


Anti-Menopausal Effect of Soybean Germ Extract and Lactobacillus gasseri in the Ovariectomized Rat Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of S30
2.2. Preparation of Bacterial Species Culture
2.3. Assay for β-Glucosidase Enzyme from Probiotics in S30
2.4. Cell Culture
2.5. Real-Time Reverse-Transcription PCR Assay
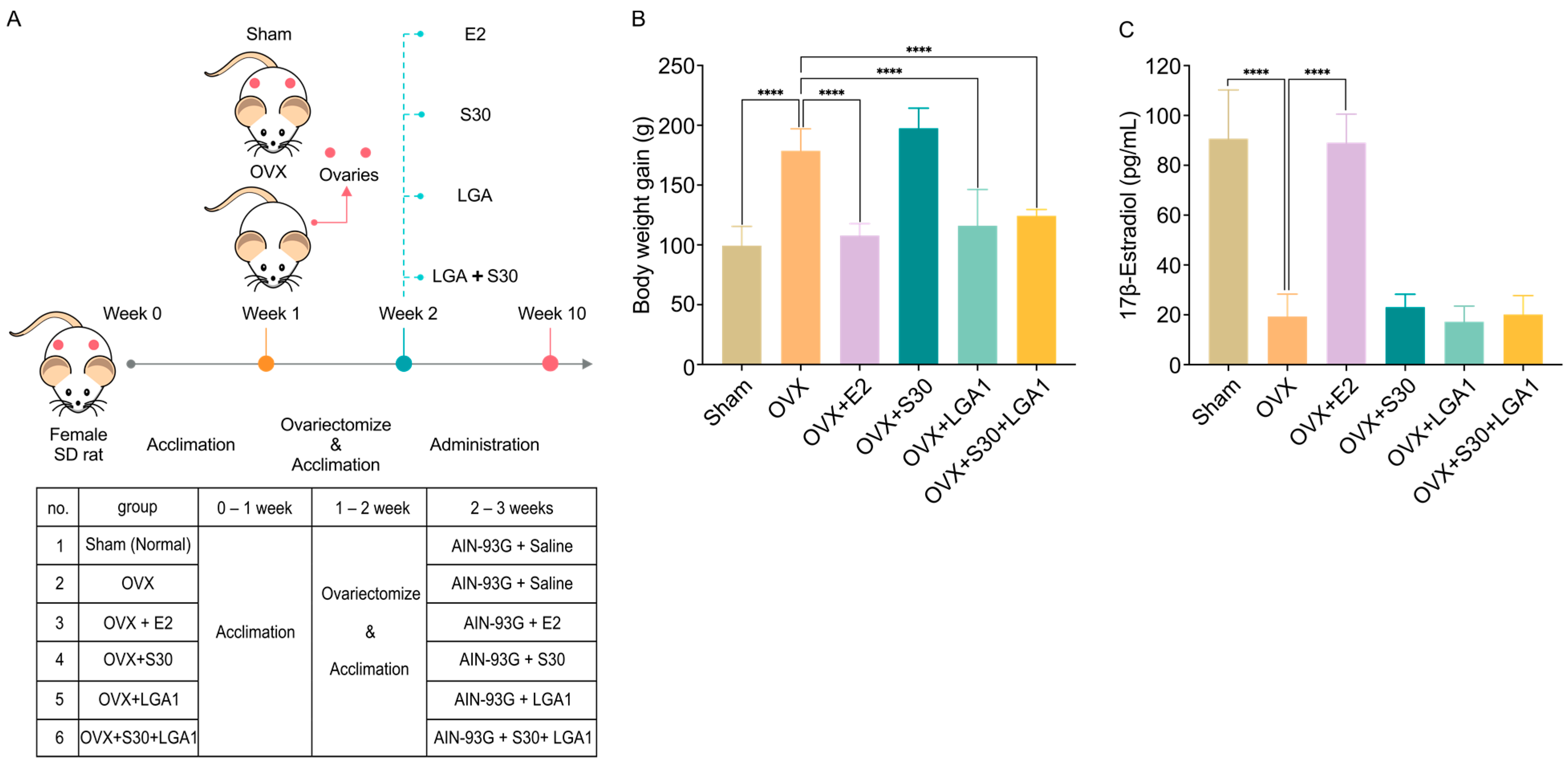
2.6. Animal Models
2.7. Micro-Computerized Tomography Analysis
2.8. Measurement of Abdominal Fat in Live Animals
2.9. Analysis of the Uterine Morphometric Parameters
2.10. Serum Biochemical Marker Analysis
2.11. Statistical Analysis
3. Results
3.1. Screening of High β-Glucosidase Producing LAB
3.2. Effect of S30+LGA1 on Estrogen Markers in MCF-7 Cells
3.3. Effect of S30+LGA1 on Bone-Related Markers in MG-63 Cells
3.4. Effect of S30+LGA1 on the Body Weight of OVX Rats
3.5. Effect of S30+LGA1 on the Abdominal Fat Volume in OVX Rats
3.6. Effect of S30+LGA1 on Uterine Weight and Tissues in OVX Rats
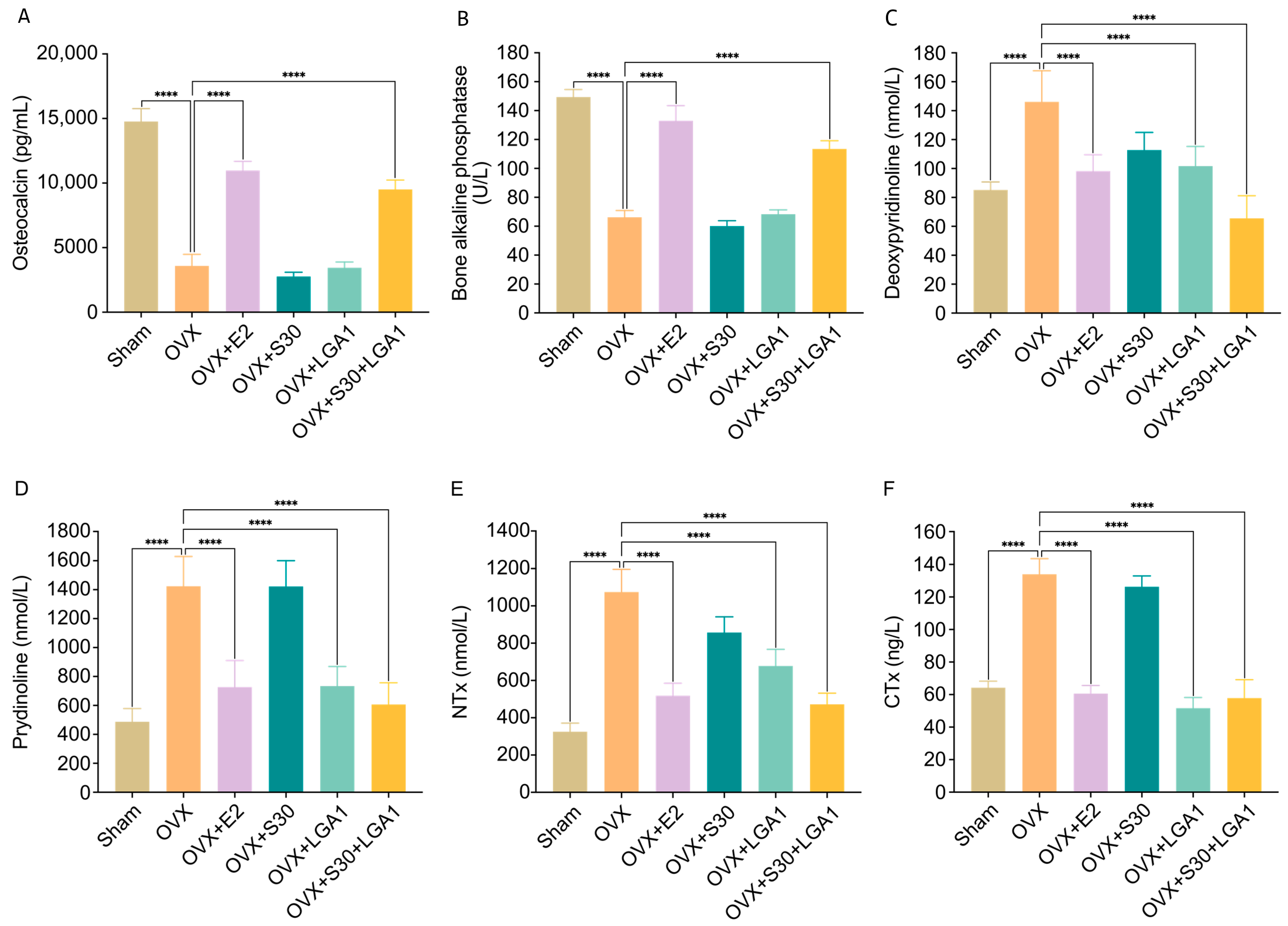
3.7. Effect of S30+LGA1 on Bone Losses in OVX Rats
3.8. Effect of S30+LGA1 on the Bone-Related Biochemical Markers in OVX Rats
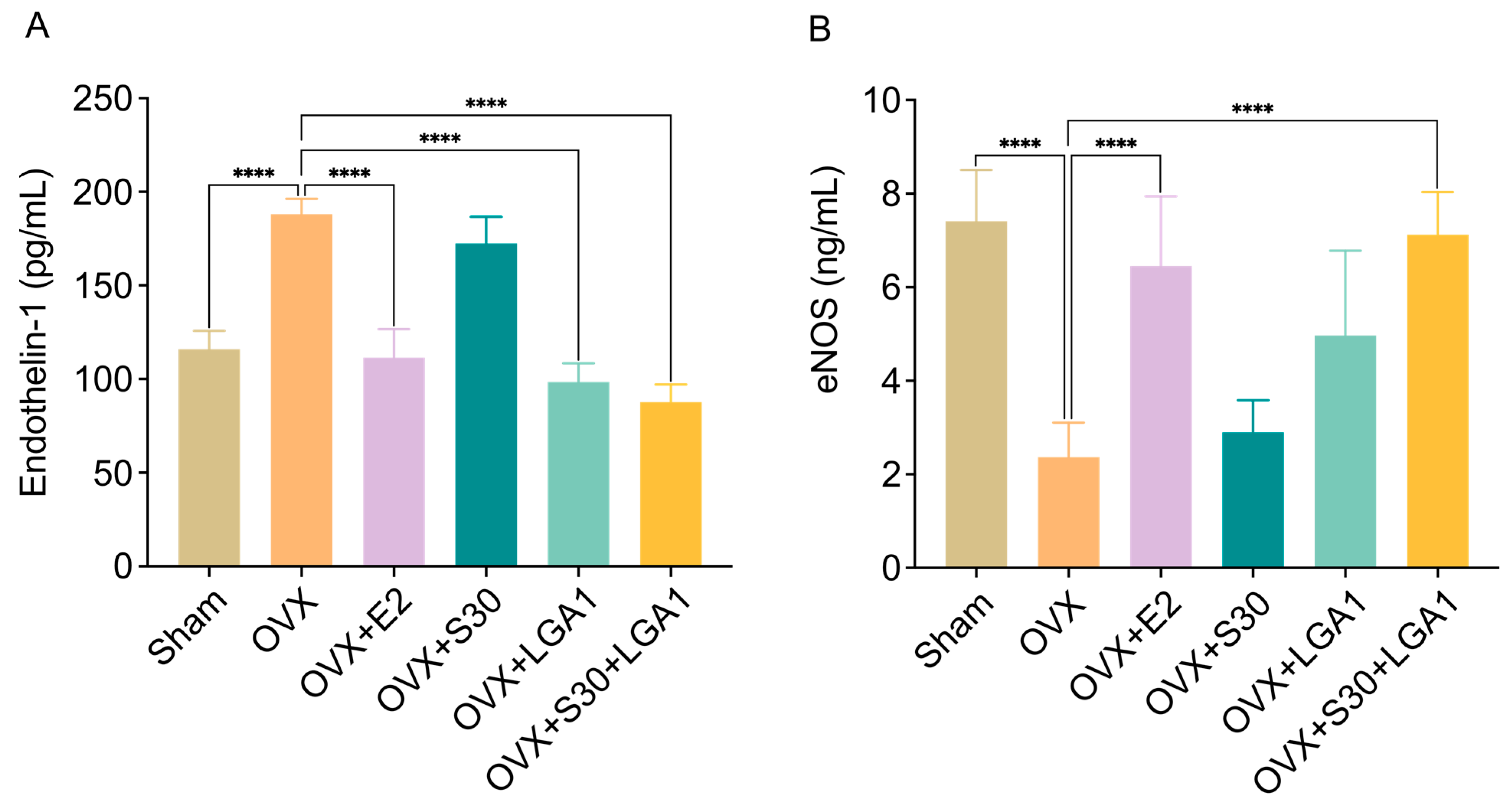
3.9. Effect of S30+LGA1 on Vascular Homeostasis in OVX Rats
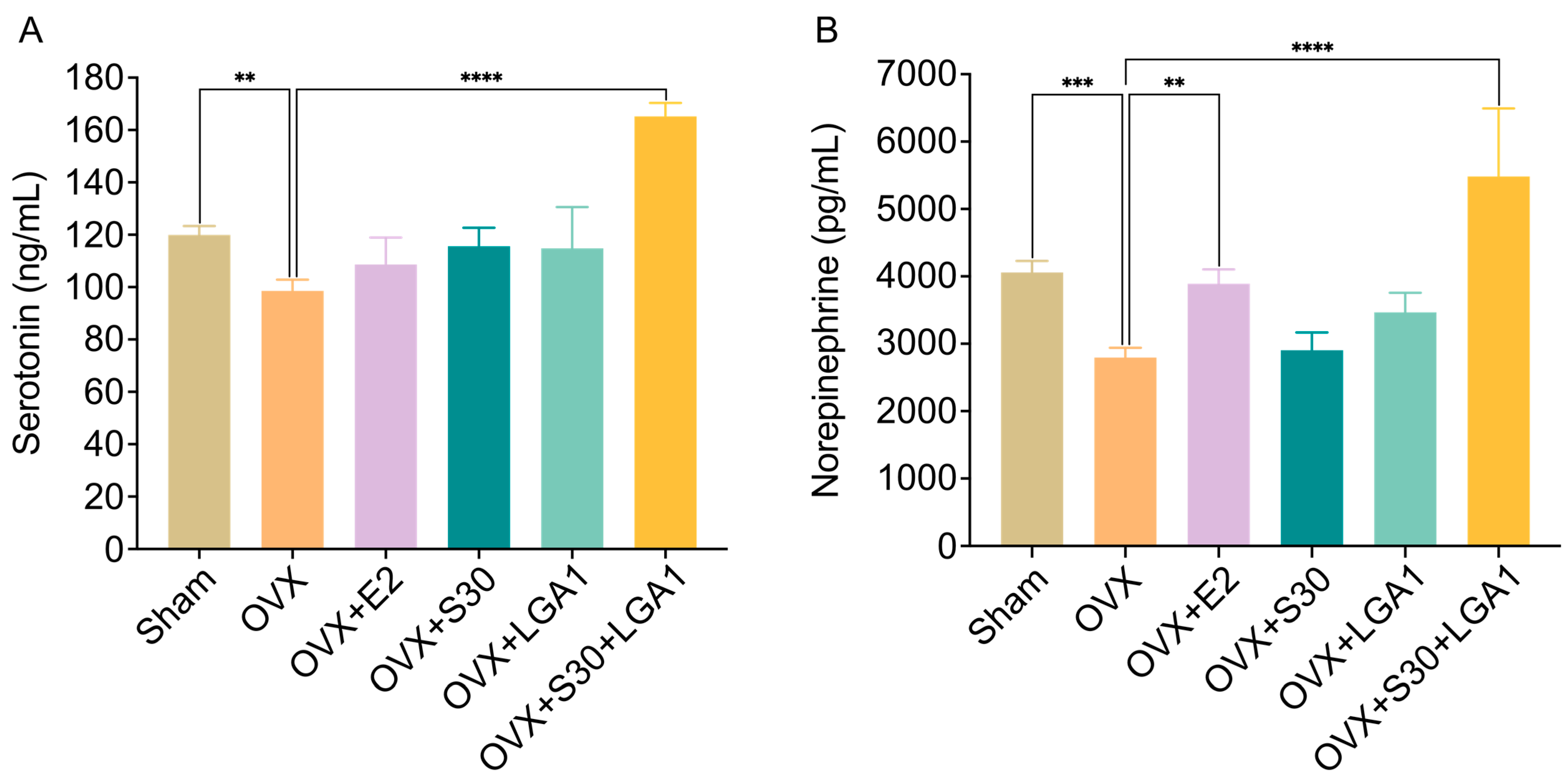
3.10. Effect of S30+LGA1 on Serotonin and Norepinephrine Levels in OVX Rats
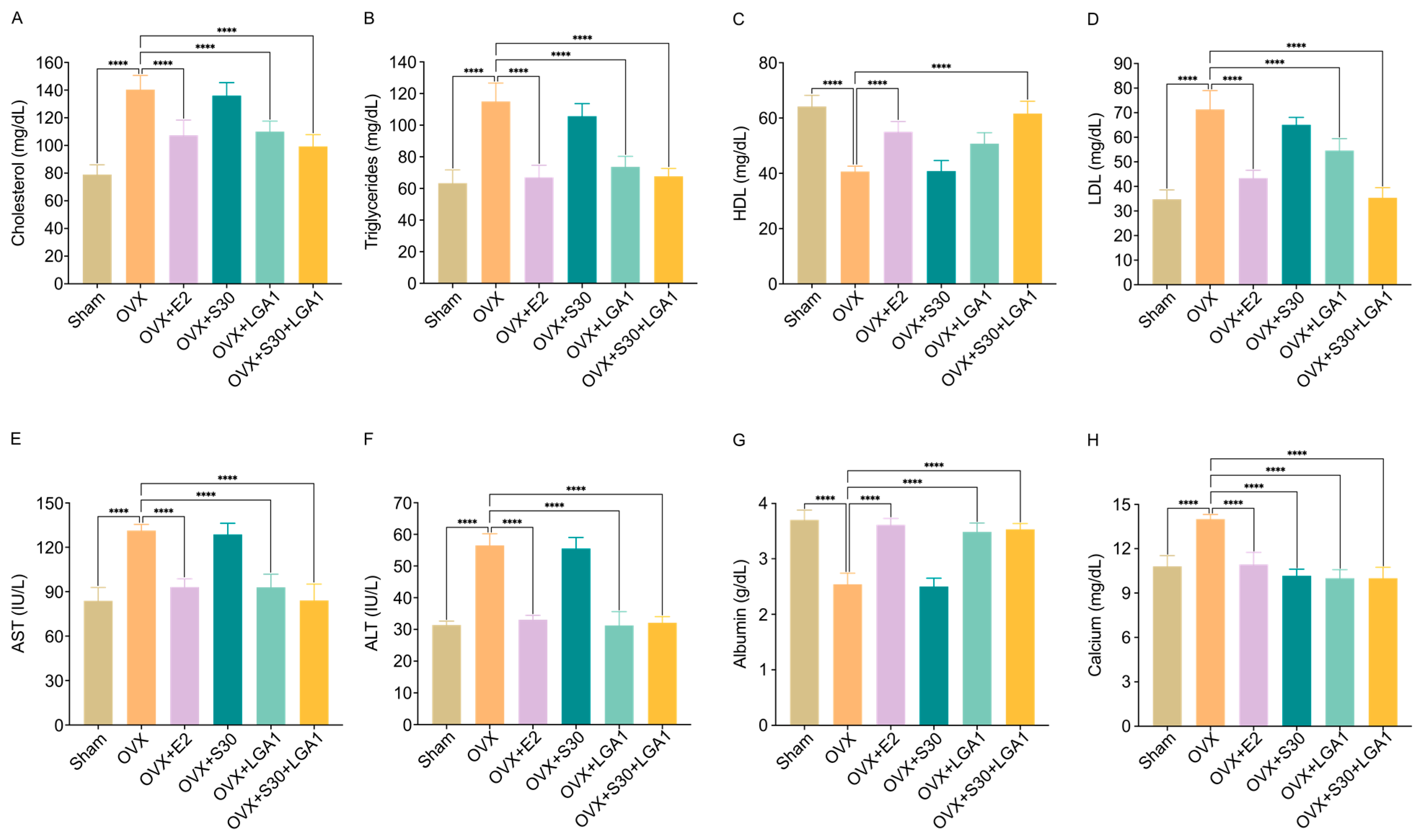
3.11. Effect of S30+LGA1 on Lipid Profile and Liver Enzymes, Albumin, and Calcium Levels in OVX Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mulhall, S.; Andel, R.; Anstey, K.J. Variation in symptoms of depression and anxiety in midlife women by menopausal status. Maturitas 2018, 108, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.R.; Pillai, A.J.; Nair, N. Cardiovascular changes in menopause. Curr. Cardiol. Rev. 2021, 17, e230421187681. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Sohail, R.; Jaffer, S.R.; Siddique, S.; Kaya, B.; Atowoju, I.; Imran, A.; Wright, W.; Pamulapati, S.; Choudhry, F.; et al. The role of estrogen therapy as a protective factor for Alzheimer’s disease and dementia in postmenopausal women: A comprehensive review of the literature. Cureus 2023, 15, e43053. [Google Scholar] [CrossRef] [PubMed]
- Kołodyńska, G.; Zalewski, M.; Rożek-Piechura, K. Urinary incontinence in postmenopausal women—Causes, symptoms, treatment. Prz. Menopauzalny 2019, 18, 46–50. [Google Scholar] [CrossRef]
- Lobo, R.A.; Gompel, A. Management of menopause: A view towards prevention. Lancet Diabetes Endocrinol. 2022, 10, 457–470. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, J.; Zhu, S.; He, M.; Ma, S.; Jia, Q.; Sun, Q.; Song, L.; Wang, Y.; Duan, L. Berberine ameliorates ovariectomy-induced anxiety-like behaviors by enrichment in equol generating gut microbiota. Pharmacol. Res. 2021, 165, 105439. [Google Scholar] [CrossRef]
- Nelson, H.D. Commonly used types of postmenopausal estrogen for treatment of hot flashes: Scientific review. JAMA 2004, 291, 1610–1620. [Google Scholar] [CrossRef]
- Flores, V.A.; Pal, L.; Manson, J.E. Hormone therapy in menopause: Concepts, controversies, and approach to treatment. Endocr. Rev. 2021, 42, 720–752. [Google Scholar] [CrossRef]
- Hashemzadeh, M.; Haseefa, F.; Peyton, L.; Park, S.; Movahed, M.R. The effects of estrogen and hormone replacement therapy on platelet activity: A review. Am. J. Blood Res. 2022, 12, 33–42. [Google Scholar]
- Chen, L.R.; Chen, K.H. Utilization of isoflavones in soybeans for women with menopausal syndrome: An overview. Int. J. Mol. Sci. 2021, 22, 3212. [Google Scholar] [CrossRef]
- Rietjens, I.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Boutas, I.; Kontogeorgi, A.; Dimitrakakis, C.; Kalantaridou, S.N. Soy isoflavones and breast cancer risk: A meta-analysis. In Vivo 2022, 36, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Ziaei, S.; Halaby, R. Dietary isoflavones and breast cancer risk. Medicines. 2017, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Coward, L.; Smith, M.; Kirk, M.; Barnes, S. Chemical modification of isoflavones in soyfoods during cooking and processing. Am. J. Clin. Nutr. 1998, 68, 1486S–1491S. [Google Scholar] [CrossRef] [PubMed]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented soy products and their potential health benefits: A review. Microorganisms 2022, 10, 1606. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kikuchi, M.; Piskula, M.K.; Kubota, Y. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. Sci. 2000, 130, 1695–1699. [Google Scholar] [CrossRef]
- Guo-zhen, X.; Li-li, H.; Shui-han, Z.; Zhou-jin, T. Advances in metabolism of glycosides by gut microbiota. Nat. Prod. Res. Dev. 2022, 34, 1261–1271. [Google Scholar] [CrossRef]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A bacterial metabolite from the daidzein isoflavone and its presumed beneficial health effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef]
- Setchell, K.D.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [CrossRef]
- Bolca, S.; Van de Wiele, T.; Possemiers, S. Gut metabotypes govern health effects of dietary polyphenols. Curr. Opin. Biotechnol. 2013, 24, 220–225. [Google Scholar] [CrossRef]
- Frankenfeld, C.L. Archives of medical science. Mol. Nutr. Food Res. 2017, 61, 1500900. [Google Scholar] [CrossRef]
- Leonard, L.M.; Choi, M.S.; Cross, T.-W.L. Maximizing the estrogenic potential of soy isoflavones through the gut microbiome: Implication for cardiometabolic health in postmenopausal women. Nutrients 2022, 14, 553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fujiyoshi, A.; Kadota, A.; Kondo, K.; Torii, S.; Okami, Y.; Hisamatsu, T.; Yano, Y.; Barinas-Mitchell, E.; Magnani, J.; et al. Cross-sectional association of equol producing status with aortic calcification in Japanese men aged 40–79 years. Sci. Rep. 2022, 12, 20114. [Google Scholar] [CrossRef]
- Delgado, S.; Guadamuro, L.; Flórez, A.B.; Vázquez, L.; Mayo, B. Fermentation of commercial soy beverages with lactobacilli and bifidobacteria strains featuring high β-glucosidase activity. Innov. Food Sci. Emerg. Technol. 2019, 51, 148–155. [Google Scholar] [CrossRef]
- Champagne, C.P.; Tompkins, T.A.; Buckley, N.D.; Green-Johnson, J.M. Effect of fermentation by pure and mixed cultures of Streptococcus thermophilus and Lactobacillus helveticus on isoflavone and B-vitamin content of a fermented soy beverage. Food Microbiol. 2010, 27, 968–972. [Google Scholar] [CrossRef]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zheng, T.; Hui, H.; Xie, G. Soybean isoflavones modulate gut microbiota to benefit the health weight and metabolism. Front. Cell. Infect. Microbiol. 2022, 12, 1004765. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef]
- Landete, J.M.; Arqués, J.; Medina, M.; Gaya, P.; de Las Rivas, B.; Muñoz, R. Bioactivation of phytoestrogens: Intestinal bacteria and Health. Crit. Rev. Food Sci. Nutr. 2016, 56, 1826–1843. [Google Scholar] [CrossRef]
- Koppel, N.; Maini Rekdal, V.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, eaag2770. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, Á.; Medina, M.; Landete, J.M. Isoflavone metabolism by a collection of lactic acid bacteria and bifidobacteria with biotechnological interest. Int. J. Food Sci. Nutr. 2016, 67, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Kim, G.M.; Lee, K.W.; Choi, I.D.; Kwon, G.-H.; Park, J.-Y.; Jeong, S.-J.; Kim, J.-S.; Kim, J.H. Conversion of isoflavone glucosides to aglycones in soymilk by fermentation with lactic acid bacteria. J. Food Sci. 2007, 72, M39–M44. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, Á.; Landete, J.M. Transformation of plant isoflavones into bioactive isoflavones by lactic acid bacteria and bifidobacteria. J. Funct. Foods 2017, 39, 198–205. [Google Scholar] [CrossRef]
- Scalabrini, P.; Rossi, M.; Spettoli, P.; Matteuzzi, D. Characterization of Bifidobacterium strains for use in soymilk fermentation. Int. J. Food Microbiol. 1998, 39, 213–219. [Google Scholar] [CrossRef]
- Kim, J.; Petz, L.N.; Ziegler, Y.S.; Wood, J.R.; Potthoff, S.J.; Nardulli, A.M. Regulation of the estrogen-responsive pS2 gene in MCF-7 human breast cancer cells. J. Steroid Biochem. Mol. Biol. 2000, 74, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Dalal, H.; Dahlgren, M.; Gladchuk, S.; Brueffer, C.; Gruvberger-Saal, S.K.; Saal, L.H. Clinical associations of ESR2 (estrogen receptor beta) expression across thousands of primary breast tumors. Sci. Rep. 2022, 12, 4696. [Google Scholar] [CrossRef] [PubMed]
- Syed, F.; Khosla, S. Mechanisms of sex steroid effects on bone. Biochem. Biophys. Res. Commun. 2005, 328, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, J.; Wiesnerova, L.; Chocholata, P.; Kulda, V.; Landsmann, L.; Cedikova, M.; Kripnerova, M.; Eberlova, L.; Babuska, V. Human cells with osteogenic potential in bone tissue research. Biomed. Eng. Online 2023, 22, 33. [Google Scholar] [CrossRef]
- Szulc, P.; Chapuy, M.; Meunier, P.; Delmas, P. Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture in elderly women. J. Clin. Investig. 1993, 91, 1769–1774. [Google Scholar] [CrossRef]
- Cairns, J.R.; Price, P.A. Direct demonstration that the vitamin k-dependent bone Gla protein is incompletely γ-carboxylated in humans. J. Bone Miner. Res. 1994, 9, 1989–1997. [Google Scholar] [CrossRef]
- Gundberg, C.M.; Weinstein, R.S. Multiple immunoreactive forms of osteocalcin in uremic serum. J. Clin. Investig. 1986, 77, 1762–1767. [Google Scholar] [CrossRef]
- Taylor, A.K.; Linkhart, S.; Mohan, S.; Christenson, R.A.; Singer, F.R.; Baylink, D.J. Multiple osteocalcin fragments in human urine and serum as detected by a midmolecule osteocalcin radioimmunoassay. J. Clin. Endocrinol. Metab. 1990, 70, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, C.; Qvist, P.; Bjarnason, N.; Christiansen, C. Measurement of a more stable region of osteocalcin in serum by ELISA with two monoclonal antibodies. Clin. Chem. 1995, 41, 1439–1445. [Google Scholar] [CrossRef]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Lüthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.H.; Lane, J.M.; Urist, M.R.; Lyons, K.M.; Lieberman, J.R. Bone morphogenetic protein-2: Biology and applications. Clin. Orthop. Relat. Res. 1996, 324, 39–46. [Google Scholar] [CrossRef]
- Miljkovic, N.D.; Cooper, G.M.; Marra, K.G. Chondrogenesis, bone morphogenetic protein-4 and mesenchymal stem cells. Osteoarthr. Cartil. 2008, 16, 1121–1130. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, S.; Lovisa, S.; Ambrose, C.G.; McAndrews, K.M.; Sugimoto, H.; Kalluri, R. Type-I collagen produced by distinct fibroblast lineages reveals specific function during embryogenesis and osteogenesis Imperfecta. Nat. Commun. 2021, 12, 7199. [Google Scholar] [CrossRef]
- Saraç, F.; Saygılı, F. Causes of high bone alkaline phosphatase. Biotechnol. Biotechnol. Equip. 2007, 21, 194–197. [Google Scholar] [CrossRef]
- Cooper, C.; Gehlbach, S.H.; Lindsay, R. Prevention and Treatment of Osteoporosis in the High-Risk Patient: A Clinician’s Guide; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar] [CrossRef]
- Keen, R. Osteoporosis: Strategies for prevention and management. Best Pract. Res. Clin. Rheumatol. 2007, 21, 109–122. [Google Scholar] [CrossRef]
- Lobo, R.A. Hormone-replacement therapy: Current thinking. Nat. Rev. Endocrinol. 2017, 13, 220–231. [Google Scholar] [CrossRef]
- Writing Group for the Women’s Health Initiative Investigators. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: Principal results from the women’s health initiative randomized controlled trial. JAMA 2002, 288, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Maraka, S.; Kennel, K.A. Bisphosphonates for the prevention and treatment of osteoporosis. BMJ 2015, 351, h3783. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Fujimoto, J.; Hori, M.; Ichigo, S.; Morishita, S.; Tamaya, T. Estrogen induces expression of c-fos and c-jun via activation of protein kinase C in an endometrial cancer cell line and fibroblasts derived from human uterine endometrium. Gynecol. Endocrinol. 1996, 10, 109–118. [Google Scholar] [CrossRef]
- Desmawati, D.; Sulastri, D. Phytoestrogens and their health effect. Open Access Maced. J. Med. Sci. 2019, 7, 495. [Google Scholar] [CrossRef]
- Kenneth, D.R.S. Phytoestrogens: The biochemistry, physiology, and implications for human health of soy isoflavones. Am. J. Clin. Nutr. 1998, 68, 1333S–1346S. [Google Scholar] [CrossRef] [PubMed]
- Huntley, A.L.; Ernst, E. A systematic review of herbal medicinal products for the treatment of menopausal symptoms. Menopause 2003, 10, 465–476. [Google Scholar] [CrossRef]
- Posadzki, P.; Lee, M.S.; Moon, T.W.; Choi, T.Y.; Park, T.Y.; Ernst, E. Prevalence of complementary and alternative medicine (CAM) use by menopausal women: A systematic review of surveys. Maturitas 2013, 75, 34–43. [Google Scholar] [CrossRef]
- Messina, M.; Gleason, C. Evaluation of the potential antidepressant effects of soybean isoflavones. Menopause 2016, 23, 1348–1360. [Google Scholar] [CrossRef]
- Bhathena, S.J.; Velasquez, M.T. Beneficial role of dietary phytoestrogens in obesity and diabetes. Am. J. Clin. Nutr. 2002, 76, 1191–1201. [Google Scholar] [CrossRef]
- Gómez-Zorita, S.; González-Arceo, M.; Fernández-Quintela, A.; Eseberri, I.; Trepiana, J.; Portillo, M.P. Scientific evidence supporting the beneficial effects of isoflavones on human health. Nutrients 2020, 12, 3853. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]
- Riggs, B.L.; Khosla, S.; Melton, L.J., III. A unitary model for involutional osteoporosis: Estrogen deficiency causes both type I and Type II osteoporosis in postmenopausal women and contributes to bone loss in aging men. J. Bone Miner. Res. 1998, 13, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Bex, M.; Vanderschueren, D.; Boonen, S. Estrogens are essential for male pubertal periosteal bone expansion. J. Clin. Endocrinol. Metab. 2004, 89, 6025–6029. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Ohnishi-Kameyama, M.; Shinohara, K. Lactobacillus gasseri: Effects on mouse intestinal flora enzyme activity and isoflavonoids in the caecum and plasma. Br. J. Nutr. 2004, 92, 771–776. [Google Scholar] [CrossRef]
- Awa, W.L.; Fach, E.; Krakow, D.; Welp, R.; Kunder, J.; Voll, A.; Zeyfang, A.; Wagner, C.; Schütt, M.; Boehm, B.; et al. Type 2 diabetes from pediatric to geriatric age: Analysis of gender and obesity among 120,183 patients from the German/Austrian DPV database. Eur. J. Endocrinol. 2012, 167, 245–254. [Google Scholar] [CrossRef]
- Wietlisbach, V.; Marques-Vidal, P.; Kuulasmaa, K.; Karvanen, J.; Paccaud, F. The relation of body mass index and abdominal adiposity with dyslipidemia in 27 general populations of the WHO MONICA Project. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 432–442. [Google Scholar] [CrossRef]
- Dang, Z.C.; Van Bezooijen, R.L.; Karperien, M.; Papapoulos, S.E.; Löwik, C.W.G.M. Exposure of KS483 cells to estrogen enhances osteogenesis and inhibits adipogenesis. J. Bone Miner. Res. 2002, 17, 394–405. [Google Scholar] [CrossRef]
- Burch, K.E.; McCracken, K.; Buck, D.J.; Davis, R.L.; Sloan, D.K.; Curtis, K.S. Relationship between circulating metabolic hormones and their central receptors during ovariectomy-induced weight gain in rats. Front. Physiol. 2022, 12, 800266. [Google Scholar] [CrossRef]
- Nogowski, L.; Maćkowiak, P.; Kandulska, K.; Szkudelski, T.; Nowak, K.W. Genistein-Induced changes in lipid metabolism of ovariectomized rats. Ann. Nutr. Metab. 1999, 42, 360–366. [Google Scholar] [CrossRef]
- Choi, J.S.; Koh, I.-U.; Song, J. Genistein reduced insulin resistance index through modulating lipid metabolism in ovariectomized rats. Nutr. Res. 2012, 32, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Cross, T.-W.L.; Zidon, T.M.; Welly, R.J.; Park, Y.-M.; Britton, S.L.; Koch, L.G.; Rottinghaus, G.E.; de Godoy, M.R.C.; Padilla, J.; Swanson, K.S.; et al. Soy improves cardiometabolic health and cecal microbiota in female low-fit rats. Sci. Rep. 2017, 7, 9261. [Google Scholar] [CrossRef]
- Torre-Villalvazo, I.; Tovar, A.R.; Ramos-Barragán, V.E.; Cerbón-Cervantes, M.A.; Torres, N. Soy protein ameliorates metabolic abnormalities in liver and adipose tissue of rats fed a high fat diet. J. Nutr. Sci. 2008, 138, 462–468. [Google Scholar] [CrossRef]
- Li, L.-l.; Yang, Y.; Ma, C.-m.; Li, X.-m.; Bian, X.; Fu, Y.; Ren, L.-k.; Wang, R.-m.; Shi, Y.-g.; Zhang, N. Effects of soybean isoflavone aglycone on osteoporosis in ovariectomized rats. Front. Nutr. 2023, 10, 1122045. [Google Scholar] [CrossRef]
- Kim, J.; Cha, S.; Lee, M.Y.; Hwang, Y.J.; Yang, E.; Ryou, C.; Jung, H.-I.; Cheon, Y.-P. Chronic low-dose nonylphenol or di-(2-ethylhexyl) phthalate has a different estrogen-like response in mouse uterus. Dev. Reprod. 2018, 22, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Sang, C.; Zhang, Y.; Chen, F.; Huang, P.; Qi, J.; Wang, P.; Zhou, Q.; Kang, H.; Cao, X.; Guo, L. Tumor necrosis factor alpha suppresses osteogenic differentiation of MSCs by inhibiting semaphorin 3B via Wnt/β-catenin signaling in estrogen-deficiency induced osteoporosis. Bone 2016, 84, 78–87. [Google Scholar] [CrossRef]
- Devareddy, L.; Khalil, D.A.; Smith, B.J.; Lucas, E.A.; Soung, D.Y.; Marlow, D.D.; Arjmandi, B.H. Soy moderately improves microstructural properties without affecting bone mass in an ovariectomized rat model of osteoporosis. Bone 2006, 38, 686–693. [Google Scholar] [CrossRef]
- Ding, H.; Chang, W.; Yao, J.; Liu, F.; Zeng, Q.; He, Z. Anti-osteoporosis effect of Notopterygium incisum Ting ex HT Chang extract in rats. Trop. J. Pharm. Res. 2019, 18, 2051–2055. [Google Scholar] [CrossRef]
- Chiang, S.-S.; Pan, T.-M. Antiosteoporotic effects of Lactobacillus-fermented soy skim milk on bone mineral density and the microstructure of femoral bone in ovariectomized mice. J. Agric. Food Chem. 2011, 59, 7734–7742. [Google Scholar] [CrossRef]
- Bouxsein, M.L.; Myers, K.S.; Shultz, K.L.; Donahue, L.R.; Rosen, C.J.; Beamer, W.G. Ovariectomy-induced bone loss varies among inbred strains of mice. J. Bone Miner. Res. 2005, 20, 1085–1092. [Google Scholar] [CrossRef]
- Mercuro, G.; Zoncu, S.; Pilia, I.; Lao, A.; Benedetto Melis, G.; Cherchi, A. Effects of acute administration of transdermal estrogen on postmenopausal women with systemic hypertension. Am. J. Cardiol. 1997, 80, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Delgado, J.L.; Díaz, J.; Quesada, T.; Teruel, M.J.G.; Llanos, M.C.; Carbonell, L.F. 17β-Estradiol prevents oxidative stress and decreases blood pressure in ovariectomized rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1599–R1605. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, K.; Nakak, T.; Marumo, T.; Suzuki, H.; Kato, R.; Saruta, T. Up-regulation of nitric oxide synthase by estradiol in human aorticendothelial cells. FEBS Lett. 1995, 360, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Simoncini, T.; Fornari, L.; Mannella, P.; Varone, G.; Caruso, A.; Garibaldi, S.; Genazzani, A.R. Differential estrogen signaling in endothelial cells upon pulsed or continuous administration. Maturitas 2005, 50, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, R.J.; Walker, B.R.; Kanagy, N.L. 17β-estradiol increases nitric oxide-dependent dilation in rat pulmonary arteries and thoracic aorta. Am. J. Physiol. Lung Cell Mol. Physiol. 2001, 280, L555–L564. [Google Scholar] [CrossRef]
- Rosselli, M.; Imthurm, B.; Macas, E.; Keller, P.J.; Dubey, R.K. Circulating nitrite/nitrate levels increase with follicular development: Indirect evidence for estradiol-mediated NO release. Biochem. Biophys. Res. Commun. 1994, 202, 1543–1552. [Google Scholar] [CrossRef]
- Feldstein, C.; Romero, C. Role of endothelins in hypertension. Am. J. Ther. 2007, 14, 147–153. [Google Scholar] [CrossRef]
- Ergul, A.; Shoemaker, K.; Puett, D.; Tackett, R.L. Gender differences in the expression of endothelin receptors in human saphenous veins in vitro. J. Pharmacol. Exp. Ther. 1998, 285, 511–517. [Google Scholar]
- Kobayashi, T.; Tamura, M.; Hayashi, M.; Katsuura, Y.; Tanabe, H.; Ohta, T.; Komoriya, K. Elevation of tail skin temperature in ovariectomized rats in relation to menopausal hot flushes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R863–R869. [Google Scholar] [CrossRef]
- Blake, C.; Fabick, K.M.; Setchell, K.D.R.; Lund, T.D.; Lephart, E.D. Neuromodulation by soy diets or equol: Anti-depressive & anti-obesity-like influences, age- & hormone-dependent effects. BMC Neurosci. 2011, 12, 28. [Google Scholar] [CrossRef]
- El-Khatib, Y.A.; Sayed, R.H.; Sallam, N.A.; Zaki, H.F.; Khattab, M.M. 17β-Estradiol augments the neuroprotective effect of agomelatine in depressive- and anxiety-like behaviors in ovariectomized rats. Psychopharmacology 2020, 237, 2873–2886. [Google Scholar] [CrossRef] [PubMed]
- Saied, N.M.; Georgy, G.S.; Hussien, R.M.; Hassan, W.A. Neuromodulatory effect of curcumin on catecholamine systems and inflammatory cytokines in ovariectomized female rats. Clin. Exp. Pharmacol. 2021, 48, 337–346. [Google Scholar] [CrossRef]
- Ko, S.H.; Kim, H.S. Menopause-associated lipid metabolic disorders and foods beneficial for postmenopausal women. Nutrients. 2020, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Banerjee, R.; Rial, B.; Arlt, W.; Adiels, M.; Boren, J.; Marinou, K.; Fisher, C.; Mostad, I.L.; Stratton, I.M.; et al. Menopausal status and abdominal obesity are significant determinants of hepatic lipid metabolism in women. J. Am. Heart Assoc. 2015, 4, e002258. [Google Scholar] [CrossRef] [PubMed]
- Ryczkowska, K.; Adach, W.; Janikowski, K.; Banach, M.; Bielecka-Dabrowa, A. Menopause and women’s cardiovascular health: Is it really an obvious relationship? Arch. Med. Sci. 2023, 19, 458–466. [Google Scholar] [CrossRef]
- Kim, H.R.; Han, M.A. Association between serum liver enzymes and metabolic syndrome in Korean adults. Int. J. Environ. Res. Public Health 2018, 15, 1658. [Google Scholar] [CrossRef]
- Ko, S.-H.; Baeg, M.K.; Han, K.-D.; Ko, S.-H.; Ahn, Y.-B. Increased liver markers are associated with higher risk of type 2 diabetes. World J. Gastroenterol. 2015, 21, 7478–7487. [Google Scholar] [CrossRef]
- Bern, M.; Sand, K.M.K.; Nilsen, J.; Sandlie, I.; Andersen, J.T. The role of albumin receptors in regulation of albumin homeostasis: Implications for drug delivery. J. Control. Release 2015, 211, 144–162. [Google Scholar] [CrossRef]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.; et al. Calcium intake in bone health: A focus on calcium-rich mineral waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Lim, T.-J.; Yun, E.J.; Kim, K.H.; Lim, S. Anti-Menopausal Effect of Soybean Germ Extract and Lactobacillus gasseri in the Ovariectomized Rat Model. Nutrients 2023, 15, 4485. https://doi.org/10.3390/nu15204485
Lee S-H, Lim T-J, Yun EJ, Kim KH, Lim S. Anti-Menopausal Effect of Soybean Germ Extract and Lactobacillus gasseri in the Ovariectomized Rat Model. Nutrients. 2023; 15(20):4485. https://doi.org/10.3390/nu15204485
Chicago/Turabian StyleLee, Sun-Hee, Tae-Joong Lim, Eun Ju Yun, Kyoung Heon Kim, and Sanghyun Lim. 2023. "Anti-Menopausal Effect of Soybean Germ Extract and Lactobacillus gasseri in the Ovariectomized Rat Model" Nutrients 15, no. 20: 4485. https://doi.org/10.3390/nu15204485
APA StyleLee, S.-H., Lim, T.-J., Yun, E. J., Kim, K. H., & Lim, S. (2023). Anti-Menopausal Effect of Soybean Germ Extract and Lactobacillus gasseri in the Ovariectomized Rat Model. Nutrients, 15(20), 4485. https://doi.org/10.3390/nu15204485