
Exploring the Role of Metabolites in Cancer and the Associated Nerve Crosstalk


Abstract
:1. Introduction
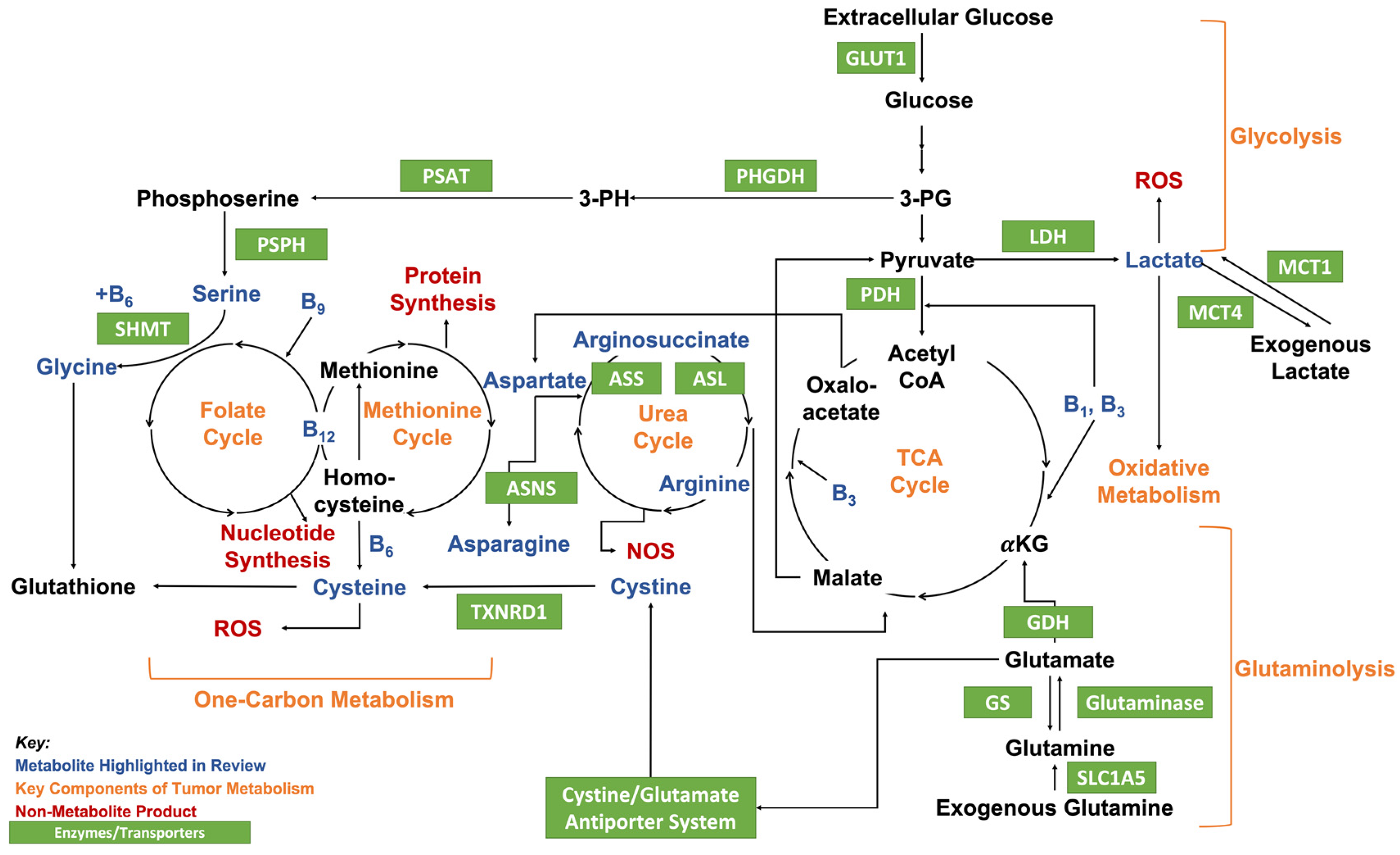
2. Metabolites and Cancer
2.1. Background
2.2. Lactate
2.3. Vitamin A
2.4. Vitamin B
2.5. Vitamin C
2.6. Vitamin D
2.7. Vitamin E
2.8. Vitamin K
2.9. Asparagine
2.10. Arginine
2.11. Serine and Glycine
2.12. Cysteine
3. Metabolites and Cancer–Nerve Crosstalk
3.1. Background
3.2. Known Contributors of Cancer–Nerve Crosstalk
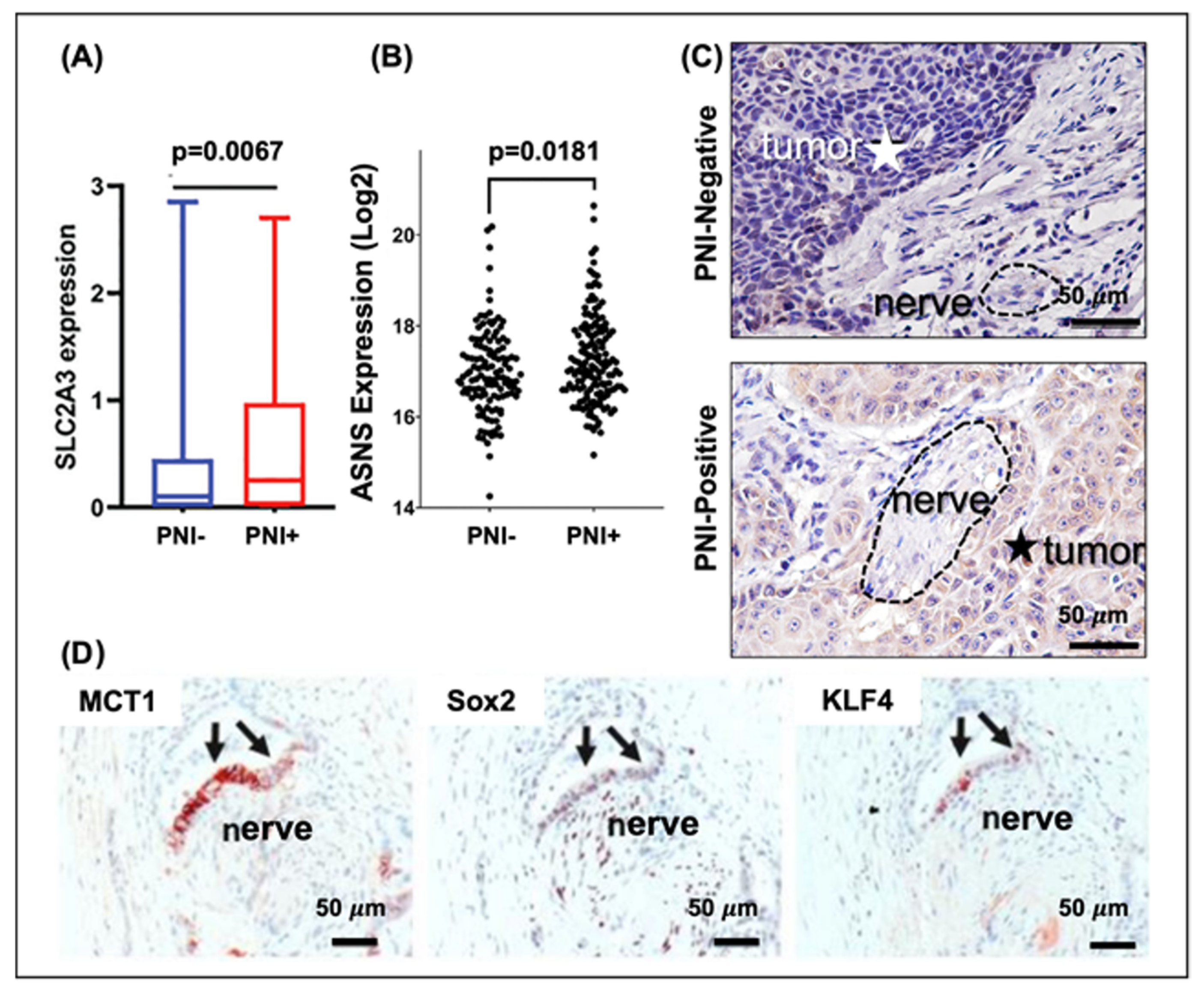
3.2.1. Vitamin C and SLC2A3
3.2.2. Asparagine and Asparagine Synthetase
3.2.3. Serine and Glycine
3.3. Proposed Targets for Cancer–Nerve Crosstalk Research
3.3.1. Lactate
3.3.2. Vitamin A
3.3.3. Vitamin B
3.3.4. Vitamin D
3.3.5. Vitamin E and K
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Cancer Facts & Figures; American Cancer Society: Atlanta, GA, USA, 2022. [Google Scholar]
- International Agency for Research on Cancer; World Health Organization: Geneva, Switzerland, 2020.
- Wu, S.; Zhu, W.; Thompson, P.; Hannun, Y.A. Evaluating intrinsic and non-intrinsic cancer risk factors. Nat. Commun. 2018, 9, 3490. [Google Scholar] [CrossRef] [PubMed]
- Emmons, K.M.; Colditz, G.A. Realizing the Potential of Cancer Prevention—The Role of Implementation Science. N. Engl. J. Med. 2017, 376, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Blot, W.J.; Tarone, R.E. Doll and Peto’s quantitative estimates of cancer risks: Holding generally true for 35 years. J. Natl. Cancer Inst. 2015, 107, djv044. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Bradbury, K.E.; Perez-Cornago, A.; Sinha, R.; Tsilidis, K.K.; Tsugane, S. Diet, nutrition, and cancer risk: What do we know and what is the way forward? BMJ 2020, 368, m511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskins, C.P.; Champ, C.E.; Miller, R.; Vyfhuis, M.A.L. Nutrition in Cancer: Evidence and Equality. Adv. Radiat. Oncol. 2020, 5, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.; Newport, E.; Morten, K.J. The Warburg effect: 80 years on. Biochem. Soc. Trans. 2016, 44, 1499–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanderHeiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.; Kim, S.S.; Lee, J. Cancer cell metabolism: Implications for therapeutic targets. Exp. Mol. Med. 2013, 45, e45. [Google Scholar] [CrossRef] [Green Version]
- Rodic, S.; Vincent, M.D. Reactive oxygen species (ROS) are a key determinant of cancer’s metabolic phenotype. Int. J. Cancer 2018, 142, 440–448. [Google Scholar] [CrossRef]
- Panieri, E.; Santoro, M.M. Ros homeostasis and metabolism: A dangerous liason in cancer cells. Cell Death Dis. 2016, 7, e2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Liu, M.; Zhang, H.; Wei, X.; Wang, J.; Xian, M.; Guo, W. Exploring cysteine regulation in cancer cell survival with a highly specific “lock and Key” fluorescent probe for cysteine. Chem. Sci. 2019, 10, 10065–10071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cluntun, A.A.; Lukey, M.J.; Cerione, R.A.; Locasale, J.W. Glutamine Metabolism in Cancer: Understanding the Heterogeneity. Trends Cancer 2017, 3, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Srivastava, S.; Zhang, J. Starve cancer cells of glutamine: Break the spell or make a hungry monster? Cancers 2019, 11, 804. [Google Scholar] [CrossRef] [Green Version]
- Wise, D.R.; Thompson, C.B. Glutamine addiction: A new therapeutic target in cancer. Trends Biochem. Sci. 2010, 35, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Teoh, S.T.; Ensink, E.; Ogrodzinski, M.P.; Yang, C.; Vazquez, A.I.; Lunt, S.Y. Cysteine catabolism and the serine biosynthesis pathway support pyruvate production during pyruvate kinase knockdown in pancreatic cancer cells. Cancer Metab. 2019, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Fan, J.; Venneti, S.; Cross, J.R.; Takagi, T.; Bhinder, B.; Djaballah, H.; Kanai, M.; Cheng, E.H.; Judkins, A.R.; et al. Asparagine Plays a Critical Role in Regulating Cellular Adaptation to Glutamine Depletion. Mol. Cell 2014, 23, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Li, T.; Da Silva, S.R.; Lee, J.J.; Lu, C.; Eoh, H.; Jung, J.U.; Gao, S.J. A critical role of glutamine and asparagine γ-Nitrogen in nucleotide biosynthesis in cancer cells hijacked by an oncogenic virus. MBio 2017, 8, e01179-17. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Park, K. Targeting Glutamine Metabolism for Cancer Treatment. Biomol. Ther. 2018, 26, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.; van der Meer, L.T.; van Leeuwen, F.N. Amino Acid Depletion Therapies: Starving Cancer Cells to Death. Trends Endocrinol. Metab. 2021, 32, 367–381. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Lee, M.S.; Jadhav, U.; Naqvi, I.; Madha, S.; Adler, A.; Mistry, M.; Naumenko, S.; Lewis, C.A.; Hitchcock, D.S.; et al. Adaptation of pancreatic cancer cells to nutrient deprivation is reversible and requires glutamine synthetase stabilization by mTORC1. Proc. Natl. Acad. Sci. USA 2021, 118, e2003014118. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, R.B.; Chandel, N.S. Targeting glucose metabolism for cancer therapy. J. Exp. Med. 2012, 209, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonveaux, P.; Végran, F.; Schroeder, T.; Wergin, M.C.; Verrax, J.; Rabbani, Z.N.; De Saedeleer, C.J.; Kennedy, K.M.; Diepart, C.; Jordan, B.F.; et al. Targeting lactate-fueled respiration selectively kills hypoxic tumor cells in mice. J. Clin. Investig. 2008, 118, 3930–3942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Y.; Tsai, W.B.; Wangpaichitr, M.; Tsukamoto, T.; Savaraj, N.; Feun, L.G.; Kuo, M.T. Arginine deiminase resistance in melanoma cells is associated with metabolic reprogramming, glucose dependence, and glutamine addiction. Mol. Cancer Ther. 2013, 12, 2581–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truman, J.P.; Ruiz, C.F.; Montal, E.; Garcia-Barros, M.; Mileva, I.; Snider, A.J.; Hannun, Y.A.; Obeid, L.M.; Mao, C. 1-Deoxysphinganine initiates adaptive responses to serine and glycine starvation in cancer cells via proteolysis of sphingosine kinase. J. Lipid Res. 2022, 63, 100154. [Google Scholar] [CrossRef]
- Fu, Y.; Ding, L.; Yang, X.; Ding, Z.; Huang, X.; Zhang, L.; Chen, S.; Hu, Q.; Ni, Y. Asparagine Synthetase-Mediated l-Asparagine Metabolism Disorder Promotes the Perineural Invasion of Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 637226. [Google Scholar] [CrossRef]
- Tarragó-Celada, J.; Foguet, C.; Tarrado-Castellarnau, M.; Marin, S.; Hernández-Alias, X.; Perarnau, J.; Morrish, F.; Hockenbery, D.; Gomis, R.R.; Ruppin, E.; et al. Cysteine and folate metabolism are targetable vulnerabilities of metastatic colorectal cancer. Cancers 2021, 13, 425. [Google Scholar] [CrossRef]
- Muramatsu, H.; Sumitomo, M.; Morinaga, S.; Kajikawa, K.; Kobayashi, I.; Nishikawa, G.; Kato, Y.; Watanabe, M.; Zennami, K.; Kanao, K.; et al. Targeting lactate dehydrogenase-A promotes docetaxel-induced cytotoxicity predominantly in castration-resistant prostate cancer cells. Oncol. Rep. 2019, 42, 224–230. [Google Scholar] [CrossRef]
- Jeschke, J.; O’Hagan, H.M.; Zhang, W.; Vatapalli, R.; Calmon, M.F.; Danilova, L.; Nelkenbrecher, C.; Van Neste, L.; Bijsmans, I.T.G.W.; Van Engeland, M.; et al. Frequent inactivation of Cysteine dioxygenase type 1 contributes to survival of breast cancer cells and resistance to anthracyclines. Clin. Cancer Res. 2013, 19, 3201–3211. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, L.J.; Smith, P.R.; Hiller, L.; Szlosarek, P.W.; Kimberley, C.; Sehouli, J.; Koensgen, D.; Mustea, A.; Schmid, P.; Crook, T. Epigenetic silencing of argininosuccinate synthetase confers resistance to platinum-induced cell death but collateral sensitivity to arginine auxotrophy in ovarian cancer. Int. J. Cancer 2009, 125, 1454–1463. [Google Scholar] [CrossRef]
- Papadimitriou, N.; Markozannes, G.; Kanellopoulou, A.; Critselis, E.; Alhardan, S.; Karafousia, V.; Kasimis, J.C.; Katsaraki, C.; Papadopoulou, A.; Zografou, M.; et al. An umbrella review of the evidence associating diet and cancer risk at 11 anatomical sites. Nat. Commun. 2021, 12, 4579. [Google Scholar] [CrossRef] [PubMed]
- Soldati, L.; Di Renzo, L.; Jirillo, E.; Ascierto, P.A.; Marincola, F.M.; De Lorenzo, A. The influence of diet on anti-cancer immune responsiveness. J. Transl. Med. 2018, 16, 75. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.B.; Aiba, I.; Long, Y.; Lin, H.K.; Feun, L.; Savaraj, N.; Kuo, M.T. Activation of Ras/PI3K/ERK pathway induces c-Myc stabilization to upregulate argininosuccinate synthetase, leading to arginine deiminase resistance in melanoma cells. Cancer Res. 2012, 72, 2622–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, J.C.; Prudner, B.C.; Lange, S.E.S.; Bean, G.R.; Schultze, M.B.; Brashears, C.B.; Radyk, M.D.A.; Redlich, N.; Tzeng, S.C.; Kami, K.; et al. Arginine Deprivation Inhibits the Warburg Effect and Upregulates Glutamine Anaplerosis and Serine Biosynthesis in ASS1-Deficient Cancers. Cell Rep. 2017, 18, 991–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doldo, E.; Costanza, G.; Agostinelli, S.; Tarquini, C.; Ferlosio, A.; Arcuri, G.; Passeri, D.; Scioli, M.G.; Orlandi, A. Vitamin A, cancer treatment and prevention: The new role of cellular retinol binding proteins. Biomed Res. Int. 2015, 2015, 624627. [Google Scholar] [CrossRef] [Green Version]
- Frei, B.; Lawson, S. Vitamin C and cancer revisited. Proc. Natl. Acad. Sci. USA 2008, 105, 11037–11038. [Google Scholar] [CrossRef] [Green Version]
- Unlu, A.; Kirca, O.; Ozdogan, M.; Nayır, E. High-dose vitamin C and cancer. J. Oncol. Sci. 2016, 1, 10–12. [Google Scholar] [CrossRef]
- Norlin, M. Effects of vitamin D in the nervous system: Special focus on interaction with steroid hormone signalling and a possible role in the treatment of brain cancer. J. Neuroendocrinol. 2020, 32, e12799. [Google Scholar] [CrossRef]
- Sailo, B.L.; Banik, K.; Padmavathi, G.; Javadi, M.; Bordoloi, D.; Kunnumakkara, A.B. Tocotrienols: The promising analogues of vitamin E for cancer therapeutics. Pharmacol. Res. 2018, 130, 259–272. [Google Scholar] [CrossRef]
- Miyazawa, S.; Moriya, S.; Kokuba, H.; Hino, H.; Takano, N.; Miyazawa, K. Vitamin K2 induces non-apoptotic cell death along with autophagosome formation in breast cancer cell lines. Breast Cancer 2020, 27, 225–235. [Google Scholar] [CrossRef]
- Mancino, M.; Ametller, E.; Gascón, P.; Almendro, V. The neuronal influence on tumor progression. Biochim. Biophys. Acta-Rev. Cancer 2011, 1816, 105–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, D.A.; Martinez, V.K.; Dougherty, P.M.; Myers, J.N.; Calin, G.A.; Amit, M. Cancer-associated neurogenesis and nerve-cancer crosstalk. Cancer Res. 2021, 6, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Gregory, E.; Dugan, R.; David, G.; Song, Y.H. The biology and engineered modeling strategies of cancer-nerve crosstalk. BBA-Rev. Cancer 2020, 1874, 188406. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, Q.; Lu, Z.; Wang, L.; Ding, L.; Xia, L.; Zhang, H.; Wang, M.; Chen, Y.; Li, G. Role of the nervous system in cancers: A review. Cell Death Discov. 2021, 7, 76. [Google Scholar] [CrossRef]
- Wang, W.; Li, L.; Chen, N.; Niu, C.; Li, Z.; Hu, J.; Cui, J. Nerves in the Tumor Microenvironment: Origin and Effects. Front. Cell Dev. Biol. 2020, 8, 601738. [Google Scholar] [CrossRef]
- Ueno, H.; Hase, K.; Mochizuki, H. Criteria for extramural perineural invasion as a prognostic factor in rectal cancer. J. Br. Surg. 2001, 88, 994–1000. [Google Scholar] [CrossRef]
- Secq, V.; Leca, J.; Bressy, C.; Guillaumond, F.; Skrobuk, P.; Nigri, J.; Lac, S.; Lavaut, M.N.; Bui, T.T.; Thakur, A.K.; et al. Stromal SLIT2 impacts on pancreatic cancer-associated neural remodeling. Cell Death Dis. 2015, 6, e1592. [Google Scholar] [CrossRef]
- Su, D.; Guo, X.; Huang, L.; Ye, H.; Li, Z.; Lin, L.; Chen, R.; Zhou, Q. Tumor-neuroglia interaction promotes pancreatic cancer metastasis. Theranostics 2020, 10, 5029–5047. [Google Scholar] [CrossRef]
- Ferdoushi, A.; Li, X.; Griffin, N.; Faulkner, S.; Jamaluddin, M.F.B.; Gao, F.; Jiang, C.C.; van Helden, D.F.; Tanwar, P.S.; Jobling, P.; et al. Schwann Cell Stimulation of Pancreatic Cancer Cells: A Proteomic Analysis. Front. Oncol. 2020, 10, 1601. [Google Scholar] [CrossRef]
- Deborde, S.; Omelchenko, T.; Lyubchik, A.; Zhou, Y.; He, S.; McNamara, W.F.; Chernichenko, N.; Lee, S.Y.; Barajas, F.; Chen, C.H.; et al. Schwann cells induce cancer cell dispersion and invasion. J. Clin. Investig. 2016, 126, 1538–1554. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Wang, W.; Hu, Z.; Shan, C.; Wang, L.; Wu, B.; Yang, Z.; Yang, X.; Lei, D. BDNF mediated TrkB activation contributes to the EMT progression and the poor prognosis in human salivary adenoid cystic carcinoma. Oral Oncol. 2015, 51, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Mahjour, F.; Dambal, V.; Shrestha, N.; Singh, V.; Noonan, V.; Kantarci, A.; Trackman, P.C. Mechanism for oral tumor cell lysyl oxidase like-2 in cancer development: Synergy with PDGF-AB. Oncogenesis 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Gao, T.; Ju, J.; Zhang, Y.; Ni, Q.; Li, Y.; Zhao, Z.; Chai, J.; Yang, X.; Sun, M. Sympathetic innervation contributes to perineural invasion of salivary adenoid cystic carcinoma via the β2-adrenergic receptor. Onco Targets Ther. 2019, 12, 1475–1495. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Lin, C.; Lin, H.Y.; Chiu, C.M.; Fang, C.W.; Liao, K.F.; Chen, D.R.; Yeh, W.L. Brain-derived neurotrophic factor regulates cell motility in human colon cancer. Endocr. Relat. Cancer 2015, 22, 455–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonkeren, S.L.; Thijssen, M.S.; Vaes, N.; Boesmans, W.; Melotte, V. The emerging role of nerves and glia in colorectal cancer. Cancers 2021, 13, 152. [Google Scholar] [CrossRef]
- Duchalais, E.; Guilluy, C.; Nedellec, S.; Touvron, M.; Bessard, A.; Touchefeu, Y.; Bossard, C.; Boudin, H.; Louarn, G.; Neunlist, M.; et al. Colorectal Cancer Cells Adhere to and Migrate Along the Neurons of the Enteric Nervous System. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Yang, Y.; Liang, X.; Du, G.; Liu, L.; Lu, L.; Dong, J.; Han, H.; Zhang, G. The clinicopathological significance of neurogenesis in breast cancer. BMC Cancer 2014, 14, 724209. [Google Scholar] [CrossRef] [Green Version]
- Sloan, E.K.; Priceman, S.J.; Cox, B.F.; Yu, S.; Pimentel, M.A.; Tangkanangnukul, V.; Arevalo, J.M.G.; Morizono, K.; Karanikolas, B.D.W.; Wu, L.; et al. The sympathetic nervous system induces a metastatic switch in primary breast cancer. Cancer Res. 2010, 70, 7042–7052. [Google Scholar] [CrossRef] [Green Version]
- Narayan, P.; Flynn, J.; Zhang, Z.; Gillespie, E.F.; Mueller, B.; Xu, A.J.; Cuaron, J.; McCormick, B.; Khan, A.J.; Cahlon, O.; et al. Perineural invasion as a risk factor for locoregional recurrence of invasive breast cancer. Sci. Rep. 2021, 11, 12781. [Google Scholar] [CrossRef]
- Kamiya, A.; Hayama, Y.; Kato, S.; Shimomura, A.; Shimomura, T.; Irie, K.; Kaneko, R.; Yanagawa, Y.; Kobayashi, K.; Ochiya, T. Genetic manipulation of autonomic nerve fiber innervation and activity and its effect on breast cancer progression. Nat. Neurosci. 2019, 22, 1289–1305. [Google Scholar] [CrossRef]
- Magnon, C.; Hall, S.J.; Lin, J.; Xue, X.; Gerber, L.; Freedland, S.J.; Frenette, P.S. Autonomic Nerve Development Contributes to Prostate Cancer Progression. Science 2013, 15, 713–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigorski, D.; Gulczyński, J.; Sejda, A.; Rogowski, W.; Iżycka-Świeszewska, E. Investigation of Neural Microenvironment in Prostate Cancer in Context of Neural Density, Perineural Invasion, and Neuroendocrine Profile of Tumors. Front. Oncol. 2021, 11, 710899. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Shang, W.; Min, X.; Weinreb, J.; Li, Q.; Leapman, M.; Wang, L.; Tian, J. Sight and switch off: Nerve density visualization for interventions targeting nerves in prostate cancer. Sci. Adv. 2020, 6, eaax6040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucido, C.T.; Wynja, E.; Madeo, M.; Williamson, C.S.; Schwartz, L.E.; Imblum, B.A.; Drapkin, R.; Vermeer, P.D. Innervation of cervical carcinoma is mediated by cancer-derived exosomes. Gynecol. Oncol. 2019, 154, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, G.-N.; Shi, Y.; Cui, L.; Leng, X.-F.; Huang, J.-M. Perineural invasion in cervical cancer: Pay attention to the indications of nerve-sparing radical hysterectomy. Ann. Transl. Med. 2019, 7, 203. [Google Scholar] [CrossRef]
- Tang, M.; Liu, Q.; Yang, X.; Chen, L.; Yu, J.; Qi, X.; Wang, Y. Perineural invasion as a prognostic risk factor in patients with early cervical cancer. Oncol. Lett. 2019, 17, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.X.; Wang, B.; Yao, Y.N.; Pan, Z.J.; Shen, Q.; Zhou, J.Y. Autonomic nervous infiltration positively correlates with pathological risk grading and poor prognosis in patients with lung adenocarcinoma. Thorac. Cancer 2016, 7, 588–598. [Google Scholar] [CrossRef]
- Azam, S.H.; Pecot, C.V. Cancer’s got nerve: Schwann cells drive perineural invasion. J. Clin. Investig. 2016, 126, 1242–1244. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.; Li, Y.; Chai, J.; Ma, C.; Ni, Q.; Shen, Z.; Wei, J.; Sun, M. The role of perineural invasion on head and neck adenoid cystic carcinoma prognosis: A systematic review and meta-analysis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2016, 122, 691–701. [Google Scholar] [CrossRef]
- Chen, S.-H.; Zhang, B.-Y.; Zhou, B.; Zhu, C.-Z.; Sun, L.-Q.; Feng, Y.-J. Perineural invasion of cancer: A complex crosstalk between cells and molecules in the perineural niche. Am. J. Cancer Res. 2019, 9, 1–21. [Google Scholar]
- Vermeer, P.D. Exosomal induction of tumor innervation. Cancer Res. 2019, 79, 3529–3535. [Google Scholar] [CrossRef]
- Gysler, S.M.; Drapkin, R. Tumor innervation: Peripheral nerves take control of the tumor microenvironment. J. Clin. Investig. 2021, 131, e147276. [Google Scholar] [CrossRef] [PubMed]
- Shirai, K.; Ebata, T.; Oda, K.; Nishio, H.; Nagasaka, T.; Nimura, Y.; Nagino, M. Perineural invasion is a prognostic factor in intrahepatic cholangiocarcinoma. World J. Surg. 2008, 32, 2395–2402. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Huang, R.; Zhang, X.; Shen, P.; Gong, J.; Zhao, J.; Liu, J.; Tang, Q.; Shu, K.; Yin, X.; et al. The impact of multifocal perineural invasion on biochemical recurrence and timing of adjuvant androgen-deprivation therapy in high-risk prostate cancer following radical prostatectomy. Prostate 2017, 77, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Jeffrey, R.B.; Patel, B.N.; Dimaio, M.A.; Rosenberg, J.; Willmann, J.K.; Olcott, E.W. Preoperative Multidetector cT Diagnosis of extrapancreatic Perineural or Duodenal invasion is associated with reduced Postoperative survival after Pancreaticoduodenectomy for Pancreatic adenocarcinoma: Preliminary Experience and Implications for Patient C. Radiology 2016, 281, 816–825. [Google Scholar] [CrossRef]
- Tao, Q.; Zhu, W.; Zhao, X.; Li, M.; Shu, Y.; Wang, D.; Li, X. Perineural Invasion and Postoperative Adjuvant Chemotherapy Efficacy in Patients With Gastric Cancer. Front. Oncol. 2020, 10, 530. [Google Scholar] [CrossRef] [Green Version]
- Bakst, R.L.; Lee, N.; He, S.; Chernichenko, N.; Chen, C.H.; Linkov, G.; Le, H.C.; Koutcher, J.; Vakiani, E.; Wong, R.J. Radiation impairs perineural invasion by modulating the nerve microenvironment. PLoS ONE 2012, 7, e39925. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ma, G.; Ma, Q.; Li, W.; Liu, J.; Han, L.; Duan, W.; Xu, Q.; Liu, H.; Wang, Z.; et al. Neurotransmitter substance P mediates pancreatic cancer perineural invasion via NK-1R in cancer cells. Mol. Cancer Res. 2013, 11, 294–302. [Google Scholar] [CrossRef] [Green Version]
- Kolokythas, A.; Cox, D.P.; Dekker, N.; Schmidt, B.L. Nerve Growth Factor and Tyrosine Kinase A Receptor in Oral Squamous Cell Carcinoma: Is There an Association With Perineural Invasion? J. Oral Maxillofac. Surg. 2010, 68, 1290–1295. [Google Scholar] [CrossRef]
- Pundavela, J.; Roselli, S.; Faulkner, S.; Attia, J.; Scott, R.J.; Thorne, R.F.; Forbes, J.F.; Bradshaw, R.A.; Walker, M.M.; Jobling, P.; et al. Nerve fibers infiltrate the tumor microenvironment and are associated with nerve growth factor production and lymph node invasion in breast cancer. Mol. Oncol. 2015, 9, 1626–1635. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, Y.; Sakitani, K.; Konishi, M.; Asfaha, S.; Jiang, Z.; Tanaka, T.; Dubeykovskaya, Z.A.; Chen, X.; Urbanska, A.M.; Nagar, K.; et al. Nerve growth factor promotes gastric tumorigenesis through aberrant cholinergic signaling. Cancer Cell 2017, 31, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, P.J.; Paulino, A.F.G. Perineural invasion in adenoid cystic carcinoma: Its causation/promotion by brain-derived neurotrophic factor. Hum. Pathol. 2002, 33, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.Y.; Zhou, T.; Chen, W.; Yin, X.D.; Yao, J.H.; Zhang, Y.F. Preliminary study correlating CX3CL1/CX3CR1 expression with gastric carcinoma and gastric carcinoma perineural invasion. World J. Gastroenterol. 2014, 20, 4428–4432. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, S.; Chen, C.H.; Deborde, S.; Bakst, R.L.; Chernichenko, N.; McNamara, W.F.; Lee, S.Y.; Barajas, F.; Yu, Z.; et al. The chemokine (CCL2-CCR2) signaling axis mediates perineural invasion. Mol. Cancer Res. 2015, 13, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Fan, Q.; Wang, Y.; Cui, Y.; Wang, Z.; Yang, L.; Sun, X.; Wang, Y. Schwann Cell-Derived CCL2 Promotes the Perineural Invasion of Cervical Cancer. Front. Oncol. 2020, 10, 19. [Google Scholar] [CrossRef]
- Madeo, M.; Colbert, P.L.; Vermeer, D.W.; Lucido, C.T.; Cain, J.T.; Vichaya, E.G.; Grossberg, A.J.; Muirhead, D.R.; Rickel, A.P.; Hong, Z.; et al. Cancer exosomes induce tumor innervation. Nat. Commun. 2018, 9, 4284. [Google Scholar] [CrossRef] [Green Version]
- Demir, I.E.; Ceyhan, G.O.; Liebl, F.; D’Haese, J.G.; Maak, M.; Friess, H. Neural invasion in pancreatic cancer: The past, present and future. Cancers 2010, 2, 1513–1527. [Google Scholar] [CrossRef] [Green Version]
- Ni, T.; Huang, T.; Gu, S.L.; Wang, J.; Liu, Y.; Sun, X.; Wang, Y.D. DRG neurons promote perineural invasion of endometrial cancer via GluR2. J. Cancer 2020, 11, 2518–2528. [Google Scholar] [CrossRef]
- Sejda, A.; Sigorski, D.; Gulczyński, J.; Wesołowski, W.; Kitlińska, J.; Iżycka-Świeszewska, E. Complexity of Neural Component of Tumor Microenvironment in Prostate Cancer. Pathobiology 2020, 87, 87–99. [Google Scholar] [CrossRef]
- Li, J.; Ma, J.; Han, L.; Xu, Q.; Lei, J.; Duan, W.; Li, W.; Wang, F.; Wu, E.; Ma, Q.; et al. Hyperglycemic tumor microenvironment induces perineural invasion in pancreatic cancer. Cancer Biol. Ther. 2015, 16, 912–921. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, W.; Zhang, X.; Yihe, M.I.N.; Zhao, Y.; Wang, B.; Wei, L.I.; Shuai, M.A.O.; Weili, M.I.N. High-glucose microenvironment promotes perineural invasion of pancreatic cancer via activation of hypoxia inducible factor 1α. Oncol. Rep. 2022, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.H.; Peng, W.B.; Zhang, P.; Yang, X.P.; Zhou, Q. Lactate in the tumour microenvironment: From immune modulation to therapy. EBioMedicine 2021, 73, 103627. [Google Scholar] [CrossRef] [PubMed]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine promotes cancer cell proliferation through use as an amino acid exchange factor. Nat. Commun. 2016, 7, 11457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.L.; Hsu, S.C.; Ann, D.K.; Yen, Y.; Kung, H.J. Arginine signaling and cancer metabolism. Cancers 2021, 13, 3541. [Google Scholar] [CrossRef]
- Alothaim, T.; Charbonneau, M.; Tang, X. HDAC6 inhibitors sensitize non-mesenchymal triple-negative breast cancer cells to cysteine deprivation. Sci. Rep. 2021, 11, 10956. [Google Scholar] [CrossRef]
- Labuschagne, C.F.; van den Broek, N.J.F.; Mackay, G.M.; Vousden, K.H.; Maddocks, O.D.K. Serine, but not glycine, supports one-carbon metabolism and proliferation of cancer cells. Cell Rep. 2014, 7, 1248–1258. [Google Scholar] [CrossRef] [Green Version]
- Maneikyte, J.; Bausys, A.; Leber, B.; Horvath, A.; Feldbacher, N.; Hoefler, G.; Strupas, K.; Stiegler, P.; Schemmer, P. Dietary glycine decreases both tumor volume and vascularization in a combined colorectal liver metastasis and chemotherapy model. Int. J. Biol. Sci. 2019, 15, 1582–1590. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.T.; Qi, Y.; Wang, Y.C.; Chi, K.K.; Chung, Y.; Ouyang, C.; Chen, Y.R.; Oh, M.E.; Sheng, X.; Tang, Y.; et al. Arginine starvation kills tumor cells through aspartate exhaustion and mitochondrial dysfunction. Commun. Biol. 2018, 1, 178. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, O.D.K.; Berkers, C.R.; Mason, S.M.; Zheng, L.; Blyth, K. Serine starvation induces stress and p53 dependent metabolic remodeling in cancer cells. Nature 2019, 493, 542–546. [Google Scholar] [CrossRef]
- Ma, J.L.; Zhao, Y.; Guo, C.Y.; Hu, H.T.; Zheng, L.; Zhao, E.J.; Li, H.L. Dietary vitamin B intake and the risk of esophageal cancer: A meta-analysis. Cancer Manag. Res. 2018, 10, 5395–5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ness, R.A.; Miller, D.D.; Li, W. The role of vitamin D in cancer prevention. Chin. J. Nat. Med. 2015, 13, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Fulan, H.; Changxing, J.; Yi Baina, W.; Wencui, Z.; Chunqing, L.; Fan, W.; Dandan, L.; Dianjun, S.; Tong, W.; Da, P.; et al. Retinol, vitamins A, C, and e and breast cancer risk: A meta-analysis and meta-regression. Cancer Causes Control 2011, 22, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Gallieni, M.; Porta, C.; Nickolas, T.L.; Khairallah, P. Vitamin K effects in human health: New insights beyond bone and cardiovascular health. J. Nephrol. 2020, 33, 239–249. [Google Scholar] [CrossRef]
- Ngo, B.; Van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting cancer vulnerabilities with high-dose vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef]
- Wu, Y.; Ye, Y.; Shi, Y.; Li, P.; Xu, J.; Chen, K.; Xu, E.; Yang, J. Association between vitamin A, retinol intake and blood retinol level and gastric cancer risk: A meta-analysis. Clin. Nutr. 2015, 34, 620–626. [Google Scholar] [CrossRef]
- Abar, L.; Vieira, A.R.; Aune, D.; Stevens, C.; Vingeliene, S.; Navarro Rosenblatt, D.A.; Chan, D.; Greenwood, D.C.; Norat, T. Blood concentrations of carotenoids and retinol and lung cancer risk: An update of the WCRF–AICR systematic review of published prospective studies. Cancer Med. 2016, 5, 2069–2083. [Google Scholar] [CrossRef] [Green Version]
- Comín-Anduix, B.; Boren, J.; Martinez, S.; Moro, C.; Centelles, J.J.; Trebukhina, R.; Petushok, N.; Lee, W.N.P.; Boros, L.G.; Cascante, M. The effect of thiamine supplementation on tumour proliferation: A metabolic control analysis study. Eur. J. Biochem. 2001, 268, 4177–4182. [Google Scholar] [CrossRef]
- Liu, S.; Huang, H.; Lu, X.; Golinski, M.; Comesse, S.; Watt, D.; Grossman, R.B.; Moscow, J.A. Down-regulation of thiamine transporter THTR2 gene expression in breast cancer and its association with resistance to apoptosis. Mol. Cancer Res. 2003, 1, 665–673. [Google Scholar]
- Ng, E.K.O.; Leung, C.P.H.; Shin, V.Y.; Wong, C.L.P.; Ma, E.S.K.; Jin, H.C.; Chu, K.M.; Kwong, A. Quantitative analysis and diagnostic significance of methylated SLC19A3 DNA in the plasma of breast and gastric cancer patients. PLoS ONE 2011, 6, e22233. [Google Scholar] [CrossRef] [Green Version]
- Arendt, J.F.H.; Sørensen, H.T.; Horsfall, L.J.; Petersen, I. Elevated vitamin B12 levels and cancer risk in UK primary care: A thin database cohort study. Cancer Epidemiol. Biomark. Prev. 2019, 28, 814–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanidi, A.; Carreras-Torres, R.; Larose, T.L.; Yuan, J.M.; Stevens, V.L.; Weinstein, S.J.; Albanes, D.; Prentice, R.; Pettinger, M.; Cai, Q.; et al. Is high vitamin B12 status a cause of lung cancer? Int. J. Cancer 2019, 145, 1499–1503. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.H.; Wang, Q.M.; Feng, L.Y.; Ke, Y.D.; Xu, Q.Z.; Wei, A.Y.; Zhang, C.; Ying, R.B. High-dose vitamin C suppresses the invasion and metastasis of breast cancer cells via inhibiting epithelial-mesenchymal transition. Onco Targets Ther. 2019, 12, 7405–7413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Yan, Y.; Ma, Y.; Yang, Y. Vitamin C at high concentrations induces cytotoxicity in malignant melanoma but promotes tumor growth at low concentrations. Mol. Carcinog. 2017, 56, 1965–1976. [Google Scholar] [CrossRef] [PubMed]
- van den Bemd, G.J.C.M.; Chang, G.T.G. Vitamin D and Vitamin D Analogs in Cancer Treatment. Curr. Drug Targets 2005, 3, 85–94. [Google Scholar] [CrossRef]
- Milczarek, M.; Psurski, M.; Kutner, A.; Wietrzyk, J. Vitamin D analogs enhance the anticancer activity of 5-fluorouracil in an in vivo mouse colon cancer model. BMC Cancer 2013, 13, 294. [Google Scholar] [CrossRef] [Green Version]
- Gorham, E.D.; Garland, C.F.; Garland, F.C.; Grant, W.B.; Mohr, S.B.; Lipkin, M.; Newmark, H.L.; Giovannucci, E.; Wei, M.; Holick, M.F. Optimal Vitamin D Status for Colorectal Cancer Prevention. A Quantitative Meta Analysis. Am. J. Prev. Med. 2007, 32, 210–216. [Google Scholar] [CrossRef]
- Wang, Y.; Chiang, Y.H.; Su, T.P.; Hayashi, T.; Morales, M.; Hoffer, B.J.; Lin, S.Z. Vitamin D3 attenuates cortical infarction induced by middle cerebral arterial ligation in rats. Neuropharmacology 2000, 39, 873–880. [Google Scholar] [CrossRef]
- Kannappan, R.; Gupta, S.C.; Kim, J.H.; Aggarwal, B.B. Tocotrienols fight cancer by targeting multiple cell signaling pathways. Genes Nutr. 2012, 7, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Yap, W.N.; Zaiden, N.; Tan, Y.L.; Ngoh, C.P.; Zhang, X.W.; Wong, Y.C.; Ling, M.T.; Yap, Y.L. Id1, inhibitor of differentiation, is a key protein mediating anti-tumor responses of gamma-tocotrienol in breast cancer cells. Cancer Lett. 2010, 291, 187–199. [Google Scholar] [CrossRef]
- Xu, W.L.; Liu, J.R.; Liu, H.K.; Qi, G.Y.; Sun, X.R.; Sun, W.G.; Chen, B.Q. Inhibition of proliferation and induction of apoptosis by γ-tocotrienol in human colon carcinoma HT-29 cells. Nutrition 2009, 25, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Chen, Y.-R.; Liu, X.; Chu, C.-Y.; Shen, L.-J.; Xu, J.; Gaur, S.; Forman, H.J.; Zhang, H.; Zheng, S.; et al. Arginine Starvation Impairs Mitochondrial Respiratory Function in ASS1-Deficient Breast Cancer Cells. Sci. Signal. 2015, 7, ra31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.X.; Cochrane, D.R.; Tessier-Cloutier, B.; Chen, S.Y.; Ho, G.; Pathak, K.V.; Alcazar, I.N.; Farnell, D.; Leung, S.; Cheng, A.; et al. Arginine Depletion Therapy with ADI-PEG20 Limits Tumor Growth in Argininosuccinate Synthase–Deficient Ovarian Cancer, including Small-Cell Carcinoma of the Ovary, Hypercalcemic Type. Clin. Cancer Res. 2020, 26, 4402–4413. [Google Scholar] [CrossRef] [PubMed]
- Selvi, I.; Basar, H.; Baydilli, N.; Murat, K.; Kaymaz, O. The importance of plasma arginine level and its downstream metabolites in diagnosing prostate cancer. Int. Urol. Nephrol. 2019, 51, 1975–1983. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Biancur, D.E.; Parker, S.J.; Yamamoto, K.; Banh, R.S.; Paulo, J.A.; Mancias, J.D.; Kimmelman, A.C. Autophagy is required for proper cysteine homeostasis in pancreatic cancer through regulation of SLC7A11. Proc. Natl. Acad. Sci. USA 2021, 118, e2021475118. [Google Scholar] [CrossRef]
- Fan, T.W.M.; Bruntz, R.C.; Yang, Y.; Song, H.; Chernyavskaya, Y.; Deng, P.; Zhang, Y.; Shah, P.P.; Beverly, L.J.; Qi, Z.; et al. De novo synthesis of serine and glycine fuels purine nucleotide biosynthesis in human lung cancer tissues. J. Biol. Chem. 2019, 294, 13464–13477. [Google Scholar] [CrossRef]
- Bonuccelli, G.; Tsirigos, A.; Whitaker-Menezes, D.; Pavlides, S.; Pestell, R.G.; Chiavarina, B.; Frank, P.G.; Flomenberg, N.; Howell, A.; Martinez-Outschoorn, U.E.; et al. Ketones and lactate “fuel” tumor growth and metastasis: Evidence that epithelial cancer cells use oxidative mitochondrial metabolism. Cell Cycle 2010, 9, 3506–3514. [Google Scholar] [CrossRef]
- San-Millán, I.; Julian, C.G.; Matarazzo, C.; Martinez, J.; Brooks, G.A. Is Lactate an Oncometabolite? Evidence Supporting a Role for Lactate in the Regulation of Transcriptional Activity of Cancer-Related Genes in MCF7 Breast Cancer Cells. Front. Oncol. 2020, 9, 1536. [Google Scholar] [CrossRef]
- Dikshit, A.; Lu, J.; Ford, A.E.; Degan, S.; Jin, Y.J.; Sun, H.; Nichols, A.; Salama, A.K.S.; Beasley, G.; Gooden, D.; et al. Potential Utility of Synthetic D-Lactate Polymers in Skin Cancer. JID Innov. 2021, 1, 100043. [Google Scholar] [CrossRef]
- Apicella, M.; Giannoni, E.; Fiore, S.; Ferrari, K.J.; Fernández-Pérez, D.; Isella, C.; Granchi, C.; Minutolo, F.; Sottile, A.; Comoglio, P.M.; et al. Increased Lactate Secretion by Cancer Cells Sustains Non-cell-autonomous Adaptive Resistance to MET and EGFR Targeted Therapies. Cell Metab. 2018, 28, 848–865.e6. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chong, Y.; Chen, M.; Dai, W.; Zhou, X.; Ji, Y.; Qiu, G.; Du, X. Targeting lactate dehydrogenase a improves radiotherapy efficacy in non-small cell lung cancer: From bedside to bench. J. Transl. Med. 2021, 19, 170. [Google Scholar] [CrossRef] [PubMed]
- Ngo, B.; Kim, E.; Osorio-Vasquez, V.; Doll, S.; Bustraan, S.; Liang, R.J.; Luengo, A.; Davidson, S.M.; Ali, A.; Ferraro, G.B.; et al. Limited environmental serine and glycine confer brain metastasis sensitivity to PHGDH inhibition. Cancer Discov. 2020, 10, 1352–1373. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fu, J.; Hu, B.; Chen, L.; Wang, J.; Fang, J.; Ge, C.; Lin, H.; Pan, K.; Fu, L.; et al. Serine Metabolism Regulates YAP Activity Through USP7 in Colon Cancer. Front. Cell Dev. Biol. 2021, 9, 639111. [Google Scholar] [CrossRef]
- Meiser, J.; Tumanov, S.; Maddocks, O.; Labuschagne, C.F.; Athineos, D.; Van Den Broek, N.; Mackay, G.M.; Gottlieb, E.; Blyth, K.; Vousden, K.; et al. Serine one-carbon catabolism with formate overflow. Sci. Adv. 2016, 2, e1601273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, M.L.; Gladden, L.B.; Nijsten, M.W.N.; Jones, K.B. Lactate and Cancer: Revisiting the Warburg Effect in an Era of Lactate Shuttling. Front. Nutr. 2015, 1, 2014–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Cruz-López, K.G.; Castro-Muñoz, L.J.; Reyes-Hernández, D.O.; García-Carrancá, A.; Manzo-Merino, J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front. Oncol. 2019, 9, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafur, D.; Svrcek, P.; White, B. Differential expression and function of the endogenous lactate receptor, GPR81, in ER alpha-positive/HER2-positive epithelial vs. post-EMT triple-negative mesenchymal breast cancer cells. J. Cancer Metastasis Treat. 2019, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Shi, X.; Zhang, W.; Li, D.; Hu, L.; Yang, J.; Zhao, J.; Wei, S.; Wei, X.; Ruan, X.; et al. LDHA induces EMT gene transcription and regulates autophagy to promote the metastasis and tumorigenesis of papillary thyroid carcinoma. Cell Death Dis. 2021, 12, 347. [Google Scholar] [CrossRef]
- Siddikuzzaman, C.G.; Berlin Grace, V.M. All trans retinoic acid and cancer. Immunopharmacol. Immunotoxicol. 2011, 33, 241–249. [Google Scholar] [CrossRef]
- Niles, R.M. Signaling pathways in retinoid chemoprevention and treatment of cancer. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 555, 97–105. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of vitamin A in the immune system. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan-Harshman, M.; Aldoori, W. Vitamin B12 and health. Can. Fam. Physician 2008, 54, 536–541. [Google Scholar] [PubMed]
- Vitamin B12: Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/VitaminB12-HealthProfessional/ (accessed on 14 March 2022).
- Paul, C.; Brady, D.M. Comparative Bioavailability and Utilization of Particular Forms of B12 Supplements with Potential to Mitigate B12-related Genetic Polymorphisms. Integr. Med. 2017, 16, 42–49. [Google Scholar]
- Weissbach, H.; Dickerman, H. Biochemical Role of Vitamin B12. Physiol. Rev. 1965, 45, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Hammond, N.; Wang, Y.; Dimachkie, M.M.; Barohn, R.J. Nutritional neuropathies. Neurol. Clin. 2013, 31, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberg, A.J.; Selhub, J.; Shah, K.V.; Viscidi, R.P.; Comstock, G.W.; Helzlsouer, K.J. The risk of cervical cancer in relation to serum concentrations of folate, vitamin B12, and homocysteine. Cancer Epidemiol. Biomark. Prev. 2000, 9, 761–764. [Google Scholar]
- Ames, B.N. DNA damage from micronutrient deficiencies is likely. Univ. Calif. Berkeley 2001, 475, 7–20. [Google Scholar]
- Ikehata, M.; Ueda, K.; Iwakawa, S. Different involvement of DNA methylation and histone deacetylation in the expression of solute-carrier transporters in 4 colon cancer cell lines. Biol. Pharm. Bull. 2012, 35, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, D.; Minamishima, Y.A.; Nakayama, K. Prolyl-hydroxylase PHD3 interacts with pyruvate dehydrogenase (PDH)-E1β and regulates the cellular PDH activity. Biochem. Biophys. Res. Commun. 2014, 451, 288–294. [Google Scholar] [CrossRef]
- Lương, K.V.Q.; Nguyễn, L.T.H. Genetic and Cellular Signaling Mechanisms The Role of Thiamine in Cancer: Possible. Cancer Genom. Proteom. 2013, 10, 169–186. [Google Scholar] [CrossRef]
- Yonashiro, R.; Eguchi, K.; Wake, M.; Takeda, N.; Nakayama, K. Pyruvate dehydrogenase PDH-E1b controls tumor progression by altering the metabolic status of cancer cells. Cancer Res. 2018, 78, 1592–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrax, J.; Buc Calderon, P. The controversial place of vitamin C in cancer treatment. Biochem. Pharmacol. 2008, 76, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Roa, F.J.; Peña, E.; Gatica, M.; Escobar-Acuña, K.; Saavedra, P.; Maldonado, M.; Cuevas, M.E.; Moraga-Cid, G.; Rivas, C.I.; Muñoz-Montesino, C. Therapeutic Use of Vitamin C in Cancer: Physiological Considerations. Front. Pharmacol. 2020, 11, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, H.; Flower, G.; Weeks, L.; Cooley, K.; Callachan, M.; McGowan, J.; Skidmore, B.; Kirchner, L.; Seely, D. Intravenous vitamin C and cancer: A systematic review. Integr. Cancer Ther. 2014, 13, 280–300. [Google Scholar] [CrossRef] [PubMed]
- Gillberg, L.; Ørskov, A.D.; Liu, M.; Harsløf, L.B.S.; Jones, P.A.; Grønbæk, K. Vitamin C—A new player in regulation of the cancer epigenome. Semin. Cancer Biol. 2018, 51, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Cook, J. Intravenous vitamin C for cancer therapy—Identifying the current gaps in our knowledge. Front. Physiol. 2018, 9, 1182. [Google Scholar] [CrossRef]
- Park, S.; Ahn, S.; Shin, Y.; Yang, Y.; Yeom, C.H. Vitamin C in cancer: A metabolomics perspective. Front. Physiol. 2018, 9, 762. [Google Scholar] [CrossRef] [Green Version]
- Vissers, M.C.M.; Das, A.B. Potential mechanisms of action for vitamin C in cancer: Reviewing the evidence. Front. Physiol. 2018, 9, 809. [Google Scholar] [CrossRef] [Green Version]
- Pawlowska, E.; Szczepanska, J.; Blasiak, J. Pro- and antioxidant effects of Vitamin C in cancer in correspondence to its dietary and pharmacological concentrations. Oxid. Med. Cell. Longev. 2019, 2019, 7286737. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Li, Y.; Gong, Y.; Huang, Q.; Cai, S.; Peng, J. Vitamin D Status and Survival in Stage II-III Colorectal Cancer. Front. Oncol. 2020, 10, 581597. [Google Scholar] [CrossRef]
- Jeon, S.M.; Shin, E.A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmijewski, M.A. Vitamin D and human health. Int. J. Mol. Sci. 2019, 20, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mena, J.M.O.; Brenner, H. Vitamin D and cancer: An overview on epidemiological studies. Adv. Exp. Med. Biol. 2014, 810, 17–32. [Google Scholar]
- Szkandera, J.; Absenger, G.; Pichler, M.; Stotz, M.; Langsenlehner, T.; Samonigg, H.; Renner, W.; Gerger, A. Association of common gene variants in vitamin D modulating genes and colon cancer recurrence. J. Cancer Res. Clin. Oncol. 2013, 139, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, C.; Hao, M.; Sun, R.; Wang, Y.; Liu, T.; Cong, X.; Liu, Y. CYP24A1 is a potential biomarker for the progression and prognosis of human colorectal cancer. Hum. Pathol. 2016, 50, 101–108. [Google Scholar] [CrossRef]
- Yang, C.S.; Suh, N.; Kong, A.N.T. Does vitamin E prevent or promote cancer? Cancer Prev. Res. 2012, 5, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Kanchi, M.M.; Shanmugam, M.K.; Rane, G.; Sethi, G.; Kumar, A.P. Tocotrienols: The unsaturated sidekick shifting new paradigms in vitamin E therapeutics. Drug Discov. Today 2017, 22, 1765–1781. [Google Scholar] [CrossRef]
- Constantinou, C.; Charalambous, C.; Kanakis, D. Vitamin E and cancer: An update on the emerging role of γ and δ tocotrienols. Eur. J. Nutr. 2020, 59, 845–857. [Google Scholar] [CrossRef]
- Peh, H.Y.; Tan, W.S.D.; Liao, W.; Wong, W.S.F. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E as effective agents for cancer prevention and therapy. Adv. Nutr. 2017, 8, 850–867. [Google Scholar] [CrossRef]
- Chang, P.N.; Yap, W.N.; Wing Lee, D.T.; Ling, M.T.; Wong, Y.C.; Yap, Y.L. Evidence of -tocotrienol as an apoptosis-inducing, invasion-suppressing, and chemotherapy drug-sensitizing agent in human melanoma cells. Nutr. Cancer 2009, 61, 357–366. [Google Scholar] [CrossRef]
- Juanola-Falgarona, M.; Salas-Salvadó, J.; Martínez-Gonzaĺez, M.A.; Corella, D.; Estruch, R.; Ros, E.; Fitó, M.; Arós, F.; Gómez-Gracia, E.; Fiol, M.; et al. Dietary intake of vitamin K is inversely associated with mortality risk. J. Nutr. 2014, 144, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Dahlberg, S.; Ede, J.; Schött, U. Vitamin K and cancer. Scand. J. Clin. Lab. Investig. 2017, 77, 555–567. [Google Scholar] [CrossRef]
- Nimptsch, K.; Rohrmann, S.; Kaaks, R.; Linseisen, J. Dietary vitamin K intake in relation to cancer incidence and mortality: Results from the Heidelberg cohort of the European Prospective Investigation into Cancer and Nutrition (EPIC-Heidelberg). Am. J. Clin. Nutr. 2010, 91, 1348–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postles, J.; Curtis, T.Y.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Halford, N.G. Changes in free amino acid concentration in rye grain in response to nitrogen and sulfur availability, and expression analysis of genes involved in asparagine metabolism. Front. Plant Sci. 2016, 7, 917. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, D.M. Twenty-five years of mTOR: Uncovering the link from nutrients to growth. Proc. Natl. Acad. Sci. USA 2017, 114, 11818–11825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine bioavailability governs metastasis in a model of breast cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Lorenzi, P.L.; Reinhold, W.C.; Rudelius, M.; Gunsior, M.; Shankavaram, U.; Bussey, K.J.; Scherf, U.; Eichler, G.S.; Martin, S.E.; Chin, K.; et al. Asparagine synthetase as a causal, predictive biomarker for L-asparaginase activity in ovarian cancer cells. Mol. Cancer Ther. 2006, 5, 2613–2623. [Google Scholar] [CrossRef] [Green Version]
- Brumano, L.P.; da Silva, F.V.; Costa-Silva, T.A.; Apolinário, A.C.; Santos, J.H.; Kleingesinds, E.K.; Monteiro, G.; Rangel-Yagui, C.D.; Benyahia, B.; Junior, A.P. Development of L-asparaginase biobetters: Current research status and review of the desirable quality profiles. Front. Bioeng. Biotechnol. 2019, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Bermudez, J.; Williams, R.T.; Guarecuco, R.; Birsoy, K. Targeting extracellular nutrient dependencies of cancer cells. Mol. Metab. 2020, 33, 67–82. [Google Scholar] [CrossRef]
- Clarkson, B.; Krakoff, I.; Burchenal, J.; Karnofsky, D.; Golby, R.; Dowling, M.; Oettegen, H.; Lipton, A. Clinical Results of Treatment with E. Coli L-Asparaginase in Adults with Leukemia, Lymphoma, and Solid Tumors. Cancer 1969, 25, 279–305. [Google Scholar] [CrossRef]
- Su, N.; Pan, Y.-X.; Zhou, M.; Harvey, R.; Hunger, S.; Kilberg, M. Correlation between Asparaginase Sensitivity and Asparagine Synthetase Protein Content, but not mRNA, in Acute Lymphoblastic Leukemia Cell Lines. Pediatr. Blood Cancer 2008, 50, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.K.; Horvath, T.D.; Tan, L.; Link, T.; Harutyunyan, K.G.; Pontikos, M.A.; Anishkin, A.; Du, D.; Martin, L.A.; Yin, E.; et al. Glutaminase activity of L-asparaginase contributes to durable preclinical activity against acute lymphoblastic leukemia. Mol. Cancer Ther. 2019, 18, 1587–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlozkova, K.; Pecinova, A.; Alquezar-Artieda, N.; Pajuelo-Reguera, D.; Simcikova, M.; Hovorkova, L.; Rejlova, K.; Zaliova, M.; Mracek, T.; Kolenova, A.; et al. Metabolic profile of leukemia cells influences treatment efficacy of L-asparaginase. BMC Cancer 2020, 20, 526. [Google Scholar] [CrossRef] [PubMed]
- Albaugh, V.L.; Pinzon-Guzman, C.; Barbul, A. Arginine Metabolism and Cancer. J. Surg. Oncol. 2017, 115, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.Y.; Wang, Y.Y.; Jing, Y.; Xu, M.H.; Zhu, Z.T.; Wang, Q.J. Abnormal arginine metabolism is associated with prognosis in patients of gastric cancer. Transl. Cancer Res. 2021, 10, 2451–2469. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Gao, Y.; Cao, Y.; Zhang, Y.; Xu, M.; Wang, Y.; Jing, Y.; Guo, S.; Jing, F.; Hu, X.; et al. Association of plasma arginine with breast cancer molecular subtypes in women of Liaoning province. IUBMB Life 2016, 68, 980–984. [Google Scholar] [CrossRef]
- Mirmiran, P.; Moghadam, S.K.; Bahadoran, Z.; Ghasemi, A.; Azizi, F. Dietary L-Arginine Intakes and the Risk of Metabolic Syndrome: A 6-Year Follow-Up in Tehran Lipid and Glucose Study. Prev. Nutr. Food Sci. 2017, 22, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Izzo, F.; Marra, P.; Beneduce, G.; Castello, G.; Vallone, P.; De Rosa, V.; Cremona, F.; Ensor, C.M.; Holtsberg, F.W.; Bomalaski, J.S.; et al. Pegylated arginine deiminase treatment of patients with unresectable hepatocellular carcinoma: Results from phase I/II studies. J. Clin. Oncol. 2004, 22, 1815–1822. [Google Scholar] [CrossRef]
- Kim, S.K.; Jung, W.H.; Koo, J.S. Differential expression of enzymes associated with serine/glycine metabolism in different breast cancer subtypes. PLoS ONE 2014, 9, e101004. [Google Scholar] [CrossRef]
- Perry, C.; Yu, S.; Chen, J.; Matharu, K.S.; Stover, P.J. Effect of vitamin B6 availability on serine hydroxymethyltransferase in MCF-7 cells. Arch. Biochem. Biophys. 2007, 462, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolfi, S.C.; Chan, L.L.-Y.; Qiu, J.; Tedeschi, P.M.; Bertino, J.R.; Hirshfield, K.M.; Oltvai, Z.N.; Vazquez, A. The metabolic demands of cancer cells are coupled to their size and protein synthesis rates. Cancer Metab. 2013, 1, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locasale, J.W. Serine, Glycine and the one-carbon cycle. Nat. Rev Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajan, M.; Hennequart, M.; Cheung, E.C.; Zani, F.; Hock, A.K.; Legrave, N.; Maddocks, O.D.K.; Ridgway, R.A.; Athineos, D.; Suárez-Bonnet, A.; et al. Serine synthesis pathway inhibition cooperates with dietary serine and glycine limitation for cancer therapy. Nat. Commun. 2021, 12, 366. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Asai, A.; Konno, M.; Koseki, J.; Taniguchi, M.; Vecchione, A.; Ishii, H. One-carbon metabolism for cancer diagnostic and therapeutic approaches. Cancer Lett. 2020, 470, 141–148. [Google Scholar] [CrossRef]
- Banjac, A.; Perisic, T.; Sato, H.; Seiler, A.; Bannai, S.; Weiss, N.; Kölle, P.; Tschoep, K.; Issels, R.D.; Daniel, P.T.; et al. The cystine/cysteine cycle: A redox cycle regulating susceptibility versus resistance to cell death. Oncogene 2008, 27, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Lewerenz, J.; Hewett, S.J.; Huang, Y.; Lambros, M.; Gout, P.W.; Kalivas, P.W.; Massie, A.; Smolders, I.; Methner, A.; Pergande, M.; et al. The cystine/glutamate antiporter system xc- in health and disease: From molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 522–555. [Google Scholar] [CrossRef] [Green Version]
- Novera, W.; Lee, Z.W.; Nin, D.S.; Dai, M.Z.Y.; Binte Idres, S.; Wu, H.; Damen, J.M.A.; Tan, T.Z.; Sim, A.Y.L.; Long, Y.C.; et al. Cysteine Deprivation Targets Ovarian Clear Cell Carcinoma Via Oxidative Stress and Iron-Sulfur Cluster Biogenesis Deficit. Antioxid. Redox Signal. 2020, 33, 1191–1208. [Google Scholar] [CrossRef]
- Cervantes-Villagrana, R.D.; Albores-García, D.; Cervantes-Villagrana, A.R.; García-Acevez, S.J. Tumor-induced neurogenesis and immune evasion as targets of innovative anti-cancer therapies. Signal Transduct. Target. Ther. 2020, 5, 99. [Google Scholar] [CrossRef]
- Brown, I.S. Pathology of Perineural Spread. J. Neurol. Surg. Part B Skull Base 2016, 77, 124–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandforth, L.; Ammar, N.; Dinges, L.A.; Röcken, C.; Arlt, A.; Sebens, S.; Schäfer, H. Impact of the Monocarboxylate transporter-1 (MCT1)-mediated cellular import of lactate on stemness properties of human pancreatic adenocarcinoma cells. Cancers 2020, 12, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Liang, J.; Duan, J.; Chen, L.; Li, H.; Zhen, T.; Zhang, F.; Dong, Y.; Shi, H.; Han, A. A Prognosis Marker SLC2A3 Correlates With EMT and Immune Signature in Colorectal Cancer. Front. Oncol. 2021, 11, 638099. [Google Scholar] [CrossRef] [PubMed]
- Banh, R.S.; Biancur, D.E.; Yamamoto, K.; Sohn, A.S.W.; Walters, B.; Kuljanin, M.; Gikandi, A.; Wang, H.; Mancias, J.D.; Schneider, R.J.; et al. Neurons Release Serine to Support mRNA Translation in Pancreatic Cancer. Cell 2020, 183, 1202–1218.e25. [Google Scholar] [CrossRef] [PubMed]
- El Soury, M.; Fornasari, B.E.; Carta, G.; Zen, F.; Haastert-Talini, K.; Ronchi, G. The role of dietary nutrients in peripheral nerve regeneration. Int. J. Mol. Sci. 2021, 22, 7417. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Klaassen, C. Tissue distribution and thyroid hormone regulation of Pept1 and Pept2 mRNA in rodents. Peptides 2006, 27, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Jung, S.; Park, W.S.; Lee, J.H.; Shin, R.; Heo, S.C.; Choe, E.K.; Lee, J.H.; Kim, K.; Chai, Y.J. Upregulation of SLC2A3 gene and prognosis in colorectal carcinoma: Analysis of TCGA data. BMC Cancer 2019, 19, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Hong, J.; Han, H.; Park, J.; Kim, D.; Park, H.; Ko, M.; Koh, Y.; Shin, D.Y.; Yoon, S.S. Decreased vitamin C uptake mediated by SLC2A3 promotes leukaemia progression and impedes TET2 restoration. Br. J. Cancer 2020, 122, 1445–1452. [Google Scholar] [CrossRef]
- Sprute, R.; Ardicli, D.; Oguz, K.K.; Malenica-Mandel, A.; Daimagüler, H.S.; Koy, A.; Coskun, T.; Wang, H.; Topcu, M.; Cirak, S. Clinical outcomes of two patients with a novel pathogenic variant in ASNS: Response to asparagine supplementation and review of the literature. Hum. Genome Var. 2019, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Ellerbroek, S.M.; Wennerberg, K.; Burridge, K. Serine phosphorylation negatively regulates RhoA in vivo. J. Biol. Chem. 2003, 278, 19023–19031. [Google Scholar] [CrossRef] [Green Version]
- Nusser, N.; Gosmanova, E.; Makarova, N.; Fujiwara, Y.; Yang, L.; Guo, F.; Luo, Y.; Zheng, Y.; Tigyi, G. Serine phosphorylation differentially affects RhoA binding to effectors: Implications to NGF-induced neurite outgrowth. Cell. Signal. 2006, 18, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Too, H.P. Mitochondrial localized STAT3 is involved in NGF induced neurite outgrowth. PLoS ONE 2011, 6, e21680. [Google Scholar] [CrossRef] [PubMed]
- Tapia, J.C.; Mentis, G.Z.; Navarrete, R.; Nualart, F.; Figueroa, E.; Sánchez, A.; Aguayo, L.G. Early expression of glycine and GABAA receptors in developing spinal cord neurons. Effects on neurite outgrowth. Neuroscience 2001, 108, 493–506. [Google Scholar] [CrossRef]
- El Hayek, L.; Khalifeh, M.; Zibara, V.; Abi Assaad, R.; Emmanuel, N.; Karnib, N.; El-Ghandour, R.; Nasrallah, P.; Bilen, M.; Ibrahim, P.; et al. Lactate mediates the effects of exercise on learning and memory through sirt1-dependent activation of hippocampal brain-derived neurotrophic factor (BDNF). J. Neurosci. 2019, 39, 2369–2382. [Google Scholar] [CrossRef] [Green Version]
- Lezi, E.; Lu, J.; Selfridge, J.E.; Burns, J.M.; Swerdlow, R.H. Lactate administration reproduces specific brain and liver exercise-related changes. J. Neurochem. 2013, 127, 91–100. [Google Scholar] [CrossRef] [Green Version]
- García-Suárez, P.C.; Rentería, I.; Plaisance, E.P.; Moncada-Jiménez, J.; Jiménez-Maldonado, A. The effects of interval training on peripheral brain derived neurotrophic factor (BDNF) in young adults: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 8937. [Google Scholar] [CrossRef]
- Bakst, R.L.; Glastonbury, C.M.; Parvathaneni, U.; Katabi, N.; Hu, K.S.; Yom, S.S. Perineural Invasion and Perineural Tumor Spread in Head and Neck Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2019, 103, 1109–1124. [Google Scholar] [CrossRef] [Green Version]
- Jha, M.K.; Morrison, B.M. Lactate Transporters Mediate Glia-Neuron Metabolic Crosstalk in Homeostasis and Disease. Front. Cell. Neurosci. 2020, 14, 589582. [Google Scholar] [CrossRef]
- Lin, H.; Muramatsu, R.; Maedera, N.; Tsunematsu, H.; Hamaguchi, M.; Koyama, Y.; Kuroda, M.; Ono, K.; Sawada, M.; Yamashita, T. Extracellular Lactate Dehydrogenase A Release From Damaged Neurons Drives Central Nervous System Angiogenesis. EBioMedicine 2018, 27, 71–85. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, M.R.; Da Rocha, R.F.; Stertz, L.; Fries, G.R.; De Oliveira, D.L.; Kapczinski, F.; Moreira, J.C.F. Total and mitochondrial nitrosative stress, decreased brain- Derived neurotrophic factor (BDNF) levels and glutamate uptake, and evidence of endoplasmic reticulum stress in the hippocampus of vitamin A-treated rats. Neurochem. Res. 2011, 36, 506–517. [Google Scholar] [CrossRef]
- Páez Pereda, M.; Missale, C.; Grübler, Y.; Arzt, E.; Schaaf, L.; Stalla, G.K. Nerve growth factor and retinoic acid inhibit proliferation and invasion in thyroid tumor cells. Mol. Cell. Endocrinol. 2000, 167, 99–106. [Google Scholar] [CrossRef]
- Arrieta, O.; Hernández-Pedro, N.; Fernández-González-Aragón, M.C.; Saavedra-Pérez, D.; Campos-Parra, A.D.; Ríos-Trejo, M.A.; Cerón-Lizárraga, T.; Martínez-Barrera, L.; Pineda, B.; Ordóñez, G.; et al. Retinoic acid reduces chemotherapyinduced neuropathy in an animal model and patients with lung cancer. Neurology 2011, 77, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Higashi, M.; Kolla, V.; Iyer, R.; Naraparaju, K.; Zhuang, T.; Kolla, S.; Brodeur, G.M. Retinoic acid-induced CHD5 upregulation and neuronal differentiation of neuroblastoma. Mol. Cancer 2015, 14, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, L.M.; Danks, M.K. New therapeutic targets for the treatment of high-risk neuroblastoma. J. Cell. Biochem. 2009, 107, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, P.; Park, H.; Baumann, M.; Dunlop, J.; Frydman, J.; Kopito, R.; McCampbell, A.; Leblanc, G.; Venkateswaran, A.; Nurmi, A.; et al. Protein misfolding in neurodegenerative diseases: Implications and strategies. Transl. Neurodegener. 2017, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, O.; Koshy, S.T.; Branco Da Cunha, C.; Shin, J.W.; Verbeke, C.S.; Allison, K.H.; Mooney, D.J. Extracellular matrix stiffness and composition jointly regulate the induction of malignant phenotypes in mammary epithelium. Nat. Mater. 2014, 13, 970–978. [Google Scholar] [CrossRef]
- Xu, C.J.; Wang, J.L.; Jin, W.L. The Emerging Therapeutic Role of NGF in Alzheimer’s Disease. Neurochem. Res. 2016, 41, 1211–1218. [Google Scholar] [CrossRef]
- Ina, A.; Kamei, Y. Vitamin B12, a chlorophyll-related analog to pheophytin a from marine brown algae, promotes neurite outgrowth and stimulates differentiation in PC12 cells. Cytotechnology 2006, 52, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Tanaka, H.; Temporin, K.; Okamoto, M.; Kuroda, Y.; Moritomo, H.; Murase, T.; Yoshikawa, H. Methylcobalamin increases Erk1/2 and Akt activities through the methylation cycle and promotes nerve regeneration in a rat sciatic nerve injury model. Exp. Neurol. 2010, 222, 191–203. [Google Scholar] [CrossRef]
- Segal, R.A. Selectivity in neurotrophin signaling: Theme and variations. Annu. Rev. Neurosci. 2003, 26, 299–330. [Google Scholar] [CrossRef]
- Markus, A.; Zhong, J.; Snider, W.D. Raf and Akt mediate distinct aspects of sensory axon growth. Neuron 2002, 35, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Tanaka, H.; Temporin, K.; Okamoto, M.; Kuroda, Y.; Moritomo, H.; Murase, T.; Yoshikawa, H. Akt/mammalian target of rapamycin signaling pathway regulates neurite outgrowth in cerebellar granule neurons stimulated by methylcobalamin. Neurosci. Lett. 2011, 495, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Baltrusch, S. The Role of Neurotropic B Vitamins in Nerve Regeneration. Biomed Res. Int. 2021, 2021, 9968228. [Google Scholar] [CrossRef] [PubMed]
- Al-saaeed, S.M.; Al-khalisy, M.H. The Regenerative Role of Vitamins B1, B6, B12 in Treatment of Peripheral Neuropathy. Int. J. Sci. Res. 2015, 6, 2319–7064. [Google Scholar] [CrossRef]
- Schloss, J.; Colosimo, M. B Vitamin Complex and Chemotherapy-Induced Peripheral Neuropathy. Curr. Oncol. Rep. 2017, 19, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Altun, I.; Kurutaş, E.B. Vitamin B complex and vitamin B12 levels after peripheral nerve injury. Neural Regen. Res. 2016, 11, 842–845. [Google Scholar] [CrossRef]
- Wrzosek, M.; Lukaszkiewicz, J.; Wrzosek, M.; Jakubczyk, A.; Matsumoto, H.; Piatkiewicz, P.; Radziwon-Zaleska, M.; Wojnar, M.; Nowicka, G. Vitamin D and the central nervous system. Pharmacol. Rep. 2013, 65, 271–278. [Google Scholar] [CrossRef]
- Deluca, G.C.; Kimball, S.M.; Kolasinski, J.; Ramagopalan, S.V.; Ebers, G.C. Review: The role of vitamin D in nervous system health and disease. Neuropathol. Appl. Neurobiol. 2013, 39, 458–484. [Google Scholar] [CrossRef]
- Annweiler, C.; Schott, A.M.; Berrut, G.; Chauviré, V.; Le Gall, D.; Inzitari, M.; Beauchet, O. Vitamin D and ageing: Neurological issues. Neuropsychobiology 2010, 62, 139–150. [Google Scholar] [CrossRef]
- Eyles, D.; Brown, J.; Mackay-Sim, A.; McGrath, J.; Feron, F. Vitamin D3 and brain development. Neuroscience 2003, 118, 641–653. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienol: The natural vitamin E to defend the nervous system? Ann. N. Y. Acad. Sci. 2004, 1031, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Morani, A.S.; Bodhankar, S.L. Neuroprotective effect of vitamin E acetate in models of mononeuropathy in rats. Neuroanatomy 2008, 7, 33–37. [Google Scholar]
- Pace, A.; Savarese, A.; Picardo, M.; Maresca, V.; Pacetti, U.; Del Monte, G.; Biroccio, A.; Leonetti, C.; Jandolo, B.; Cognetti, F.; et al. Neuroprotective effect of vitamin E supplementation in patients treated with cisplatin chemotherapy. J. Clin. Oncol. 2003, 21, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.K.; Kamei, Y. Novel effect of vitamin K1 (phylloquinone) and vitamin K2 (menaquinone) on promoting nerve growth factor-mediated neurite outgrowth from PC12D cells. Neurosci. Lett. 2002, 323, 9–12. [Google Scholar] [CrossRef]
- Graham, D.K.; Deryckere, D.; Davies, K.D.; Earp, H.S. The TAM family: Phosphatidylserine-sensing receptor tyrosine kinases gone awry in cancer. Nat. Rev. Cancer 2014, 14, 769–785. [Google Scholar] [CrossRef]
- Shafit-Zagardo, B.; Gruber, R.C.; DuBois, J.C. The role of TAM family receptors and ligands in the nervous system: From development to pathobiology. Pharmacol. Ther. 2018, 188, 97–117. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Metabolite | Cancer Type | Role in Tumor Progression |
|---|---|---|
| Vitamin A | Breast | 4-HPR induces cell death [37]; vitamin A and retinol reduce risk [105] |
| Colon/Colorectal | 4-HPR induces cell death [37] | |
| Head/Neck | 4-HPR induces cell death [37] | |
| Gastric | Inhibits polycyclic hydrocarbon-induced carcinomas [37,108] | |
| Lung | Blood levels of α- and β-carotene, total carotenoids, and retinol are inversely associated with cancer risk [109] | |
| Prostate | 4-HPR induces cell death [37] | |
| Vitamin B1 | Breast | Intermediate concentrations promote Ehrlich’s ascites proliferation in thiamine-deficient patients; high concentrations inhibit proliferation [110]; patients exhibit decreased expression of SLC9A3 transporter gene [111,112] |
| Head/Neck | Patients exhibit decreased expression of SLC9A3 transporter gene [112] Intake reduces risk of esophageal cancer [103] | |
| Vitamin B3 | Head/Neck | Intake reduces risk of esophageal cancer [103] |
| Vitamin B6 | Head/Neck | Intake reduces risk of esophageal cancer [103] |
| Vitamin B9 | Head/Neck | Intake reduces risk of esophageal cancer [103] |
| Vitamin B12 | Head/Neck | Intake increases risk of esophageal cancer [103] |
| Leukemia/Lymphoma | Elevated plasma levels associated with 1-year cancer risk [113] | |
| Liver | Elevated plasma levels associated with 1-year cancer risk [113] | |
| Lung | Positively associated with cancer risk in dose-dependent manner [114] | |
| Pancreatic | Elevated plasma levels associated with 1-year cancer risk [113] | |
| Vitamin C | Breast | Low concentrations induce cell invasiveness; high doses restrict EMT [115] |
| Skin | Low doses reduce cell viability and invasiveness; high doses promote cell migration [116] | |
| Vitamin D | Breast | Calcitroil and D3 analogs suppress MMP-2 and -9 and VCAM-1; low serum D3 levels are associated with high incidence [117] |
| Colon/Colorectal | Low serum D3 levels are associated with high incidence [117]; analog PRI-2191 enhances ability of 5-FU to restrict cell cycle [118]; serum levels of ≥33 ng/mL correlates with a 50% decreased risk [119] | |
| Gastric | Low serum D3 levels associated with high incidence [117] | |
| Head/Neck | MART-10 induces cell-cycle arrest and suppresses p21 and p27 [120] | |
| Prostate | Lower serum levels are associated with an increased risk; D3 and analogs inhibit invasiveness and expression of MMP-2 and -9 and VCAM-1 [117] | |
| Vitamin E | Breast | Tocotrienols exhibit chemotherapeutic and antitumor properties [41]; γ-tocotrienol induces cell-cycle arrest [121,122] |
| Colon/Colorectal | Tocotrienols exhibit antitumor properties [41]; γ-tocotrienol mediates apoptosis via apoptotic caspase-3 and NFκB suppression [123] | |
| Liver | Tocotrienols exhibit chemotherapeutic properties [41] | |
| Lung | Tocotrienols exhibit chemotherapeutic properties [41] | |
| Pancreatic | Tocotrienols exhibit chemotherapeutic properties [41] | |
| Prostate | Tocotrienols exhibit chemotherapeutic properties [41] | |
| Vitamin K | Breast | K2 induces nonapoptotic cell death [42] |
| Arginine | Breast | Low plasma levels act as a prognostic biomarker [124]; arginine starvation is used to treat arginosuccinate synthase-deficient patients [101,124] |
| Ovarian | Cancer cells are deficient in arginosuccinate synthase-1; ADI-PED-20 is used to degrade arginine [125] | |
| Prostate | Low plasma levels act as a prognostic biomarker [126] | |
| Skin | Cells are deficient in arginosuccinate synthase-1; ADI-PEG20-resistant cancer exhibits c-MYC binding to the promoter of arginosuccinate synthase-1 [26] | |
| Asparagine | Breast | Maintains health of glutamine-independent cells [96] |
| Cervical | Facilitates mTOR activation in the absence of glutamine [96] | |
| Liposarcoma | Maintains health of glutamine-independent cells [96] | |
| Cysteine | Breast | Inhibition of histone deacetylase-6 sensitizes TNBC cells to cysteine deprivation via cystine/glutamate antiporter-targeted therapies [98] |
| Colon/Colorectal | Starvation induces a reduction in liver-metastatic cell proliferation [29] | |
| Pancreatic | The deletion of cystine transporter gene SLC7A11 inhibits autophagy and diminishes cysteine homeostasis [127] | |
| Glycine | Colon/Colorectal | Metabolism increases when starved of serine [128]; serine–glycine inhibition should be used in conjunction to decrease tumor size [102] |
| Lung | De novo serine and glycine are allocated to glutathione synthesis [128] | |
| Lactate | Breast | 10 mM L-lactate acts as chemoattractant and facilitates migration [129]; intermediate and high supplementation upregulates oncogene, proliferation gene, tumor suppressor, and transcription-factor expression [130]; |
| Cervical | When given glucose and lactate, oxidative cancer cells prefer lactate; cells thrive when given lactate supplementation; oxidative cells exhibit high expression of MCT1 versus MCT4; MCT1 inhibition induces necrosis in oxidative cells [25]; DLAD targets metabolism [131] | |
| Colon/Colorectal | Glycolytic cells fail to thrive upon glucose starvation with lactate supplementation [25] | |
| Head/Neck | DLAD targets metabolism [131] | |
| Lung | NFκB signaling [132]; LDH-A inhibition sensitizes cells to radiation [133] | |
| Skin | DLAD targets metabolism [131] | |
| Serine | Breast | Cells prefer serine over glycine and exhibit a decrease in nucleic acid synthesis when starved of serine [99]; brain metastatic cells upregulate de novo serine when starved of exogenous alternative [134] |
| Colon/Colorectal | Cells prefer serine over glycine and exhibit a decrease in nucleic acid synthesis when starved of serine [99]; metabolism increases when starved of serine; starvation decreases YAP activation [135]; serine–glycine catabolism induced by stress promotes formate production [136]; serine–glycine inhibition should be used in conjunction to decrease tumor size; starvation induces metabolic stress and p53-activated glycolysis [102] | |
| Lung | Promotes purine synthesis in cancer cells; de novo serine and glycine are allocated to glutathione synthesis [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, I.; Gregory, E.; Atwood, C.; Lee, S.-O.; Song, Y.H. Exploring the Role of Metabolites in Cancer and the Associated Nerve Crosstalk. Nutrients 2022, 14, 1722. https://doi.org/10.3390/nu14091722
Gu I, Gregory E, Atwood C, Lee S-O, Song YH. Exploring the Role of Metabolites in Cancer and the Associated Nerve Crosstalk. Nutrients. 2022; 14(9):1722. https://doi.org/10.3390/nu14091722
Chicago/Turabian StyleGu, Inah, Emory Gregory, Casey Atwood, Sun-Ok Lee, and Young Hye Song. 2022. "Exploring the Role of Metabolites in Cancer and the Associated Nerve Crosstalk" Nutrients 14, no. 9: 1722. https://doi.org/10.3390/nu14091722
APA StyleGu, I., Gregory, E., Atwood, C., Lee, S.-O., & Song, Y. H. (2022). Exploring the Role of Metabolites in Cancer and the Associated Nerve Crosstalk. Nutrients, 14(9), 1722. https://doi.org/10.3390/nu14091722