

Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioactive Components of SI and Prediction of Target Genes
2.2. Target Genes of RA
2.3. Protein–Protein Interaction (PPI) Network Analysis
2.4. Gene Ontology and Pathway Analysis
2.5. Network Visualization
2.6. Molecular Docking
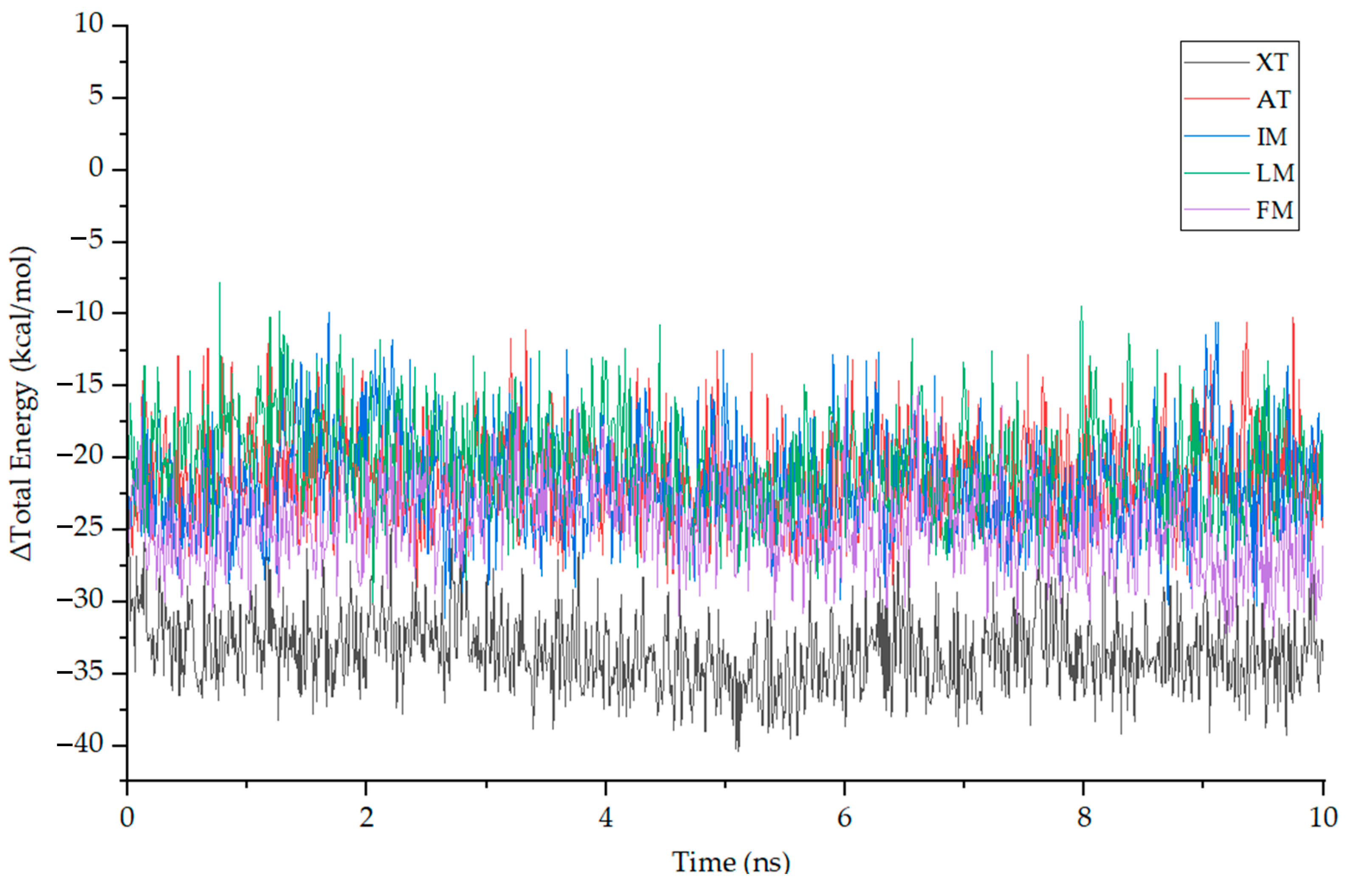
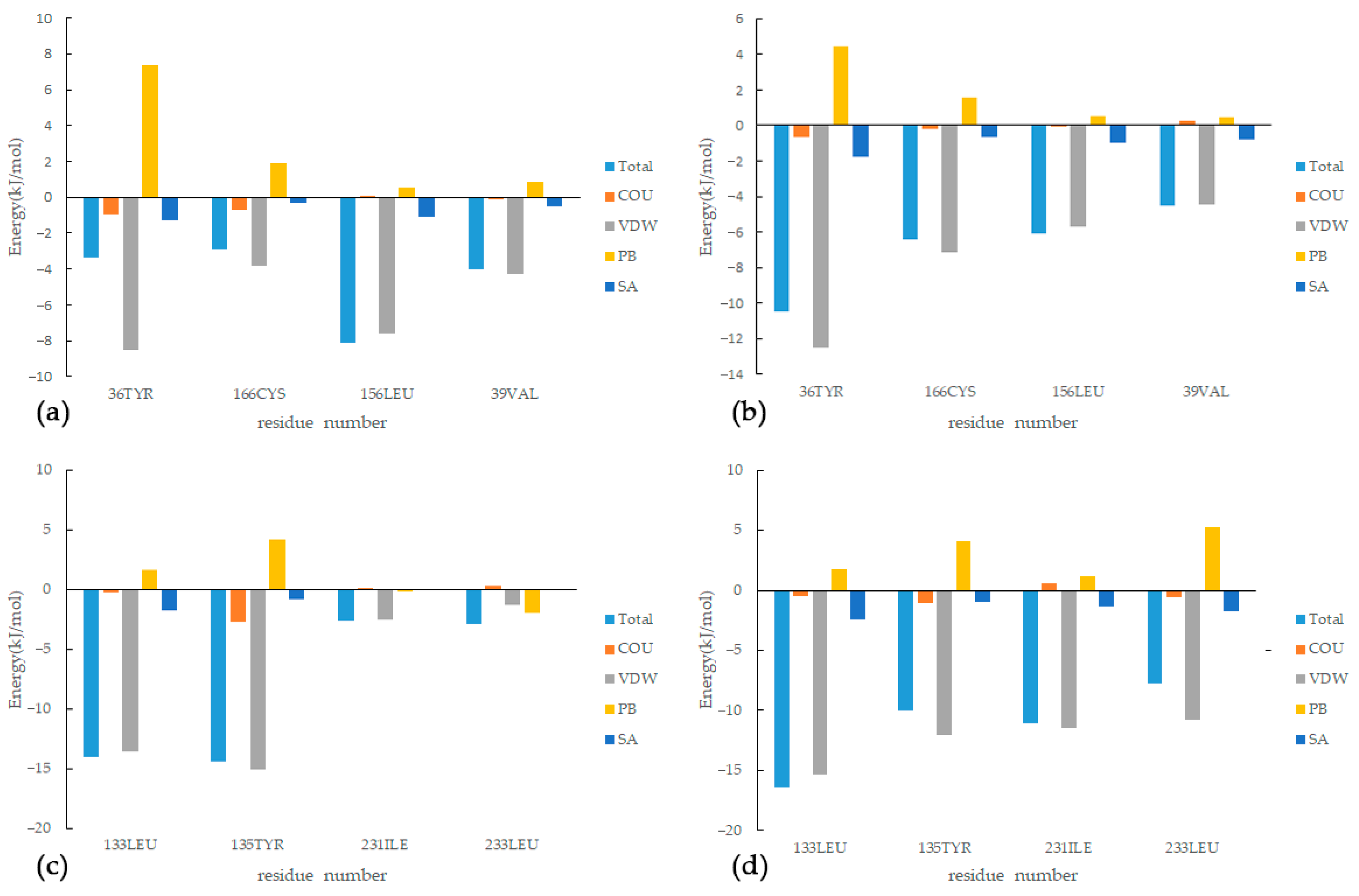
2.7. Molecular Dynamics (MD) Simulation
3. Results
3.1. Bioactive Components and Key Targets
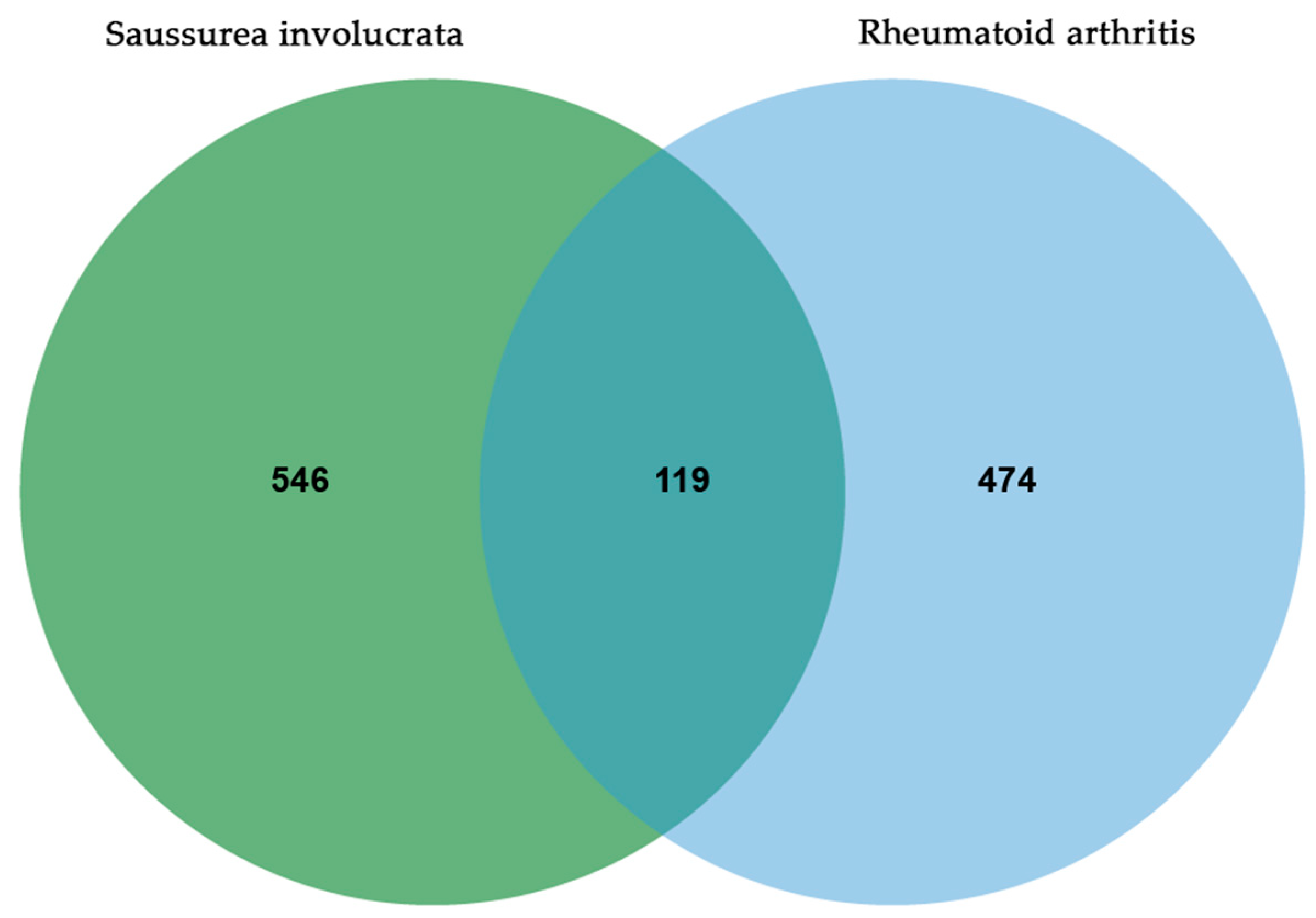
3.2. PPI Network Analysis
3.3. KEGG Pathway Enrichment Analysis
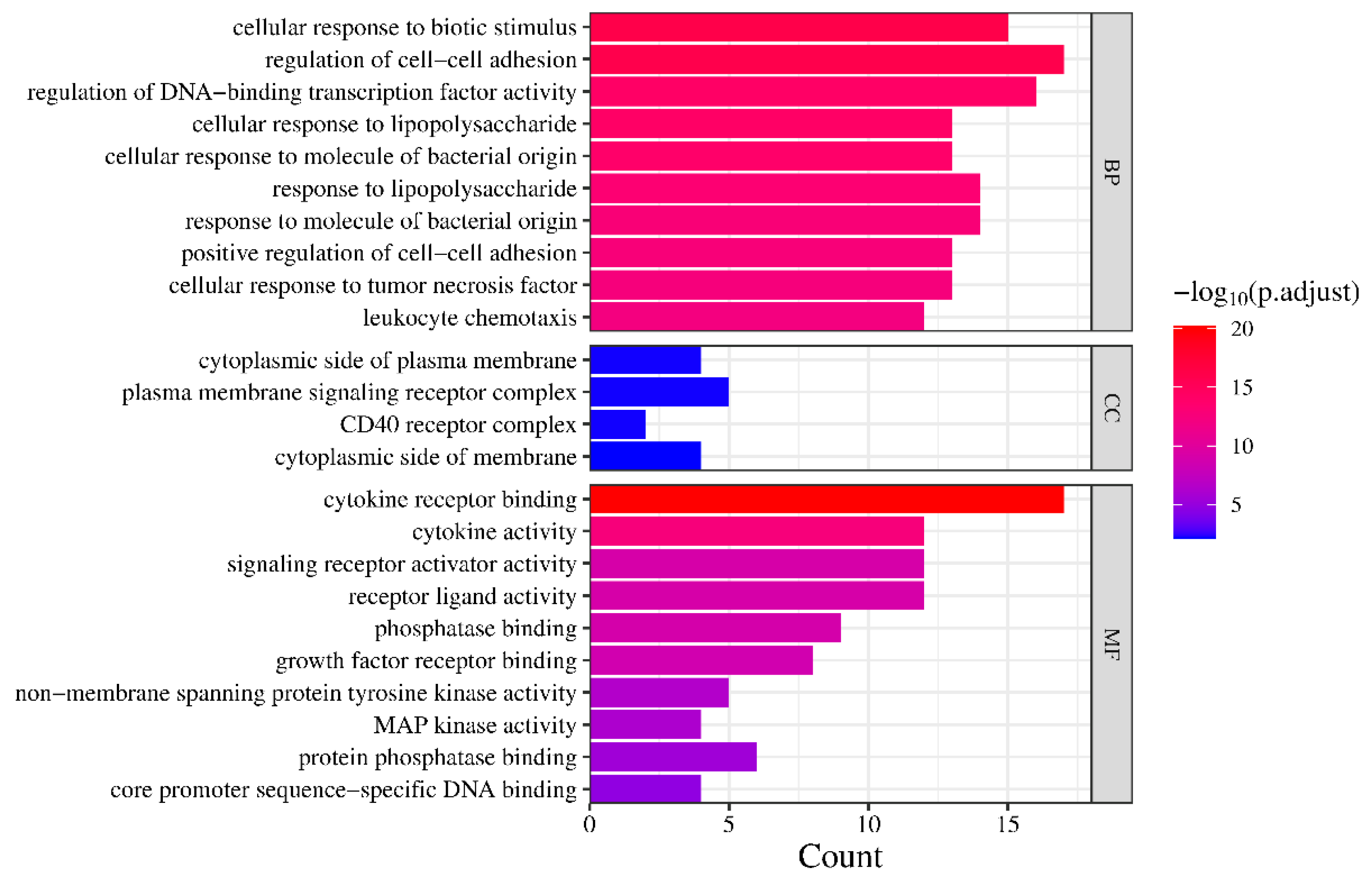
3.4. GO Enrichment Analysis
3.5. Network Constructions and Analysis
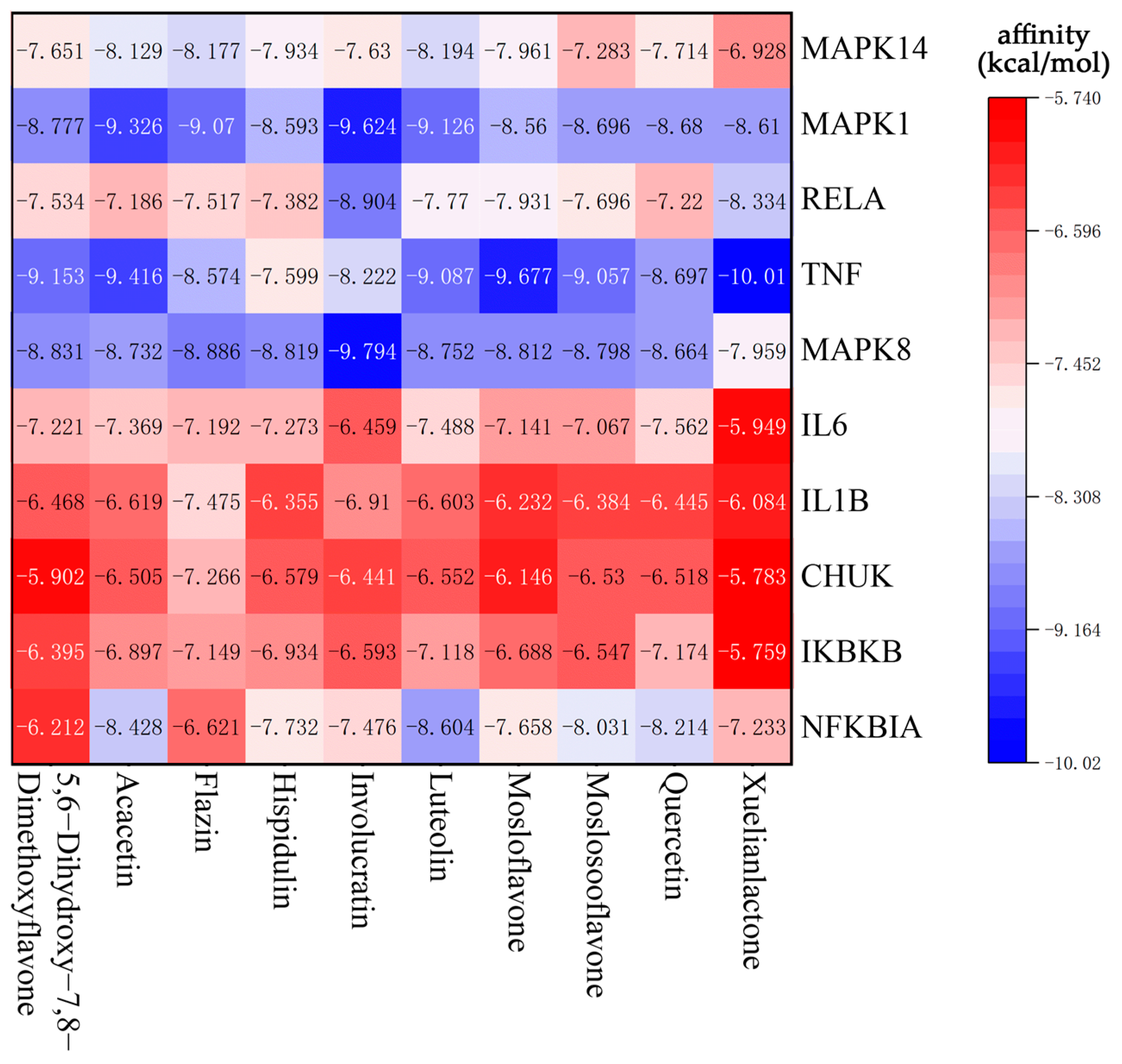
3.6. Molecular Docking
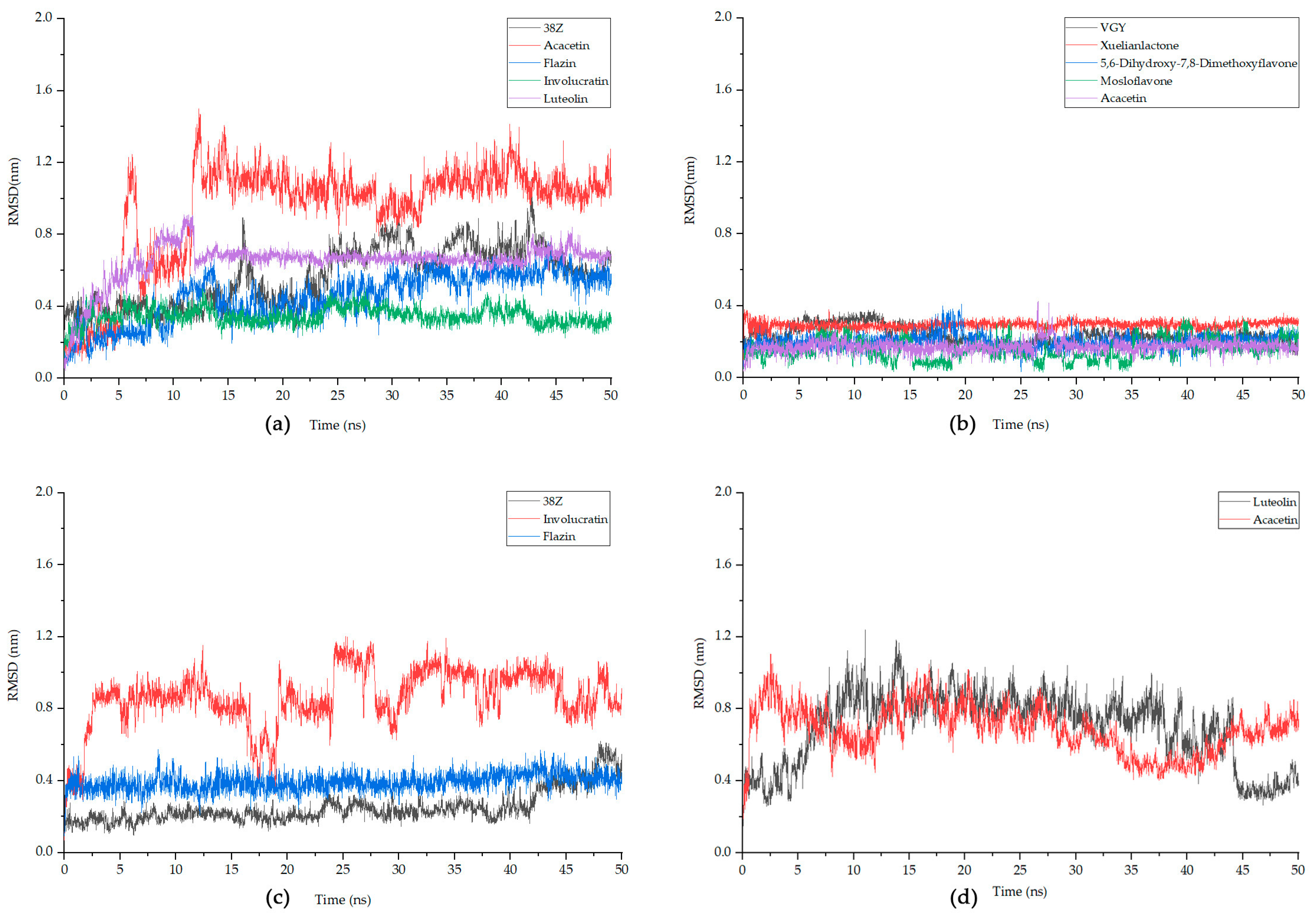
3.7. Molecular Dynamics Simulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Almutairi, K.; Nossent, J.; Preen, D.; Keen, H.; Inderjeeth, C. The global prevalence of rheumatoid arthritis: A meta-analysis based on a systematic review. Rheumatol. Int. 2021, 41, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhu, S.; Tan, A.; Xie, X. Disease burden and quality of life of rheumatoid arthritis in China: A systematic review. Chin. J. Evid. Based Med. 2013, 13, 300–307. [Google Scholar] [CrossRef]
- Zeng, X.; Tian, X.; Li, M. China Rheumatoid Arthritis Development Report 2020; Liaoning Science and Technology Press: Shenyang, China, 2021. [Google Scholar]
- Tian, X.; Wang, Q.; Li, M.; Zhao, Y.; Zhang, Z.; Huang, C.; Liu, Y.; Xu, H.; Chen, Y.; Wu, L. 2018 Chinese guidelines for the diagnosis and treatment of rheumatoid arthritis. Rheumatol. Immunol. Res. 2021, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- van Vollenhoven, R.F. Sex differences in rheumatoid arthritis: More than meets the eye. BMC Med. 2009, 7, 12. [Google Scholar]
- Goemaere, S.; Ackerman, C.; Goethals, K.; De Keyser, F.; Van der Straeten, C.; Verbruggen, G.; Mielants, H.; Veys, E. Onset of symptoms of rheumatoid arthritis in relation to age, sex and menopausal transition. J. Rheumatol. 1990, 17, 1620–1622. [Google Scholar] [CrossRef]
- Hitchon, C.A.; El-Gabalawy, H.S. Infection and rheumatoid arthritis: Still an open question. Curr. Opin. Rheumatol. 2011, 23, 352–357. [Google Scholar] [CrossRef]
- Jiang, X.; Alfredsson, L. Modifiable environmental exposure and risk of rheumatoid arthritis—Current evidence from genetic studies. Arthritis Res. Ther. 2020, 22, 154. [Google Scholar] [CrossRef]
- Wu, D.; Luo, Y.; Li, T.; Zhao, X.; Lv, T.; Fang, G.; Ou, P.; Li, H.; Luo, X.; Huang, A. Systemic complications of rheumatoid arthritis: Focus on pathogenesis and treatment. Front. Immunol. 2022, 13, 1051082. [Google Scholar] [CrossRef]
- Zhang, W.; Anis, A.H. The economic burden of rheumatoid arthritis: Beyond health care costs. Clin. Rheumatol. 2011, 30, 25–32. [Google Scholar] [CrossRef]
- Fidahic, M.; Kadic, A.J.; Radic, M.; Puljak, L. Celecoxib for rheumatoid arthritis. Cochrane Database Syst. Rev. 2017. [Google Scholar] [CrossRef]
- Yeo, J.; Lee, Y.M.; Lee, J.; Park, D.; Kim, K.; Kim, J.; Park, J.; Kim, W.J. Nitric oxide-scavenging nanogel for treating rheumatoid arthritis. Nano Lett. 2019, 19, 6716–6724. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.; Mousavi, M.J.; Jamalzehi, S.; Alimohammadi, R.; Bezvan, M.H.; Mohammadi, H.; Aslani, S. Strategies toward rheumatoid arthritis therapy; the old and the new. J. Cell. Physiol. 2019, 234, 10018–10031. [Google Scholar] [CrossRef] [PubMed]
- Shea, B.; Swinden, M.V.; Ghogomu, E.T.; Ortiz, Z.; Katchamart, W.; Rader, T.; Bombardier, C.; Wells, G.A.; Tugwell, P. Folic acid and folinic acid for reducing side effects in patients receiving methotrexate for rheumatoid arthritis. Cochrane Database Syst. Rev. 2013, 2014, CD000951. [Google Scholar] [CrossRef]
- Tu, A.B.; Lewis, J.S. Biomaterial-based immunotherapeutic strategies for rheumatoid arthritis. Drug Deliv. Transl. Res. 2021, 11, 2371–2393. [Google Scholar] [CrossRef] [PubMed]
- Junshan, L.; Shaoqing, C. Advances in Chemical and Pharmacological Research of Saussurea. Chin. Pharm. J. 1998, 3–6. [Google Scholar]
- Ruiping, Y.; Yongfeng, J.; Liang, W. Research on Drug Resources of Saussurea. In Proceedings of the International Congress of Traditional Medicine, Beijing, China, 22–24 April 2000; p. 2. [Google Scholar]
- Committee, N.P. Pharmacopoeia of People’s Republic of China. Part 1; Medicine Science and Technology Press of China: Beijing, China, 2020; pp. 55–56. [Google Scholar]
- Fan, W.-X.; Yang, W.-P.; Liu, H.-S. Research progress on culture technologies, chemical components, and pharmacological activities of Saussurea involucrata cells. Chin. J. Chin. Mater. Med. 2021, 46, 3522–3528. [Google Scholar] [CrossRef]
- Chik, W.-I.; Zhu, L.; Fan, L.-L.; Yi, T.; Zhu, G.-Y.; Gou, X.-J.; Tang, Y.-N.; Xu, J.; Yeung, W.-P.; Zhao, Z.-Z. Saussurea involucrata: A review of the botany, phytochemistry and ethnopharmacology of a rare traditional herbal medicine. J. Ethnopharmacol. 2015, 172, 44–60. [Google Scholar] [CrossRef]
- Cao, Z.-X.; Li, H.-H.; Li, A.; Liu, P.-Y.; Zhao, Y.-Y.; Mao, P.-Q.; Li, G.-L. Analysis of flavonoids and antitumor activity of transgenic Saussurea involucrate. Chin. J. Chin. Mater. Med. 2018, 43, 4491–4497. [Google Scholar] [CrossRef]
- Jing, L.; He, L.; Fan, P.; Jia, Z.; Ma, H. Chemical constituents with anti-hypoxia activity from Saussurea involucrata. Zhong Yao Cai 2015, 38, 89–92. [Google Scholar] [CrossRef]
- Huan, W.; Qiu, X.; Xu, F. A systematic review clinical efficacy about Xuelian treatment for rheumatoid arthritis. Chin. Pract. Med. 2008, 3, 59–60. [Google Scholar] [CrossRef]
- Song, S.; Zhou, J.; Li, Y.; Liu, J.; Li, J.; Shu, P. Network pharmacology and experimental verification based research into the effect and mechanism of Aucklandiae Radix–Amomi Fructus against gastric cancer. Sci. Rep. 2022, 12, 9401. [Google Scholar] [CrossRef] [PubMed]
- Na, Z.; Fengrong, Z.; Huayong, T.; Qingqing, L.; Xiulan, H.; Zhiyong, L. Benefit evaluation of Tujia medicine Tianzhusan added medicine prescription based on traditionaknowledge investigation and network pharmacology. J. Hunan Univ. Chin. Med. 2022, 42, 633–641. [Google Scholar]
- Ye, X.W.; Wang, H.L.; Cheng, S.Q.; Xia, L.J.; Xu, X.F.; Li, X.R. Network Pharmacology-Based Strategy to Investigate the Pharmacologic Mechanisms of Coptidis Rhizoma for the Treatment of Alzheimer’s Disease. Front. Aging Neurosci. 2022, 14, 890046. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Jianguo, X.; Guihua, L.; Yongjun, Z.; Xincun, W.; Guipeng, X. In situ rat intestinal absorption of two active components in Saussurea involucrate. Chin. J. Hosp. Pharm. 2011, 31, 1407–1410. Available online: https://www.cnki.net/ (accessed on 12 April 2023).
- Wu, Q.; Ma, J.; Shu, Z.; Ren, Y.; Li, D.-Z.; Zhang, Y.-L. Potential therapeutic effect of Saussureae Involucratae Herba on breast cancer and its mechanism based on network pharmacology. Chin. J. Chin. Mater. Med. 2021, 46, 4824–4832. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Y.; Liu, Q.; Ye, C.; Zhao, S.; Zhang, H. Ferula sinkiangensis against gastric cancer: A network pharmacology, molecular docking and cell experiment study. Transl. Cancer Res. 2023, 12, 743. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, Q.; Wang, Y. Network pharmacology-based investigation of potential targets of astragalus membranaceous-angelica sinensis compound acting on diabetic nephropathy. Sci. Rep. 2021, 11, 19496. [Google Scholar] [CrossRef]
- Choi, N.-R.; Jung, D.; Kim, S.-C.; Park, J.-W.; Choi, W.-G.; Kim, B.-J. Analysis of Network Pharmacological Efficacy and Therapeutic Effectiveness in Animal Models for Functional Dyspepsia of Foeniculi fructus. Nutrients 2023, 15, 2644. [Google Scholar] [CrossRef]
- Khan, S.A.; Lee, T.K.W. Network-pharmacology-based study on active phytochemicals and molecular mechanism of cnidium monnieri in treating hepatocellular carcinoma. Int. J. Mol. Sci. 2022, 23, 5400. [Google Scholar] [CrossRef]
- Shi, H.; Tian, S.; Tian, H. Network pharmacology interpretation of fuzheng–jiedu decoction against colorectal cancer. Evid. Based Complement. Alternat. Med. 2021, 2021, 4652492. [Google Scholar] [CrossRef]
- Hu, H.; Wang, H.; Yang, X.; Li, Z.; Zhan, W.; Zhu, H.; Zhang, T. Network pharmacology analysis reveals potential targets and mechanisms of proton pump inhibitors in breast cancer with diabetes. Sci. Rep. 2023, 13, 7623. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, Y.; Wang, W.; He, Y.; Zhong, H.; Zhou, X.; Chen, Y.; Cai, X.J.; Liu, L.Q. Mechanisms underlying the therapeutic effects of Qingfeiyin in treating acute lung injury based on GEO datasets, network pharmacology and molecular docking. Comput. Biol. Med. 2022, 145, 105454. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, X.; Li, G. Integrated bioinformatics and network pharmacology to identify the therapeutic target and molecular mechanisms of Huangqin decoction on ulcerative Colitis. Sci. Rep. 2022, 12, 159. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Mohammed, S.A.D.; Lu, F.; Chen, P.; Wang, Y.; Liu, S. Network Pharmacology and Molecular Docking-Based Mechanism Study to Reveal Antihypertensive Effect of Gedan Jiangya Decoction. BioMed Res. Int. 2022, 2022, 3353464. [Google Scholar] [CrossRef]
- Bai, X.; Tang, Y.; Li, Q.; Chen, Y.; Liu, D.; Liu, G.; Fan, X.; Ma, R.; Wang, S.; Li, L. Network pharmacology integrated molecular docking reveals the bioactive components and potential targets of Morinda officinalis–Lycium barbarum coupled-herbs against oligoasthenozoospermia. Sci. Rep. 2021, 11, 2220. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, C.; Duan, J.; Zhou, T.; Liu, X.; Lu, S.; Xu, F. A Network Pharmacology Approach to Investigate the Anti-Depressive Mechanism of Gardeniae fructus. Int. J. Pharmacol. 2020, 16, 382–397. [Google Scholar] [CrossRef]
- Yan, H.-Y.; Zou, C.-C. Study on anticoagulant material basis and mechanism of Trichosanthis Semen and its shell and kernel based on spectrum-effect relationship integrated molecular docking. Chin. J. Chin. Mater. Med. 2022, 47, 1370–1382. [Google Scholar] [CrossRef]
- Vidya, N.; Vadivukkarasi, B.; Manivannan, G.; Anbarasu, K. Molecular modeling and docking studies of glutamate racemase in Vibrio vulnificus CMCP6. In Silico Biol. 2008, 8, 471–483. [Google Scholar]
- Cao, J.-Y.; Dong, Q.; Wang, Z.-Y.; Zhao, Y.; Ren, Y.; Liu, C.; Dang, J.; Yu, R.-T.; Tao, Y.-D. Arylnaphthalide lignans from Saussurea medusa and their anti-inflammatory activities. Arab. J. Chem. 2022, 15, 104155. [Google Scholar] [CrossRef]
- Jiménez, J.; Doerr, S.; Martínez-Rosell, G.; Rose, A.S.; De Fabritiis, G. DeepSite: Protein-binding site predictor using 3D-convolutional neural networks. Bioinformatics 2017, 33, 3036–3042. [Google Scholar] [CrossRef]
- Wang, Y. Study on Mechanism of Five flavonoids Inhibiting hIAPP Aggregation and NtMGAM Activity Based on Molecular Simulation. Master’s Thesis, Chongqing University, Chongqing, China, 2020. [Google Scholar]
- Zimeng, A.; Qianfang, F.; Yaruo, X.; Xinming, Y.; Yuning, G.; Xilian, Z. Based on network pharmacology, molecular docking, and molecular dynamics simulation to explore the target mechanism of Rongchang Capsules in the treatment of pediatric epilepsy. Tianjin J. Tradit. Chin. Med. 2023, 40, 495–505. [Google Scholar]
- Valdés-Tresanco, M.S.; Valdés-Tresanco, M.E.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A New Tool to Perform End-State Free Energy Calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef] [PubMed]
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [PubMed]
- Saranya, P.; Karunya, R.; Keerthi Varshini, G.; Kowsikan, K.; Prathiksha, R. In-silico docking studies of selected phytochemicals against papain like protease of SARS-CoV-2. Vegetos 2023, 36, 188–194. [Google Scholar] [CrossRef]
- Wilson Alphonse, C.R.; Kannan, R.R. In silico exploration of antioxidants as oxidation protectant for PITRM1 peptidase activity, an Alzheimer disease target. J. Cell. Biochem. 2023, 124, 1002–1011. [Google Scholar] [CrossRef]
- Zhao, Q.; Deng, P.-Y.; Wang, H.-Y.; Luo, Q.; Tian, Y.-F.; Wang, G.-X.; Yang, Y.-Z.; Li, C.-G.; Chen, G. Molecular dynamics simulation of the interactions between puerarin and acetylcholinesterase. J. At. Mol. Phys. 2022, 39, 27–33. [Google Scholar] [CrossRef]
- Ralph, J.A.; Morand, E.F. MAPK phosphatases as novel targets for rheumatoid arthritis. Expert Opin. Ther. Targets 2008, 12, 795–808. [Google Scholar] [CrossRef]
- Liu, S.; Ma, H.; Zhang, H.; Deng, C.; Xin, P. Recent advances on signaling pathways and their inhibitors in rheumatoid arthritis. Clin. Immunol. 2021, 230, 108793. [Google Scholar] [CrossRef]
- Ponce, C.; Torres, M.; Galleguillos, C.; Sovino, H.; Boric, M.A.; Fuentes, A.; Johnson, M.C. Nuclear factor κB pathway and interleukin-6 are affected in eutopic endometrium of women with endometriosis. Reproduction 2009, 137, 727–737. [Google Scholar] [CrossRef]
- Liang, Y.; Zhou, Y.; Shen, P. NF-kappaB and its regulation on the immune system. Cell. Mol. Immunol. 2004, 1, 343–350. [Google Scholar]
- Kim, E.K.; Kwon, J.-E.; Lee, S.-Y.; Lee, E.-J.; Kim, D.S.; Moon, S.-J.; Lee, J.; Kwok, S.-K.; Park, S.-H.; Cho, M.-L. IL-17-mediated mitochondrial dysfunction impairs apoptosis in rheumatoid arthritis synovial fibroblasts through activation of autophagy. Cell Death Dis. 2018, 8, e2565. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, W.B.; Miossec, P. IL-17 as a future therapeutic target for rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, S.; Dahal, K.; Sharma, S. Anti-IL-17 therapy in treatment of rheumatoid arthritis: A systematic literature review and meta-analysis of randomized controlled trials. Rheumatol. Int. 2016, 36, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, Y. Function and regulation of the osteoclast in the pathological changes of bone destruction in rheumatoid arthritis. Chin. J. Osteoporos. 2016, 22, 1168–1173. [Google Scholar] [CrossRef]
- Perricone, C.; Ceccarelli, F.; Matteo, S.; Di Carlo, G.; Bogdanos, D.P.; Lucchetti, R.; Pilloni, A.; Valesini, G.; Polimeni, A.; Conti, F. Porphyromonas gingivalis and rheumatoid arthritis. Curr. Opin. Rheumatol. 2019, 31, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Volin, M.V. Soluble adhesion molecules in the pathogenesis of rheumatoid arthritis. Curr. Pharm. Des. 2005, 11, 633–653. [Google Scholar] [CrossRef]
- Hao, J.; Qi, F.; Wang, H.; Su, L.; Li, X.; Zhang, N.; Sun, W.; Wei, W. Network pharmacology-based prediction of inhibiting leukocyte recruitment and angiogenesis of total glucosides of peony against rheumatoid arthritis. Ann. Palliat. Med. 2022, 11, 3085101. [Google Scholar] [CrossRef]
- Tang, M.; Zeng, Y.; Peng, W.; Xie, X.; Yang, Y.; Ji, B.; Li, F. Pharmacological aspects of natural quercetin in rheumatoid arthritis. Drug Des. Dev. Ther. 2022, ume 16, 2043–2053. [Google Scholar] [CrossRef]
- Liu, X.; Tao, T.; Yao, H.; Zheng, H.; Wang, F.; Gao, Y. Mechanism of action of quercetin in rheumatoid arthritis models: Meta-analysis and systematic review of animal studies. Inflammopharmacology 2023, 31, 1629–1645. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, J.; Huang, Q.; Guo, L. Luteolin inhibits proliferation and affects the function of stimulated rat synovial fibroblasts. Cell Biol. Int. 2009, 33, 135–147. [Google Scholar] [CrossRef]
- Choi, E.M.; Lee, Y.S. Luteolin suppresses IL-1β-induced cytokines and MMPs production via p38 MAPK, JNK, NF-kappaB and AP-1 activation in human synovial sarcoma cell line, SW982. Food Chem. Toxicol. 2010, 48, 2607–2611. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Yang, Z.G.; Hu, P.F.; Bao, J.P.; Wu, L.D. Acacetin inhibits expression of matrix metalloproteinases via a MAPK-dependent mechanism in fibroblast-like synoviocytes. J. Cell. Mol. Med. 2015, 19, 1910–1915. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, N.; Hagiwara, M.; Tada, H.; Komatsu, T.; Sugiura, S.; Kobayashi, K.; Kato, Y.; Ishida, N.; Nishida, K.; Ninomiya, M. Acacetin inhibits expression of E-selectin on endothelial cells through regulation of the MAP kinase signaling pathway and activation of NF-κB. Immunopharmacol. Immunotoxicol. 2013, 35, 471–477. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | Molecular Formula | Molecular Weight | PubChem CID |
|---|---|---|---|---|
| 1 | Dinatin | C16H12O6 | 300.26 | 5281628 |
| 2 | Alloisoimperatorin | C16H14O4 | 270.28 | 5317436 |
| 3 | Beta-Sitosterol | C29H50O | 414.70 | 222284 |
| 4 | Kaempferol | C15H10O6 | 286.24 | 5280863 |
| 5 | Luteolin | C15H10O6 | 286.24 | 5280445 |
| 6 | Flazin | C17H12N2O4 | 308.29 | 5377686 |
| 7 | Quercetin | C15H10O7 | 302.23 | 5280343 |
| 8 | Involucratin | C20H27NO4 | 345.40 | 15628146 |
| 9 | Guaianolide | C14H20O3 | 236.31 | - |
| 10 | Eriodictyol | C15H12O6 | 288.25 | 440735 |
| 11 | Hispidulin | C16H12O6 | 300.26 | 5281628 |
| 12 | Methyl Caffeate Acid | C10H10O4 | 194.18 | 689075 |
| 13 | Cinnamic Acid | C9H8O2 | 148.16 | 444539 |
| 14 | Hexadecanoic Acid | C16H32O2 | 256.42 | 985 |
| 15 | 1-Monolinolein | C21H38O4 | 354.50 | 6256628 |
| 16 | (2S,3S,4R)-2-Aminoicosane-1,3,4-Triol | C20H43NO3 | 345.60 | 12302752 |
| 17 | Moslosooflavone | C17H14O5 | 298.29 | 188316 |
| 18 | Mosloflavone | C17H14O5 | 298.29 | 471722 |
| 19 | 5,6-Dihydroxy-7,8-Dimethoxyflavone | C17H14O6 | 314.29 | 821356 |
| 20 | D-3-Phenyllactic Acid | C9H10O3 | 166.17 | 643327 |
| 21 | Alisol C | C30H46O5 | 486.70 | 101306923 |
| 22 | Alisol C Monoacetate | C32H48O6 | 528.70 | 14036813 |
| 23 | (E)-P-Coumaroylagmatine(1+) | C14H21N4O2+ | 277.34 | 25245514 |
| 24 | Alisol B | C30H48O4 | 472.70 | 15558620 |
| 25 | Acacetin | C16H12O5 | 284.26 | 5280442 |
| 26 | Oroselol | C14H12O4 | 244.24 | 160600 |
| 27 | Xuelianlactone | C15H20O3 | 248.32 | 147111 |
| No. | Gene Symbol | Gene ID | Gene Name |
|---|---|---|---|
| 1 | TNF | 7124 | Tumor necrosis factor |
| 2 | RELA | 5970 | Transcription factor p65 |
| 3 | IL6 | 3569 | Interleukin-6 |
| 4 | JUN | 3725 | Proto-oncogene c-JUN |
| 5 | MAPK1 | 5594 | Mitogen-activated protein kinase 1 |
| 6 | MAPK14 | 1432 | MAP kinase p38 alpha |
| 7 | IL10 | 3586 | Interleukin-10 |
| 8 | IL1B | 3553 | Interleukin-1 beta |
| 9 | CCL2 | 6347 | C-C motif chemokine 2 |
| 10 | IL4 | 3565 | Interleukin-4 |
| 11 | IL2 | 3558 | Interleukin-2 |
| 12 | STAT1 | 6772 | Signal transducer and activator of transcription 1-alpha/beta |
| 13 | CXCL8 | 3576 | Interleukin-8 |
| 14 | IL1A | 3552 | Interleukin-1 alpha |
| 15 | JAK1 | 3716 | Tyrosine-protein kinase JAK1 |
| 16 | CXCL10 | 3627 | C-X-C motif chemokine 10 |
| 17 | NFKBIA | 4792 | NF-kappa-B inhibitor alpha |
| 18 | JAK3 | 3718 | Tyrosine-protein kinase JAK3 |
| 19 | TP53 | 7157 | Cellular tumor antigen p53 |
| 20 | VEGFA | 7422 | Vascular endothelial growth factor A |
| 21 | MAPK11 | 5600 | MAP kinase p38 beta |
| 22 | TYK2 | 7297 | Tyrosine-protein kinase TYK2 |
| 23 | MAPK8 | 5599 | Mitogen-activated protein kinase 8 |
| 24 | ITGB3 | 3690 | Integrin alpha-2/beta-3 |
| 25 | NR3C1 | 2908 | Glucocorticoid receptor |
| 26 | PPARA | 5465 | Peroxisome proliferator-activated receptor alpha |
| 27 | PTK2 | 5747 | Focal adhesion kinase 1 |
| 28 | CHUK | 1147 | Inhibitor of nuclear factor kappa-B kinase subunit alpha |
| 29 | IKBKB | 3551 | Inhibitor of nuclear factor kappa-B kinase subunit beta |
| 30 | MMP3 | 4314 | Stromelysin-1 |
| 31 | MMP2 | 4313 | 72 kDa type IV collagenase |
| 32 | CD40LG | 959 | CD40 ligand |
| 33 | SYK | 6850 | Tyrosine-protein kinase SYK |
| No. | Ingredients | Degree |
|---|---|---|
| 1 | Quercetin | 60 |
| 2 | Luteolin | 43 |
| 3 | Acacetin | 33 |
| 4 | Xuelianlactone | 26 |
| 5 | Moslosooflavone | 25 |
| 6 | Hispidulin | 24 |
| 7 | Mosloflavone | 24 |
| 8 | Involucratin | 23 |
| 9 | 5,6-Dihydroxy-7,8-Dimethoxyflavone | 23 |
| 10 | Flazin | 21 |
| No. | Receptor | Degree of Receptor | Structure | Target Chain | Target Position | Ligand | Affinity (kcal/mol) |
|---|---|---|---|---|---|---|---|
| 1 | MAPK14 | 38 | 5WJJ | A | 1–360 | Luteolin | −8.194 |
| 2 | MAPK1 | 37 | 4QTA | A | 1–360 | Involucratin | −9.624 |
| 3 | RELA | 35 | 6NV2 | P | 39–51 | Involucratin | −8.904 |
| 4 | TNF | 33 | 7JRA | A/B/C | 77–233 | Xuelianlactone | −10.01 |
| 5 | MAPK8 | 33 | 4QTD | A | 1–363 | Involucratin | −9.794 |
| 6 | IL6 | 33 | 7NXZ | A | 30–212 | Quercetin | −7.562 |
| 7 | IL1B | 31 | 5R8Q | A | 117–269 | Flazin | −7.475 |
| 8 | CHUK | 31 | 3BRT | A/C | 732–745 | Flazin | −7.266 |
| 9 | IKBKB | 30 | 3BRT | A/C | 701–730 | Quercetin | −7.174 |
| 10 | NFKBIA | 28 | 1IKN | D | 67–302 | Luteolin | −8.604 |
| Target | MAPK1 | ||||
| Compounds | 38Z | Acacetin | Flazin | Involucratin | Luteolin |
| RMSD Average (nm) | 0.567 ± 0.162 | 0.940 ± 0.290 | 0.455 ± 0.139 | 0.346 ± 0.048 | 0.650 ± 0.112 |
| Target | TNF | ||||
| Compounds | VGY | Xuelianlactone | 5,6-Dihydroxy-7,8- Dimethoxyflavone | Mosloflavone | Acacetin |
| RMSD Average (nm) | 0.235 ± 0.049 | 0.295 ± 0.022 | 0.205 ± 0.040 | 0.155 ± 0.060 | 0.175 ± 0.034 |
| Target | MAPK8 | NFKIBA | |||
| Compounds | 38Z | Involucratin | Flazin | Luteolin | Acacetin |
| RMSD Average (nm) | 0.251 ± 0.091 | 0.862 ± 0.160 | 0.390 ± 0.049 | 0.698 ± 0.199 | 0.686 ± 0.132 |
| Complex | Contribution | |||||||
|---|---|---|---|---|---|---|---|---|
| ΔG | −TΔS | ΔH | MM | PB | SA | COU | VDW | |
| XT | −30.03 | 5.35 | −35.38 | −45.34 | 14.48 | −4.53 | −3.20 | −42.13 |
| AT | −23.42 | 1.73 | −25.15 | −41.16 | 21.43 | −5.42 | −5.74 | −35.42 |
| IM | −11.96 | 3.69 | −15.65 | −45.88 | 35.93 | −5.71 | −9.88 | −35.99 |
| LM | −10.99 | 3.74 | −14.73 | −43.06 | 33.36 | −5.03 | −10.11 | −32.95 |
| FM | −22.00 | 2.89 | −24.89 | −43.49 | 23.80 | −5.20 | −5.47 | −38.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Wu, X.; Yu, R. Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation. Nutrients 2023, 15, 4294. https://doi.org/10.3390/nu15194294
Chen J, Wu X, Yu R. Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation. Nutrients. 2023; 15(19):4294. https://doi.org/10.3390/nu15194294
Chicago/Turabian StyleChen, Jinghua, Xiaoke Wu, and Ruitao Yu. 2023. "Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation" Nutrients 15, no. 19: 4294. https://doi.org/10.3390/nu15194294
APA StyleChen, J., Wu, X., & Yu, R. (2023). Unraveling the Therapeutic Mechanism of Saussurea involucrata against Rheumatoid Arthritis: A Network Pharmacology and Molecular Modeling-Based Investigation. Nutrients, 15(19), 4294. https://doi.org/10.3390/nu15194294