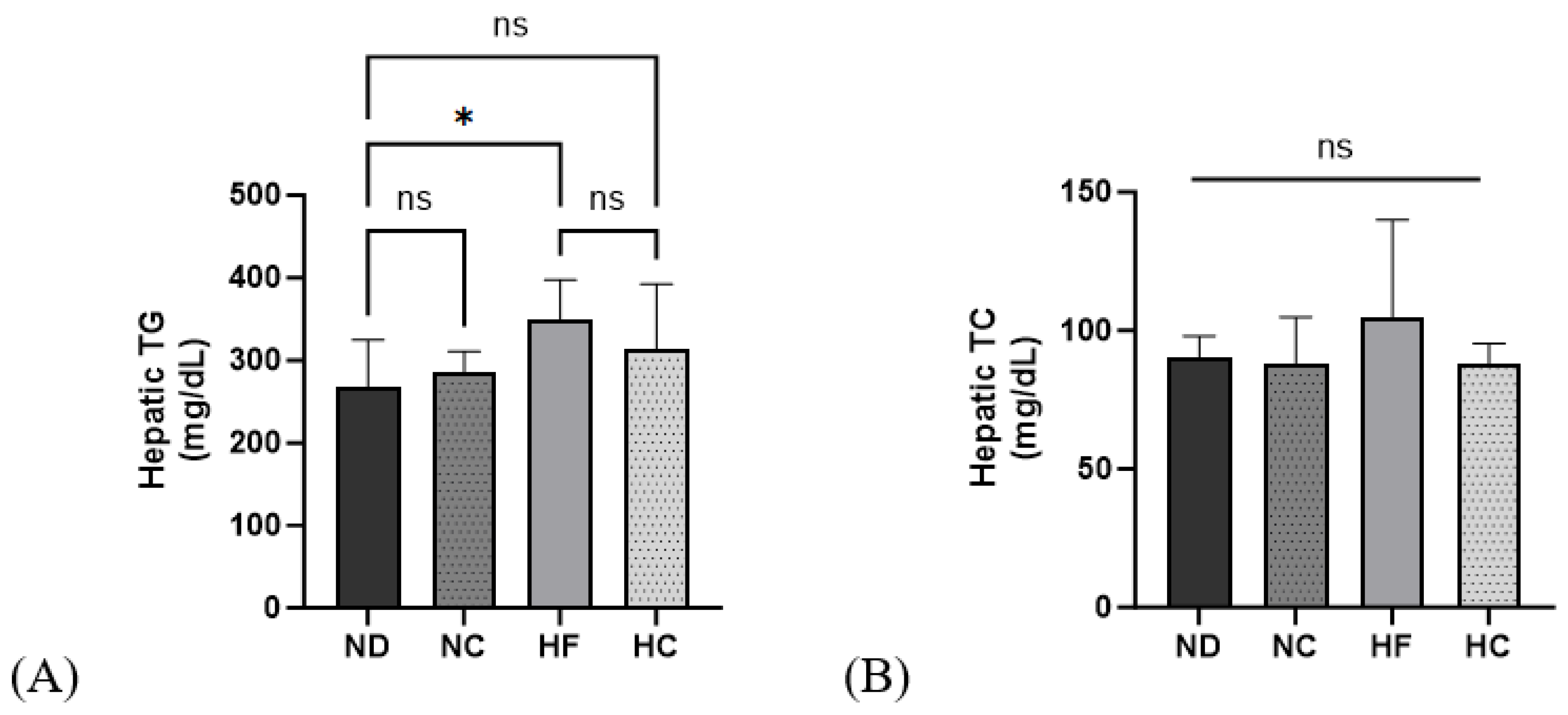
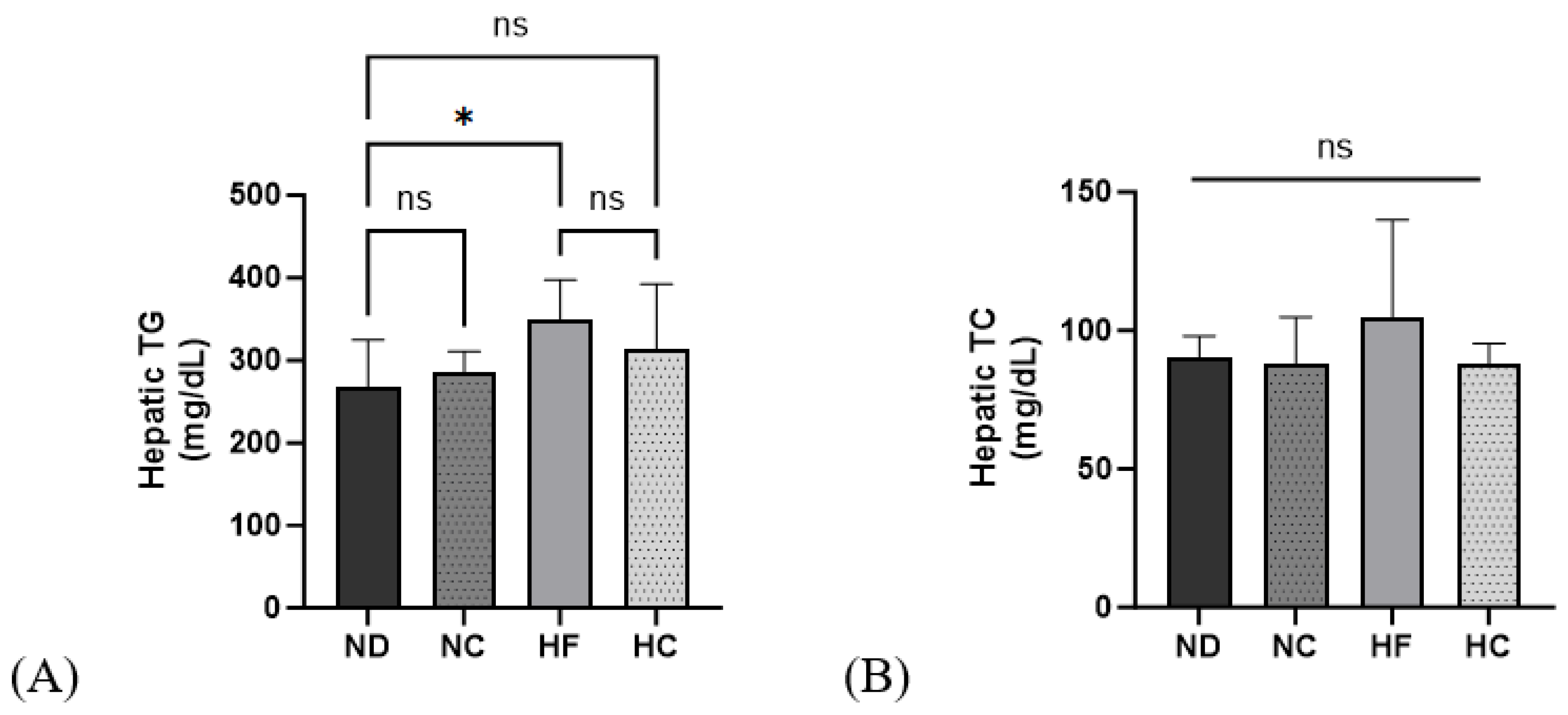
3.7. Effects of Cinnamon Extract on Hepatic TG and TC Levels
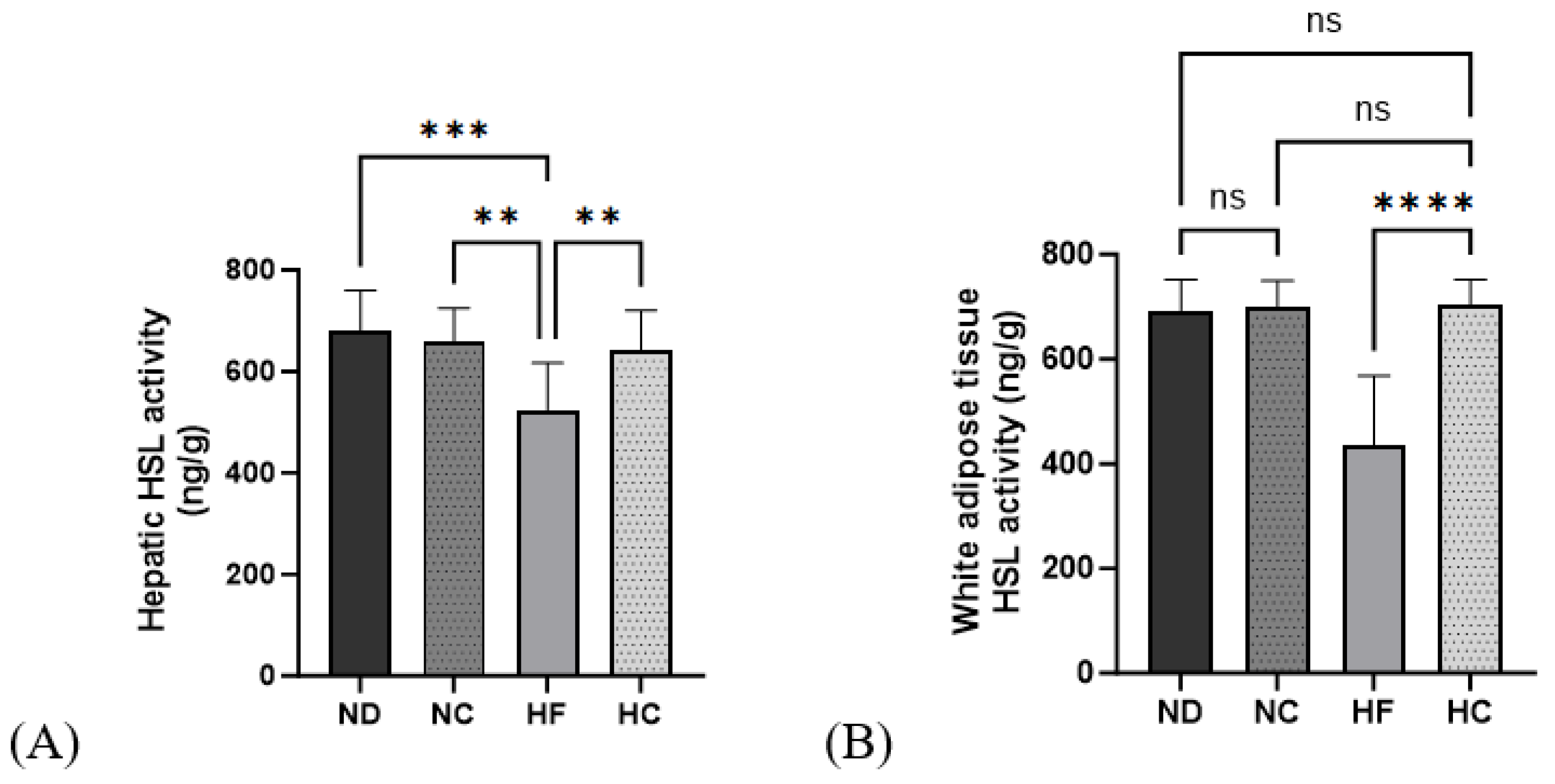
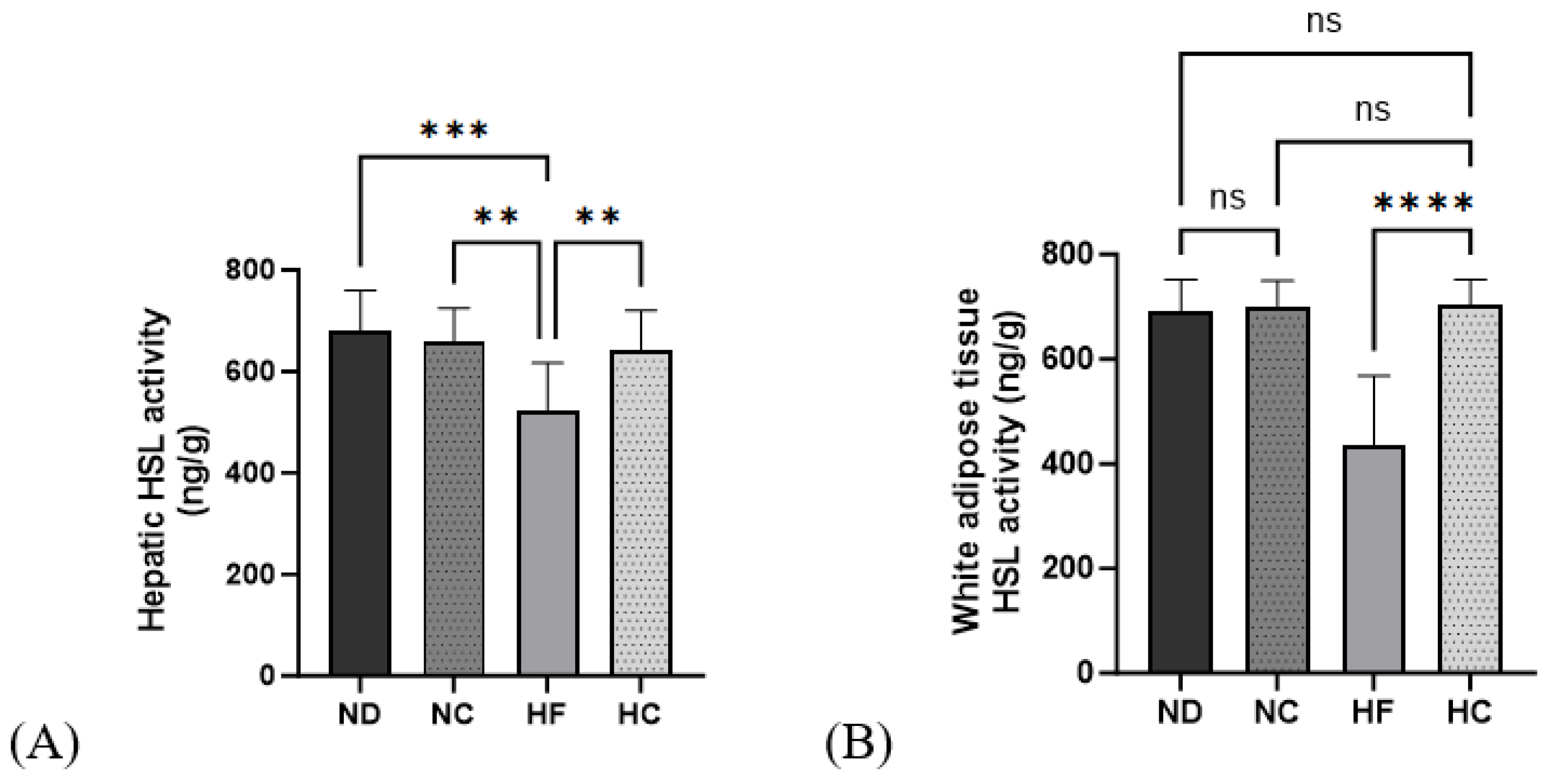
3.8. Effects of Cinnamon Extract on HSL Activity in the Liver and WAT
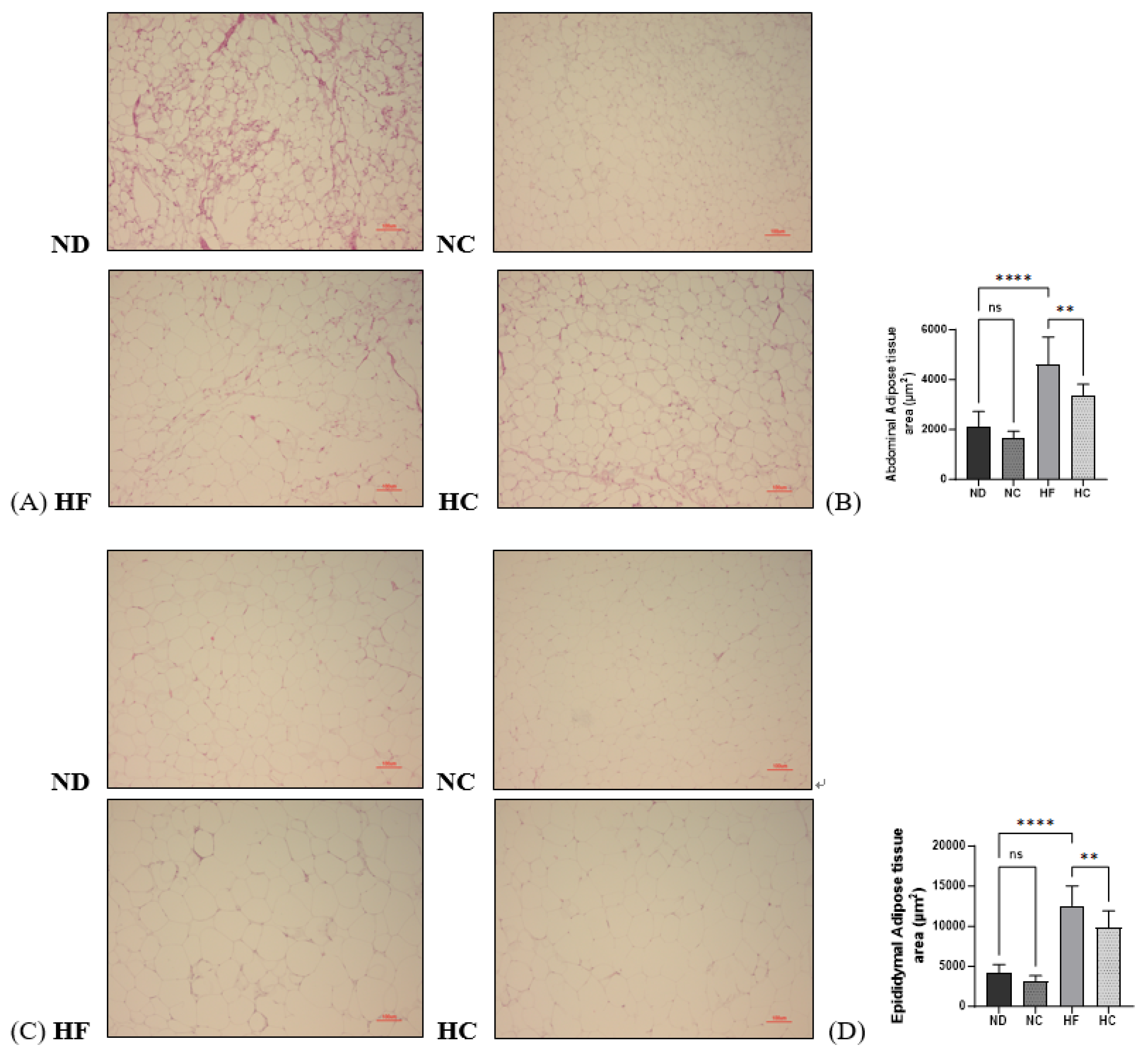
3.9. Effects of Cinnamon Extract on the Adipose Tissue Area (µm2) of Abdominal Adipose Tissue and Epididymal Adipose Tissue
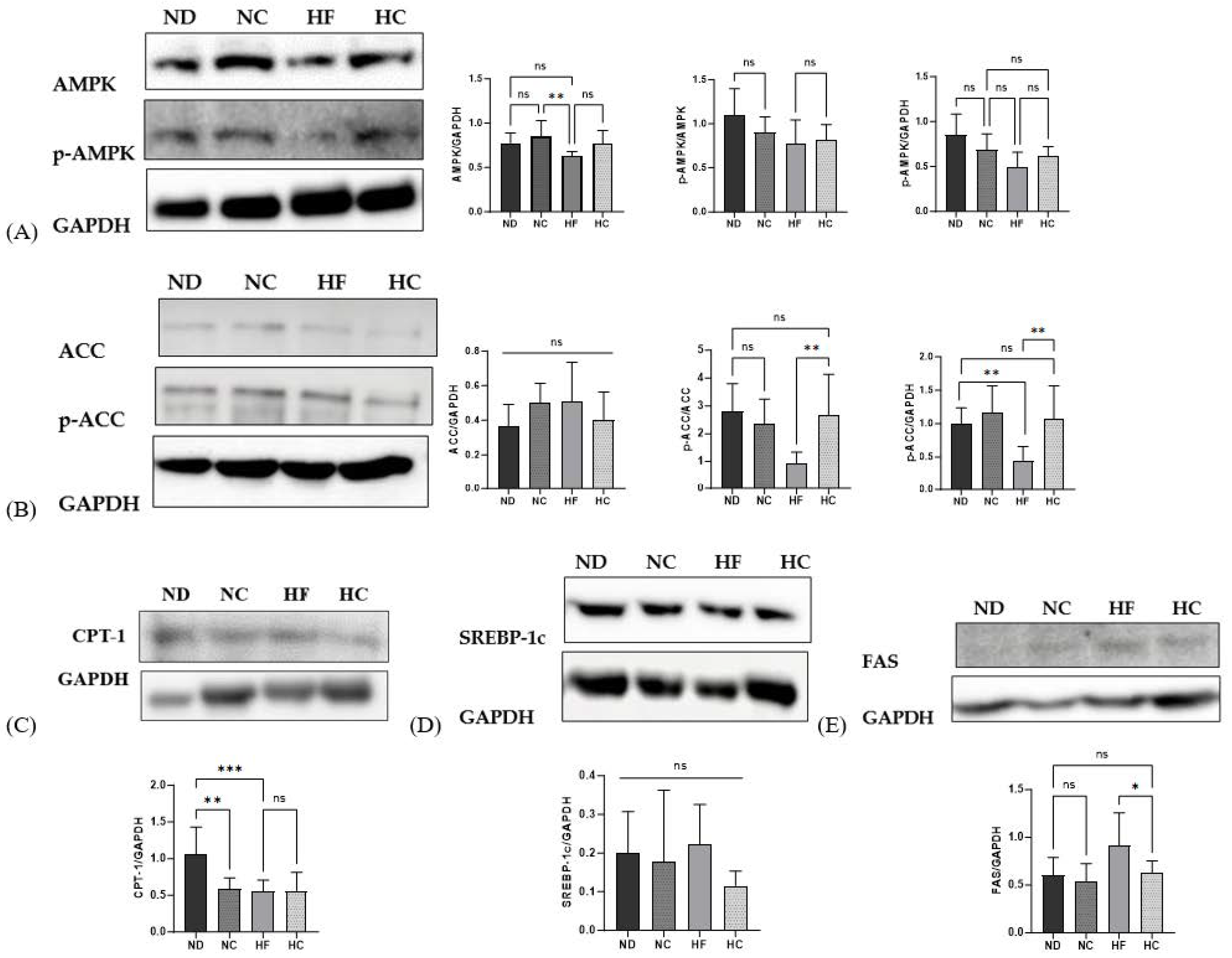
3.10. Effects of Cinnamon Extract on the Protein Expression Levels, as Related to Adipogenic, Lipolysis, and Lipid Synthesis Markers in the Liver
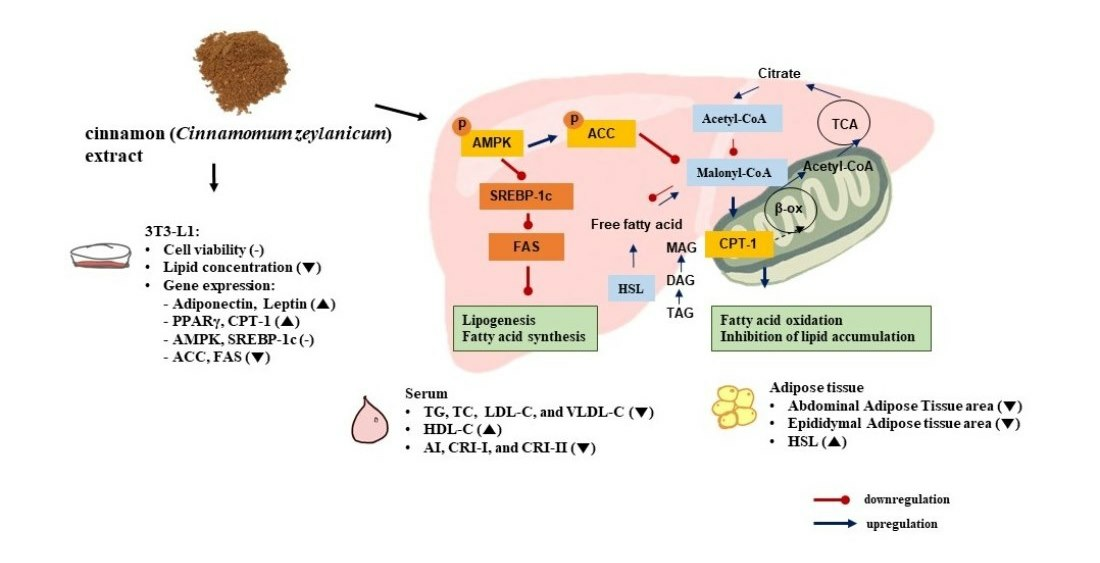
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, E.S.; Choi, S.J.; Kim, H.D.; Kang, M.H.; Ji, Y.-J.; Kim, G.-S.; Jang, G.Y. Anti-Obesity Activity in 3T3-L1 Cells of Cornus officinalis Fruits Harvested at Different Times. Processes 2022, 10, 2008. [Google Scholar] [CrossRef]
- Ezenwaka, C.E.; Okoye, O.; Esonwune, C.; Onuoha, P.; Dioka, C.; Osuji, C.; Oguejiofor, C.; Meludu, S. High prevalence of abdominal obesity increases the risk of the metabolic syndrome in Nigerian type 2 diabetes patients: Using the International Diabetes Federation worldwide definition. Metab. Syndr. Relat. Disord. 2014, 12, 277–282. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.; Jo, M.J.; Cho, E.; Ahn, S.Y.; Kwon, Y.J.; Ko, G.J. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef] [PubMed]
- Suriyagoda, L.; Mohotti, A.J.; Vidanarachchi, J.K.; Kodithuwakku, S.P.; Chathurika, M.; Bandaranayake, P.C.G.; Hetherington, A.M.; Beneragama, C.K. “Ceylon cinnamon”: Much more than just a spice. Plants People Planet. 2021, 3, 319–336. [Google Scholar] [CrossRef]
- Lee, J.H.; Choi, K.S. Assessment of Nutrient Intake and Dietary Quality of Korean Adults in Metabolic Syndrome Patients According to Taking Medical Care: Based on the 2017 Korea National Health and Nutrition Examination Survey. Korean J. Community Nutr. 2022, 27, 321–340. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Jagan Mohan Rao, L. Chemistry, Biogenesis, and Biological Activities of Cinnamomum zeylanicum. Crit. Rev. Food Sci. Nutr. 2011, 51, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Gan, S.H. Cinnamon: A multifaceted medicinal plant. Evid.-Based Complement. Altern. Med. Ecam 2014, 2014, 642942. [Google Scholar] [CrossRef]
- Vangalapati, M.; Satya, N.S.; Prakash, D.S.; Avanigadda, S. A review on pharmacological activities and clinical effects of cinnamon species. Res. J. Pharm. Biol. Chem. Sci. 2012, 3, 653–663. [Google Scholar]
- Muhoza, B.; Qi, B.; Harindintwali, J.D.; Koko, M.Y.F.; Zhang, S.; Li, Y. Encapsulation of cinnamaldehyde: An insight on delivery systems and food applications. Crit. Rev. Food Sci. Nutr. 2023, 63, 2521–2543. [Google Scholar] [CrossRef]
- Srinivasan, K. Antioxidant potential of spices and their active constituents. Crit. Rev. Food Sci. Nutr. 2014, 54, 352–372. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Ren, J.; Kong, L.; Yan, G.; Liu, C.; Han, Y.; Sun, H.; Wang, X.J. Ephedrae Herba: A Review of Its Phytochemistry, Pharmacology, Clinical Application, and Alkaloid Toxicity. Molecules 2023, 28, 663. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Choung, S.Y. Antihyperglycemic and antihyperlipidemic action of Cinnamomi Cassiae (Cinnamon bark) extract in C57BL/Ks db/db mice. Arch. Pharmacal Res. 2010, 33, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.P. Obesity and the development of type 2 diabetes: The effects of fatty tissue inflammation. Diabetes Metab. Syndr. Obes. 2010, 3, 253–262. [Google Scholar] [CrossRef]
- Konige, M.; Wang, H.; Sztalryd, C. Role of adipose specific lipid droplet proteins in maintaining whole body energy homeostasis. Biochim. Biophys. Acta. 2014, 1842, 393–401. [Google Scholar] [CrossRef]
- Park, C.H.; Rhyu, D.Y.; Sharma, B.R.; Yokozawa, T. Inhibition of preadipocyte differentiation and lipid accumulation by 7-O-galloyl-d-sedoheptulose treatment in 3T3-L1 adipocytes. Biomed. Prev. Nutr. 2013, 3, 319–324. [Google Scholar] [CrossRef]
- Etesami, B.; Ghaseminezhad, S.; Nowrouzi, A.; Rashidipour, M.; Yazdanparast, R. Investigation of 3T3-L1 Cell Differentiation to Adipocyte, Affected by Aqueous Seed Extract of Phoenix dactylifera L. Rep. Biochem. Mol. Biol. 2020, 9, 14–25. [Google Scholar] [CrossRef]
- Peterson, J.M.; Aja, S.; Wei, Z.; Wong, G.W. CTRP1 protein enhances fatty acid oxidation via AMP-activated protein kinase (AMPK) activation and acetyl-CoA carboxylase (ACC) inhibition. J. Biol. Chem. 2012, 287, 1576–1587. [Google Scholar] [CrossRef]
- Ke, R.; Xu, Q.; Li, C.; Luo, L.; Huang, D. Mechanisms of AMPK in the maintenance of ATP balance during energy metabolism. Cell Biol. Int. 2018, 42, 384–392. [Google Scholar] [CrossRef]
- Fang, K.; Wu, F.; Chen, G.; Dong, H.; Li, J.; Zhao, Y.; Xu, L.; Zou, X.; Lu, F. Diosgenin ameliorates palmitic acid-induced lipid accumulation via AMPK/ACC/CPT-1A and SREBP-1c/FAS signaling pathways in LO2 cells. BMC Complement. Altern. Med. 2019, 19, 255. [Google Scholar] [CrossRef]
- Scaglia, N.; Chisholm, J.W.; Igal, R.A. Inhibition of StearoylCoA Desaturase-1 Inactivates Acetyl-CoA Carboxylase and Impairs Proliferation in Cancer Cells: Role of AMPK. PLoS ONE 2009, 4, e6812. [Google Scholar] [CrossRef]
- Schindler, M.; Pendzialek, M.; Grybel, K.J.; Seeling, T.; Gürke, J.; Fischer, B.; Navarrete Santos, A. Adiponectin stimulates lipid metabolism via AMPK in rabbit blastocysts. Hum. Reprod. 2017, 32, 1382–1392. [Google Scholar] [CrossRef] [PubMed]
- Lingesh, A.; Paul, D.; Naidu, V.; Satheeshkumar, N. AMPK activating and anti adipogenic potential of Hibiscus rosa sinensis flower in 3T3-L1 cells. J. Ethnopharmacol. 2019, 233, 123–130. [Google Scholar] [CrossRef]
- Ebrahimi, B.; Baroutian, S.; Li, J.; Zhang, B.; Ying, T.; Lu, J. Combination of marine bioactive compounds and extracts for the prevention and treatment of chronic diseases. Front. Nutr. 2023, 9, 1047026. [Google Scholar] [CrossRef] [PubMed]
- DeBose-Boyd, R.A.; Ye, J. SREBPs in Lipid Metabolism, Insulin Signaling, and Beyond. Trends Biochem. Sci. 2018, 43, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-H.; Liu, Y.; Kim, H.-S. Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats. Nutrients 2016, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.Y.; Jeong, I.K.; Nam, S.Y.; Kim, B.W.; Kwon, H.J. Inhibitory Effects and Molecular Mechanism of Adipocyte Differentiation by Rosae laevigata Fructus Ethanol Extracs. Microbiol. Biotechnol. Lett. 2016, 44, 89–97. [Google Scholar] [CrossRef]
- Lee, M.R.; Kim, J.E.; Choi, J.Y.; Park, J.J.; Kim, H.R.; Song, B.R.; Park, J.W.; Kang, M.J.; Choi, Y.W.; Kim, K.M.; et al. Morusin Functions as a Lipogenesis Inhibitor as Well as a Lipolysis Stimulator in Differentiated 3T3-L1 and Primary Adipocytes. Molecules 2018, 23, 2004. [Google Scholar] [CrossRef]
- Stoddart, M.J. Cell viability assays: Introduction. Methods Mol. Biol. 2011, 740, 1–6. [Google Scholar] [CrossRef]
- Lee, W.; Song, G.; Bae, H. Suppressive Effect of Fraxetin on Adipogenesis and Reactive Oxygen Species Production in 3T3-L1 Cells by Regulating MAPK Signaling Pathways. Antioxidants 2022, 11, 1893. [Google Scholar] [CrossRef]
- Preuss, H.G.; Echard, B.; Polansky, M.M.; Anderson, R. Whole cinnamon and aqueous extracts ameliorate sucrose-induced blood pressure elevations in spontaneously hypertensive rats. J. Am. Coll. Nutr. 2006, 25, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Couturier, K.; Batandier, C.; Awada, M.; Hininger-Favier, I.; Canini, F.; Anderson, R.A.; Leverve, X.; Roussel, A.M. Cinnamon improves insulin sensitivity and alters the body composition in an animal model of the metabolic syndrome. Arch. Biochem. Biophys. 2010, 501, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Knopfholz, J.; Disserol, C.C.; Pierin, A.J.; Schirr, F.L.; Streisky, L.; Takito, L.L.; Massucheto Ledesma, P.; Faria-Neto, J.R.; Olandoski, M.; da Cunha, C.L.; et al. Validation of the friedewald formula in patients with metabolic syndrome. Cholesterol 2014, 2014, 261878. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, T.; Hajihosseini, M.; Moossavi, M.; Hemmati, M.; Ziaee, M. Cardiovascular Risk Factors and Atherogenic Indices in an Iranian Population: Birjand East of Iran. Clin. Med. Insights Cardiol. 2018, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Olamoyegun, M.A.; Oluyombo, R.; Asaolu, S.O. Evaluation of dyslipidemia, lipid ratios, and atherogenic index as cardiovascular risk factors among semi-urban dwellers in Nigeria. Ann. Afr. Med. 2016, 15, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Kim, H.S. Cacao powder supplementation attenuates oxidative stress, cholinergic impairment, and apoptosis in D-galactose-induced aging rat brain. Sci. Rep. 2021, 11, 17914. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Dawson, H.D.; Schoene, N.W.; Polansky, M.M.; Anderson, R.A. Cinnamon polyphenols regulate multiple metabolic pathways involved in insulin signaling and intestinal lipoprotein metabolism of small intestinal enterocytes. Nutrition 2012, 28, 1172–1179. [Google Scholar] [CrossRef]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Farooqi, I.S.; O’Rahilly, S. Leptin: A pivotal regulator of human energy homeostasis. Am. J. Clin. Nutr. 2009, 89, 980S–984S. [Google Scholar] [CrossRef]
- Zhao, S.; Kusminski, C.M.; Scherer, P.E. Adiponectin, Leptin and Cardiovascular Disorders. Circ. Res. 2021, 128, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M. Adiponectin: Role in physiology and pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Villapol, S. Roles of Peroxisome Proliferator-Activated Receptor Gamma on Brain and Peripheral Inflammation. Cell. Mol. Neurobiol. 2018, 38, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Jeong, O.; Kim, H.S. Dietary chokeberry and dried jujube fruit attenuates high-fat and high-fructose diet-induced dyslipidemia and insulin resistance via activation of the IRS-1/PI3K/Akt pathway in C57BL/6 J mice. Nutr. Metab. 2019, 16, 38. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Deng, H.; Wang, S.; El-Seedi, H.; Cao, H.; Chen, L.; Teng, H. Dietary supplementation with casein/cyanidin-3-O-glucoside nanoparticles alters the gut microbiota in high-fat fed C57BL/6 mice. Food Chem. 2023, 412, 135494. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Yu, Z.; Chiang, Y.; Yang, Y.; Chai, T.; Foltz, W.; Lu, H.; Fantus, I.G.; Jin, T. Curcumin Prevents High Fat Diet Induced Insulin Resistance and Obesity via Attenuating Lipogenesis in Liver and Inflammatory Pathway in Adipocytes. PLoS ONE 2012, 7, e28784. [Google Scholar] [CrossRef]
- Reid, B.N.; Ables, G.P.; Otlivanchik, O.A.; Schoiswohl, G.; Zechner, R.; Blaner, W.S.; Goldberg, I.J.; Schwabe, R.F.; Chua, S.C., Jr.; Huang, L.S. Hepatic overexpression of hormone-sensitive lipase and adipose triglyceride lipase promotes fatty acid oxidation, stimulates direct release of free fatty acids, and ameliorates steatosis. J. Biol. Chem. 2008, 283, 13087–13099. [Google Scholar] [CrossRef]
- Foretz, M.; Even, P.C.; Viollet, B. AMPK Activation Reduces Hepatic Lipid Content by Increasing Fat Oxidation In Vivo. Int. J. Mol. Sci. 2018, 19, 2826. [Google Scholar] [CrossRef]
- Fullerton, M.D.; Galic, S.; Marcinko, K.; Sikkema, S.; Pulinilkunnil, T.; Chen, Z.P.; O’Neill, H.M.; Ford, R.J.; Palanivel, R.; O’Brien, M.; et al. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nat. Med. 2013, 19, 1649–1654. [Google Scholar] [CrossRef]
- Zeng, H.L.; Huang, S.L.; Xie, F.C.; Zeng, L.M.; Hu, Y.H.; Leng, Y. Yhhu981, a novel compound, stimulates fatty acid oxidation via the activation of AMPK and ameliorates lipid metabolism disorder in ob/ob mice. Acta Pharmacol. Sin. 2015, 36, 343–352. [Google Scholar] [CrossRef]
- Yu, L.; Zhao, Y.; Zhao, Y. Advances in the pharmacological effects and molecular mechanisms of emodin in the treatment of metabolic diseases. Front. Pharmacol. 2023, 14, 1240820. [Google Scholar] [CrossRef]
- Kim, M.J.; Sim, D.Y.; Lee, H.M.; Lee, H.J.; Kim, S.H. Hypolipogenic Effect of Shikimic Acid Via Inhibition of MID1IP1 and Phosphorylation of AMPK/ACC. Int. J. Mol. Sci. 2019, 20, 582. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.A.; Pinkosky, S.L.; Filippov, S.; Hanselman, J.C.; Cramer, C.T.; Newton, R.S. AMP-activated protein kinase: An emerging drug target to regulate imbalances in lipid and carbohydrate metabolism to treat cardio-metabolic diseases. J. Lipid Res. 2012, 53, 2490–2514. [Google Scholar] [CrossRef] [PubMed]
- Liang, K. Mitochondrial CPT1A: Insights into structure, function, and basis for drug development. Front. Pharmacol. 2023, 14, 1160440. [Google Scholar] [CrossRef] [PubMed]
- Ferré, P.; Phan, F.; Foufelle, F. SREBP-1c and lipogenesis in the liver: An update. Biochem. J. 2021, 478, 3723–3739. [Google Scholar] [CrossRef]
- Ranasinghe, P.; Galappaththy, P.; Constantine, G.R.; Jayawardena, R.; Weeratunga, H.D.; Premakumara, S.; Katulanda, P. Cinnamomum zeylanicum (Ceylon cinnamon) as a potential pharmaceutical agent for type-2 diabetes mellitus: Study protocol for a randomized controlled trial. Trials 2017, 18, 446. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
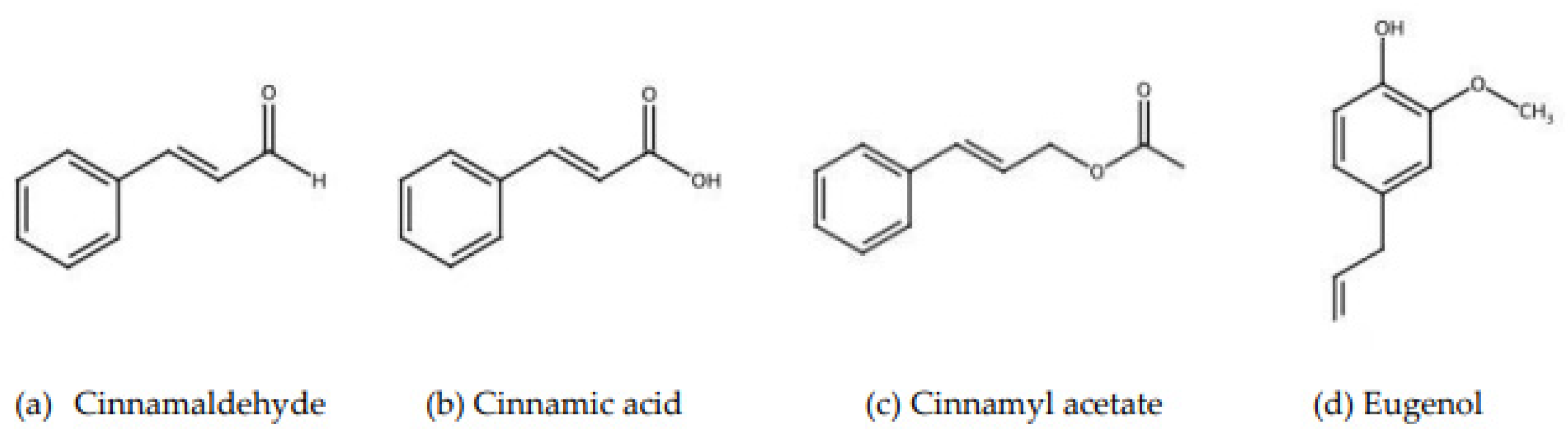
| Part of Cinnamon | Compound | Percentage |
|---|---|---|
| Leaves | Cinnamaldehyde | 1.00–5.00 |
| Eugenol | 70.00–95.00 | |
| Bark | Cinnamaldehyde | 65.00–80.00 |
| Eugenol | 5.00–10.00 | |
| Root bark | Camphor | 60.00 |
| Fruit | Trans-Cinnamyl acetate | 42.00–54.00 |
| Caryophyllene | 9.00–14.00 |
| Target Gene | Primer Sequence |
|---|---|
| Leptin | Forward: 5′-GGA CTT CAT TCC TGG GCT TCA-3′ Reverse: 5′-TGC AGC ACA TTT TGG GAA GG-3′ |
| Adiponectin | Forward: 5′-ACT ACC TGC TAC ATG GCC ACA-3′ Reverse: 5′-AGC AGA TGT GTC CAG ATG TTG-3′ |
| Pparγ | Forward: 5′-CAC CAA CTT CGG AAT CAG CTC-3′ Reverse: 5′-CAA CCA TTG GGT CAG CTC TTG-3′ |
| Ampk | Forward: 5′-ACC TGA GAA CGT CCT GCT TG-3′ Reverse: 5′-GGC CTG CGT ACA ATC TTC CT-3′ |
| Acc | Forward: 5′-ATG GGC GGA ATG GTC TCT TT-3′ Reverse: 5′-TGG GGA CCT TGT CTT CAT CA-3′ |
| Fas | Forward: 5′-TGC ACC CTG ACC CAG AAT AC-3′ Reverse: 5′-CGG CTC AAG GGT TCC ATG TT-3′ |
| Cpt-1 | Forward: 5′-GCC ACT GAT GAA GGA GGG AG-3′ Reverse: 5′-AAT TTG TGG CCC ACC AGG AT-3′ |
| Srebp-1c | Forward: 5′-GCG CTA CCG GTC TTC TAT CA-3′ Reverse: 5′-TGC TGC CAA AAG ACA AGG G-3′ |
| Gapdh | Forward:5′-CCG TGT TCC TAC CCC CAA TG-3′ Reverse: 5′-GTT GCT GTT GAA GTC GCA GG-3′ |
| Cinnamon Extract | |
|---|---|
| Calories (kcal/100 g) | 368.9 |
| Sodium (g/100 g) | 0.1 |
| Carbohydrate (g/100 g) | 90.7 |
| Sugars (g/100 g) | 8.4 |
| Crude fat (g/100 g) | 0.1 |
| Saturated fat (g/100 g) | 0.0 |
| Crude protein (g/100 g) | 1.3 |
| Moisture (g/100 g) | 2.1 |
| Crude ash (g/100 g) | 5.8 |
| Group | ND | NC | HF | HC |
|---|---|---|---|---|
| Macronutrient composition (%) | ||||
| Protein, % of energy | 21.0 | 20.9 | 20.0 | 20.1 |
| Fat, % of energy | 16.0 | 15.9 | 45.0 | 44.6 |
| Carbohydrate, % of energy | 63.0 | 63.2 | 35.0 | 35.3 |
| Energy density, kcal/kg | 3960 | 3996.9 | 4700 | 4689.9 |
| Ingredients (g) | ||||
| Protein | ||||
| Casein, lactic, 30 mesh | 200.0 | 198.0 | 233.1 | 230.7 |
| Cystine, L | 3.0 | 3.0 | 3.5 | 3.5 |
| Protein from cinnamon extract | - | 0.13 | - | 0.13 |
| Carbohydrate | ||||
| Starch, corn | 397.5 | 393.5 | 84.8 | 84.0 |
| Lodex 10 | 132.0 | 130.7 | 116.5 | 115.4 |
| Sucrose, finely granulated | 100.0 | 99.0 | 206.0 | 204.0 |
| Carbohydrate from cinnamon extract | - | 9.9 | - | 9.9 |
| Fiber | ||||
| Solka Floc, FCC200 | 50.0 | 49.5 | 58.3 | 57.7 |
| Fat | ||||
| Soybean oil, USP | 70.0 | 69.3 | 29.1 | 28.8 |
| Lard | - | - | 206.8 | 204.8 |
| Fat from cinnamon extract | - | 0.01 | - | 0.01 |
| Mineral | ||||
| S10022G | 35.0 | 34.7 | - | - |
| S10026B | - | - | 58.3 | 57.7 |
| Vitamin | ||||
| V10037 | 10.0 | 9.9 | - | - |
| V10001C | - | - | 1.2 | 1.2 |
| Choline bitartrate | 2.5 | 2.5 | 2.3 | 2.3 |
| Dye | ||||
| Red FD&C #40 | - | - | 0.06 | 0.06 |
| Alum. lake 35-42% | ||||
| Total (g) | 1000.0 | 1000.14 | 999.96 | 1000.14 |
| Group | ND | NC | HF | HC |
|---|---|---|---|---|
| Initial body weight (g) | 24.00 ± 1.68 | 24.07 ± 1.24 | 25.11 ± 1.58 | 25.26 ± 1.07 |
| Final body weight (g) | 33.06 ± 2.55 *** | 28.93 ± 1.40 *** | 40.23 ± 4.19 | 37.37 ± 2.14 |
| Body weight gain (g) | 9.06 ± 2.09 **** | 4.86 ± 1.02 *** | 15.12 ± 3.15 | 12.11 ± 1.25 * |
| Food intake (g/day) | 3.46 ± 0.41 | 3.88 ± 1.07 | 3.41 ± 1.02 | 3.89 ± 0.99 |
| FER (%) | 6.11 ± 0.73 **** | 4.15 ± 0.45 * | 13.72 ± 0.67 | 10.46 ± 0.81 ** |
| Abdominal fat (g) | 0.20 ± 0.07 **** | 0.18 ± 0.07 ns | 0.46 ± 0.08 | 0.27 ± 0.11 **** |
| Epididymal fat (g) | 0.66 ± 0.19 **** | 0.52 ± 0.19 | 2.01 ± 0.44 | 1.83 ± 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.; Ahn, S.; Zhou, X.; Lim, Y.J.; Hong, S.; Kim, H.-S. Effects of Cinnamon (Cinnamomum zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet. Nutrients 2023, 15, 5110. https://doi.org/10.3390/nu15245110
Oh J, Ahn S, Zhou X, Lim YJ, Hong S, Kim H-S. Effects of Cinnamon (Cinnamomum zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet. Nutrients. 2023; 15(24):5110. https://doi.org/10.3390/nu15245110
Chicago/Turabian StyleOh, Joohee, SongHee Ahn, Xiangqin Zhou, Yu Jin Lim, Sookyeong Hong, and Hyun-Sook Kim. 2023. "Effects of Cinnamon (Cinnamomum zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet" Nutrients 15, no. 24: 5110. https://doi.org/10.3390/nu15245110
APA StyleOh, J., Ahn, S., Zhou, X., Lim, Y. J., Hong, S., & Kim, H.-S. (2023). Effects of Cinnamon (Cinnamomum zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet. Nutrients, 15(24), 5110. https://doi.org/10.3390/nu15245110