
Disproportionate Vitamin A Deficiency in Women of Specific Ethnicities Linked to Differences in Allele Frequencies of Vitamin A-Related Polymorphisms

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source and Study Population for Serum Retinol Levels
2.2. Genetic Variations of the Vitamin A-Related Gene Polymorphisms
2.3. Combinations of Low-Serum Retinol Risk Alleles
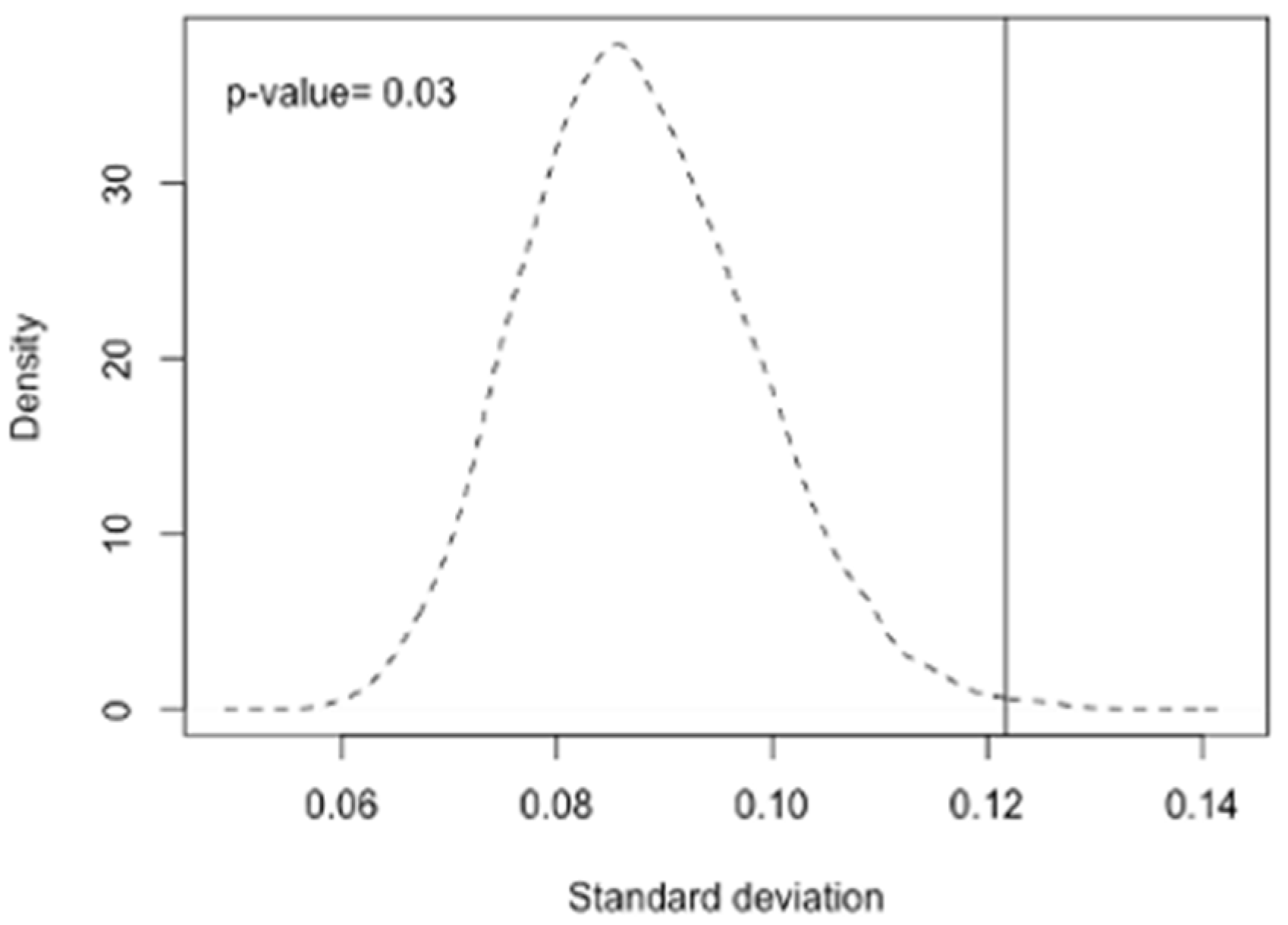
2.4. Statistics Analysis
3. Results
3.1. Differences in the Proportion of Vitamin A Deficiency in Pregnant Women between Different Ethnic Groups
3.1.1. The Bronx Study
3.1.2. NHANES
3.2. Genetic Variations and Differences of Vitamin A Deficiency Proportions
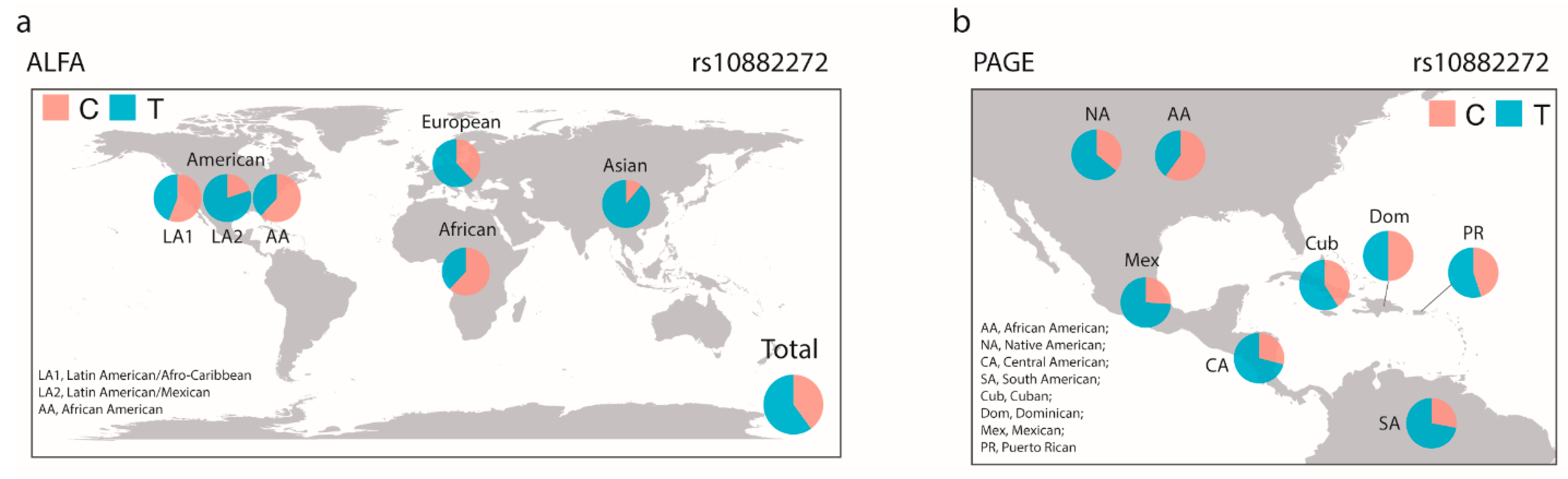
3.2.1. Differences of Allele Frequencies of Serum Retinol Levels Associated SNPs between Different Ethnic Groups
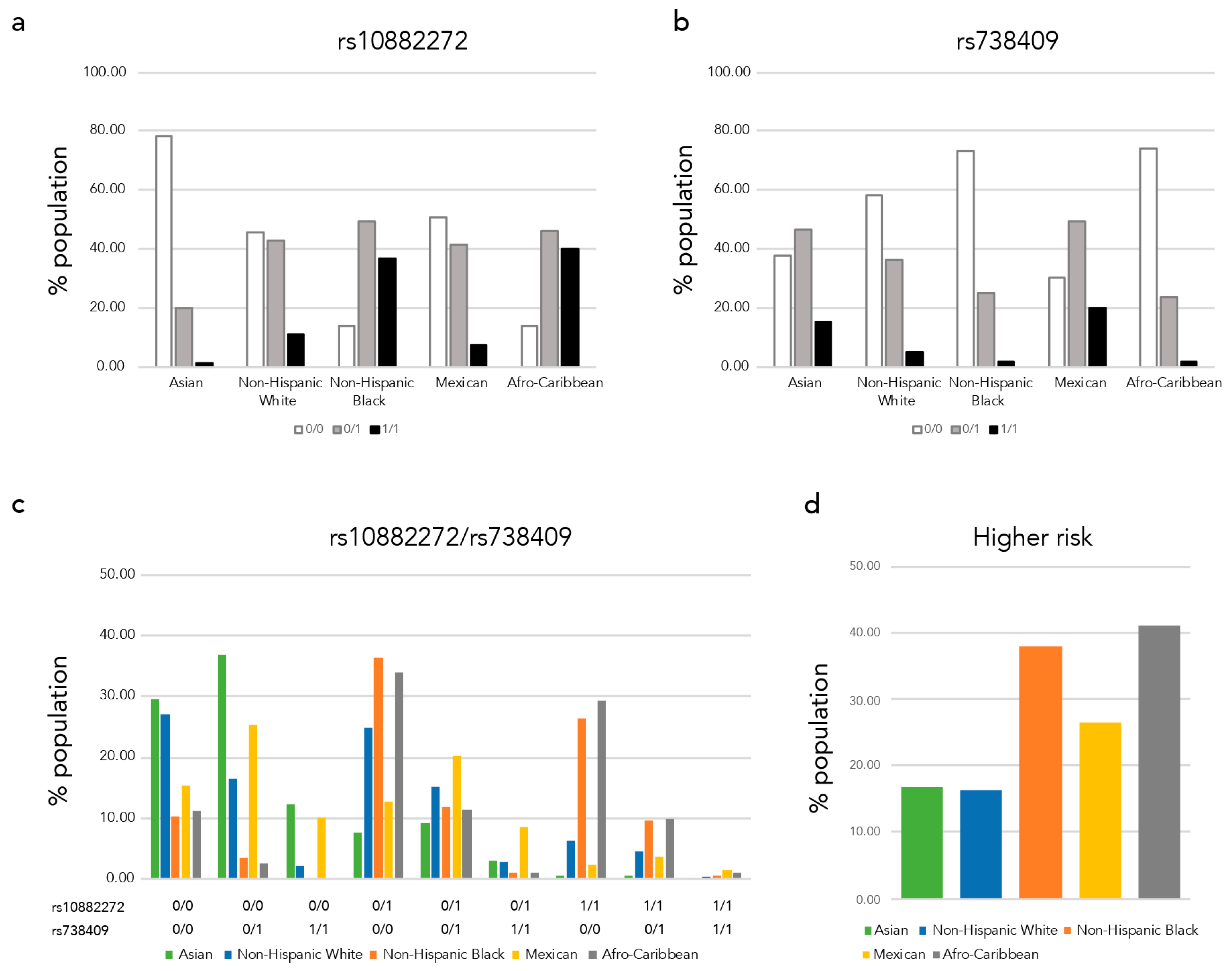
3.2.2. Proportions of Low-Serum Retinol Higher-Risk Populations Vary between Race/Ethnic Groups
3.2.3. Variations of Allele Frequencies of Polymorphisms Associated with Serum Retinol and Beta-Carotene Levels as Well as with the Beta-Carotene Bioactivities between Race/Ethnicity Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vogel, S.; Piantedosi, R.; O′Byrne, S.M.; Kako, Y.; Quadro, L.; Gottesman, M.E.; Goldberg, I.J.; Blaner, W.S. Retinol-binding protein-deficient mice: Biochemical basis for impaired vision. Biochemistry 2002, 41, 15360–15368. [Google Scholar] [CrossRef] [PubMed]
- Quadro, L.; Blaner, W.S.; Salchow, D.J.; Vogel, S.; Piantedosi, R.; Gouras, P.; Freeman, S.; Cosma, M.P.; Colantuoni, V.; Gottesman, M.E. Impaired retinal function and vitamin A availability in mice lacking retinol-binding protein. EMBO J. 1999, 18, 4633–4644. [Google Scholar] [CrossRef]
- Liu, X.; Cui, T.; Li, Y.; Wang, Y.; Wang, Q.; Li, X.; Bi, Y.; Wei, X.; Liu, L.; Li, T.; et al. Vitamin A supplementation in early life enhances the intestinal immune response of rats with gestational vitamin A deficiency by increasing the number of immune cells. PLoS ONE 2014, 9, e114934. [Google Scholar] [CrossRef]
- Sirisinha, S. The pleiotropic role of vitamin A in regulating mucosal immunity. Asian Pac. J. Allergy Immunol. 2015, 33, 71–89. [Google Scholar] [PubMed]
- Williams, J.B.; Napoli, J.L. Metabolism of retinoic acid and retinol during differentiation of F9 embryonal carcinoma cells. Proc. Natl. Acad. Sci. USA 1985, 82, 4658–4662. [Google Scholar] [CrossRef]
- Lee, T.F.; Mak, K.M.; Rackovsky, O.; Lin, Y.-L.; Kwong, A.J.; Loke, J.C.; Friedman, S.L. Downregulation of hepatic stellate cell activation by retinol and palmitate mediated by adipose differentiation-related protein (ADRP). J. Cell Physiol. 2010, 223, 648–657. [Google Scholar] [CrossRef]
- Cammas, L.; Romand, R.; Fraulob, V.; Mura, C.; Dollé, P. Expression of the murine retinol dehydrogenase 10 (Rdh10) gene correlates with many sites of retinoid signalling during embryogenesis and organ differentiation. Dev. Dyn. 2007, 236, 2899–2908. [Google Scholar] [CrossRef]
- Janick-Buckner, D.; Barua, A.B.; Olson, J.A. Induction of HL-60 cell differentiation by water-soluble and nitrogen-containing conjugates of retinoic acid and retinol. FASEB J. 1991, 5, 320–325. [Google Scholar] [CrossRef]
- Marx, J.; Naudé, H.; Pretorius, E. The Effects of Hypo- and Hypervitaminosis a and Its Involvement in Foetal Nervous System Development and Post-Natal Sensorimotor Functioning—A Review. Br. J. Dev. Disabil. 2006, 52, 47–64. [Google Scholar] [CrossRef]
- Zile, M.H. Vitamin A and embryonic development: An overview. J. Nutr. 1998, 128, 455S–458S. [Google Scholar] [CrossRef]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schulze, K.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)-Vitamin A Review. J. Nutr. 2016, 146, 1816S–1848S. [Google Scholar] [CrossRef]
- CDC. CDC′s Second Nutrition Report: A Comprehensive Biochemical Assessment of the Nutrition Status of the U.S. Population. Available online: https://www.cdc.gov/nutritionreport/pdf/4page_%202nd%20nutrition%20report_508_032912.pdf (accessed on 20 May 2020).
- WHO. Global Prevalence of Vitamin A Deficiency in Populations at Risk 1995–2005; WHO Global Database on Vitamin A Deficiency; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Schleicher, R.L.; Sternberg, M.R.; Pfeiffer, C.M. Race-ethnicity is a strong correlate of circulating fat-soluble nutrient concentrations in a representative sample of the U.S. population. J. Nutr. 2013, 143, 966S–976S. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.; Lyden, E.; Abresch, C.; Anderson-Berry, A. Serum retinol concentrations, race, and socioeconomic status in of women of childbearing age in the United States. Nutrients 2016, 8, 508. [Google Scholar] [CrossRef] [PubMed]
- Garretto, D.; Kim, Y.-K.; Quadro, L.; Rhodas, R.R.; Pimentel, V.; Crnosija, N.A.; Nie, L.; Bernstein, P.; Tropper, P.; Neal-Perry, G.S. Vitamin A and β-carotene in pregnant and breastfeeding post-bariatric women in an urban population. J. Perinat. Med. 2019, 47, 183–189. [Google Scholar] [CrossRef]
- Toprani, A.; Li, W.; Hadler, J.L. Trends in Mortality Disparities by Area-Based Poverty in New York City, 1990–2020. J. Urban Health 2016, 93, 538–550. [Google Scholar] [CrossRef] [PubMed]
- U.S. Census Bureau US Census. Available online: https://data.census.gov/cedsci/ (accessed on 13 July 2020).
- Hanson, C.; Schumacher, M.V.; Lyden, E.; Su, D.; Furtado, J.; Cammack, R.; Bereitschaft, B.; Van Ormer, M.; Needelman, H.; McGinn, E.; et al. Fat-soluble vitamins A and E and health disparities in a cohort of pregnant women at delivery. J. Nutr. Sci. 2018, 7, e14. [Google Scholar] [CrossRef]
- Borel, P.; Desmarchelier, C. Genetic Variations Associated with Vitamin A Status and Vitamin A Bioavailability. Nutrients 2017, 9, 246. [Google Scholar] [CrossRef]
- Pirazzi, C.; Valenti, L.; Motta, B.M.; Pingitore, P.; Hedfalk, K.; Mancina, R.M.; Burza, M.A.; Indiveri, C.; Ferro, Y.; Montalcini, T.; et al. PNPLA3 has retinyl-palmitate lipase activity in human hepatic stellate cells. Hum. Mol. Genet. 2014, 23, 4077–4085. [Google Scholar] [CrossRef]
- Mondul, A.M.; Yu, K.; Wheeler, W.; Zhang, H.; Weinstein, S.J.; Major, J.M.; Cornelis, M.C.; Männistö, S.; Hazra, A.; Hsing, A.W.; et al. Genome-wide association study of circulating retinol levels. Hum. Mol. Genet. 2011, 20, 4724–4731. [Google Scholar] [CrossRef]
- Allele Frequency Aggregator. Available online: https://www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 13 July 2020).
- Population Architecture Using Genomics and Epidemiology. Available online: https://www.pagestudy.org/index.php/home (accessed on 12 July 2020).
- 1000 Genomes Project Consortium; Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1092 human genomes. Nature 2012, 491, 56–65. [Google Scholar] [PubMed]
- Sorlie, P.D.; Avilés-Santa, L.M.; Wassertheil-Smoller, S.; Kaplan, R.C.; Daviglus, M.L.; Giachello, A.L.; Schneiderman, N.; Raij, L.; Talavera, G.; Allison, M.; et al. Design and implementation of the Hispanic Community Health Study/Study of Latinos. Ann. Epidemiol. 2010, 20, 629–641. [Google Scholar] [CrossRef] [PubMed]
- GTEx Consortium Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing, R.C.T. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 November 2020).
- Blumberg, J.B.; Frei, B.; Fulgoni, V.L.; Weaver, C.M.; Zeisel, S.H. Contribution of dietary supplements to nutritional adequacy by socioeconomic subgroups in adults of the united states. Nutrients 2017, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- The NHGRI-EBI Catalog of Human Genome-Wide Association Studies GWAS Catalog. Available online: https://www.ebi.ac.uk/gwas/ (accessed on 13 July 2020).
- Thompson, S.J.; Sargsyan, A.; Lee, S.-A.; Yuen, J.J.; Cai, J.; Smalling, R.; Ghyselinck, N.; Mark, M.; Blaner, W.S.; Graham, T.E. Hepatocytes are the principal source of circulating RBP4 in mice. Diabetes 2017, 66, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Kovarova, M.; Königsrainer, I.; Königsrainer, A.; Machicao, F.; Häring, H.-U.; Schleicher, E.; Peter, A. The Genetic Variant I148M in PNPLA3 Is Associated With Increased Hepatic Retinyl-Palmitate Storage in Humans. J. Clin. Endocrinol. Metab. 2015, 100, E1568–E1574. [Google Scholar] [CrossRef] [PubMed]
- Looker, A.C.; Johnson, C.L.; Underwood, B.A. Serum retinol levels of persons aged 4–74 years from three Hispanic groups. Am. J. Clin. Nutr. 1988, 48, 1490–1496. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention Second Nutrition Report. 2012. Available online: https://www.cdc.gov/nutritionreport/report_2012.html (accessed on 13 July 2020).
- Gueguen, S.; Leroy, P.; Gueguen, R.; Siest, G.; Visvikis, S.; Herbeth, B. Genetic and environmental contributions to serum retinol and alpha-tocopherol concentrations: The Stanislas Family Study. Am. J. Clin. Nutr. 2005, 81, 1034–1044. [Google Scholar] [CrossRef]
- Blomhoff, R.; Green, M.; Berg, T.; Norum, K. Transport and storage of vitamin A. Science 1990, 250, 399–404. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E. Omega-3 Fatty Acids and FFAR. Front. Endocrinol. 2014, 5, 115. [Google Scholar] [CrossRef]
- Quadro, L.; Hamberger, L.; Colantuoni, V.; Gottesman, M.E.; Blaner, W.S. Understanding the physiological role of retinol-binding protein in vitamin A metabolism using transgenic and knockout mouse models. Mol. Asp. Med. 2003, 24, 421–430. [Google Scholar] [CrossRef]
- Baeten, J.M.; Richardson, B.A.; Bankson, D.D.; Wener, M.H.; Kreiss, J.K.; Lavreys, L.; Mandaliya, K.; Bwayo, J.J.; McClelland, R.S. Use of serum retinol-binding protein for prediction of vitamin A deficiency: Effects of HIV-1 infection, protein malnutrition, and the acute phase response. Am. J. Clin. Nutr. 2004, 79, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Blaner, W.S. Retinol-binding protein: The serum transport protein for vitamin A. Endocr. Rev. 1989, 10, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Soprano, D.R.; Soprano, K.J.; Goodman, D.S. Retinol-binding protein messenger RNA levels in the liver and in extrahepatic tissues of the rat. J. Lipid Res. 1986, 27, 166–171. [Google Scholar] [CrossRef]
- Tian, C.; Stokowski, R.P.; Kershenobich, D.; Ballinger, D.G.; Hinds, D.A. Variant in PNPLA3 is associated with alcoholic liver disease. Nat. Genet. 2010, 42, 21–23. [Google Scholar] [CrossRef]
- He, S.; McPhaul, C.; Li, J.Z.; Garuti, R.; Kinch, L.; Grishin, N.V.; Cohen, J.C.; Hobbs, H.H. A sequence variation (I148M) in PNPLA3 associated with nonalcoholic fatty liver disease disrupts triglyceride hydrolysis. J. Biol. Chem. 2010, 285, 6706–6715. [Google Scholar] [CrossRef]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2008, 40, 1461–1465. [Google Scholar] [CrossRef]
- Larrieta-Carrasco, E.; Acuña-Alonzo, V.; Velázquez-Cruz, R.; Barquera-Lozano, R.; León-Mimila, P.; Villamil-Ramírez, H.; Menjivar, M.; Romero-Hidalgo, S.; Méndez-Sánchez, N.; Cárdenas, V.; et al. PNPLA3 I148M polymorphism is associated with elevated alanine transaminase levels in Mexican Indigenous and Mestizo populations. Mol. Biol. Rep. 2014, 41, 4705–4711. [Google Scholar] [CrossRef]
- Chen, F.; Marquez, H.; Kim, Y.-K.; Qian, J.; Shao, F.; Fine, A.; Cruikshank, W.W.; Quadro, L.; Cardoso, W.V. Prenatal retinoid deficiency leads to airway hyperresponsiveness in adult mice. J. Clin. Investig. 2014, 124, 801–811. [Google Scholar] [CrossRef]
- Desai, T.J.; Chen, F.; Lü, J.; Qian, J.; Niederreither, K.; Dollé, P.; Chambon, P.; Cardoso, W.V. Distinct roles for retinoic acid receptors alpha and beta in early lung morphogenesis. Dev. Biol. 2006, 291, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Wirth, J.P.; Petry, N.; Tanumihardjo, S.A.; Rogers, L.M.; McLean, E.; Greig, A.; Garrett, G.S.; Klemm, R.D.W.; Rohner, F. Vitamin A Supplementation Programs and Country-Level Evidence of Vitamin A Deficiency. Nutrients 2017, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Vitamin A Supplementation during Pregnancy. Available online: https://www.who.int/elena/titles/guidance_summaries/vitamina_pregnancy/en/ (accessed on 13 July 2020).
- Radhika, M.S.; Bhaskaram, P.; Balakrishna, N.; Ramalakshmi, B.A.; Devi, S.; Kumar, B.S. Effects of vitamin A deficiency during pregnancy on maternal and child health. BJOG 2002, 109, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Bhat, P.V.; Manolescu, D.-C. Role of vitamin A in determining nephron mass and possible relationship to hypertension. J. Nutr. 2008, 138, 1407–1410. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Participants (n) | VAD * (n = 40) | VAS † (n = 27) | p Trend | |
|---|---|---|---|---|
| Ethnicity, n (%) | Hispanic/Latin American | 29 (72.5) | 15 (55.6) | 0.311 |
| non-Hispanic Black | 8 (20.0) | 7 (25.9) | ||
| Other | 3 (7.5) | 5 (18.5) | ||
| Age (y) | 29.1 (5.4) | 31.3 (6.1) | 0.138 | |
| Multipara, n (%) | No | 10 (25.0) | 7 (26.9) | 1.000 |
| Yes | 30 (75.0) | 19 (73.1) | ||
| Bariatric surgery, n(%) | No | 21 (52.5) | 13 (48.1) | 0.806 |
| Yes | 19 (47.5) | 14 (51.9) | ||
| Education level, n (%) | 0.458 | |||
| <High school | 5 (13.2) | 4 (16.7) | ||
| High School | 22 (57.9) | 10 (41.7) | ||
| College | 11 (28.9) | 10 (41.7) | ||
| Body Mass Index (kg/m2) | Prepregnancy | 31.8 (7.6) | 32.1 (7.4) | 0.878 |
| At delivery | 36.0 (7.1) | 36.9 (6.3) | 0.619 | |
| Obese (prepregnancy), n (%) | 0.952 | |||
| Normal Weight | 5 (12.8) | 3 (11.5) | ||
| Over Weight | 9 (23.1) | 8 (30.8) | ||
| Class I obesity | 13 (33.3) | 8 (30.8) | ||
| Class II obesity | 12 (30.8) | 7 (26.9) | ||
| Gestational weight gain (GWG) | 24.4 (20.1) | 26.2 (12.3) | 0.683 | |
| GWG recommended level, n (%) | Below | 5 (13.2) | 2 (7.7) | 0.691 |
| Above | 33 (86.8) | 24 (92.3) | ||
| Delivery mode (C-section), n (%) | No | 24 (61.5) | 12 (46.2) | 0.309 |
| Yes | 15 (38.5) | 14 (53.8) | ||
| Baby weight (g) | 3053.8 (602.9) | 3104.4 (601.1) | 0.742 | |
| Large for genstational age, n (%) | No | 32 (97.0) | 22 (95.7) | 1.000 |
| Yes | 1 (3.0) | 1 (4.3) | ||
| Serum retinol (µmol/L) | Postpartum (mother) | 0.5 (0.4) | 0.5 (0.6) | 0.886 |
| Cord blood | 0.2 (0.1) | 0.3 (0.1) | 0.041 | |
| Serum beta carotene (ng/100 µL) | Third trimester (mother) | 3.3 (2.0) | 6.2 (5.9) | 0.007 |
| Postpertum (mother) | 2.6 (2.1) | 3.1 (1.7) | 0.413 | |
| Cord blood | 1.7 (0.6) | 2.0 (0.6) | 0.554 | |
| Non-Hispanic White | Non-Hispanic Black | Mexican American | Other Hispanic | Other | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total * | Pregnant | Pregnant with PIR <1.85 | Total * | Pregnant | Pregnant with PIR <1.85 | Total * | Pregnant | Pregnant with PIR <1.85 | Total * | Pregnant | Pregnant with PIR <1.85 | Total * | Pregnant | Pregnant with PIR <1.85 | ||
| Total N (%) | 1684 (41.3) | 392 (45.5) | 115 (26.9) | 976 (23.9) | 145 (16.8) | 91 (21.3) | 1071 (26.2) | 239 (27.8) | 181 (42.3) | 168 (4.1) | 36 (4.2) | 22 (5.1) | 183 (4.5) | 49 (5.7) | 19 (4.4) | |
| Vitamin A status | Deficient | 60 (3.6) | 43 (11.0) | 14 (12.2) | 128 (13.1) | 47 (32.4) | 33 (36.3) | 72 (6.7) | 36 (15.1) | 29 (16.0) | 15 (8.9) | 10 (27.8) | 6 (27.3) | 14 (7.7) | 6 (12.2) | 2 (10.5) |
| Sufficient | 1624 (96.4) | 349 (89.0) | 101 (87.8) | 848 (86.9) | 98 (67.6) | 58 (63.7) | 999 (93.3) | 203 (84.9) | 152 (84.0) | 153 (91.1) | 26 (72.2) | 16 (72.7) | 169 (92.3) | 43 (87.8) | 17 (89.5) | |
| Age (y) | Mean (SD) | 28.3 (7.6) | 27.4 (5.3) | 24.5 (5.3) | 25.6 (8.0) | 24.3 (5.6) | 23.4 (4.8) | 26.2 (7.9) | 25.5 (5.5) | 25.5 (5.5) | 27.2 (7.7) | 26.9 (6.1) | 25.7 (6.3) | 28.2 (7.8) | 28.2 (4.8) | 25.4 (4.8) |
| Poverty Income Rate | Mean (SD) | 2.9 (1.6) | 3.1 (1.6) | 1.0 (0.5) | 1.8 (1.5) | 1.7 (1.4) | 0.7 (0.5) | 1.7 (1.3) | 1.5 (1.2) | 0.9 (0.5) | 1.9 (1.4) | 1.9 (1.6) | 0.9 (0.5) | 2.7 (1.7) | 3.1 (1.9) | 1.0 (0.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, M.; Wang, T.; Garretto, D.; Isasi, C.R.; Cardoso, W.V.; Greally, J.M.; Quadro, L. Disproportionate Vitamin A Deficiency in Women of Specific Ethnicities Linked to Differences in Allele Frequencies of Vitamin A-Related Polymorphisms. Nutrients 2021, 13, 1743. https://doi.org/10.3390/nu13061743
Suzuki M, Wang T, Garretto D, Isasi CR, Cardoso WV, Greally JM, Quadro L. Disproportionate Vitamin A Deficiency in Women of Specific Ethnicities Linked to Differences in Allele Frequencies of Vitamin A-Related Polymorphisms. Nutrients. 2021; 13(6):1743. https://doi.org/10.3390/nu13061743
Chicago/Turabian StyleSuzuki, Masako, Tao Wang, Diana Garretto, Carmen R. Isasi, Wellington V. Cardoso, John M. Greally, and Loredana Quadro. 2021. "Disproportionate Vitamin A Deficiency in Women of Specific Ethnicities Linked to Differences in Allele Frequencies of Vitamin A-Related Polymorphisms" Nutrients 13, no. 6: 1743. https://doi.org/10.3390/nu13061743
APA StyleSuzuki, M., Wang, T., Garretto, D., Isasi, C. R., Cardoso, W. V., Greally, J. M., & Quadro, L. (2021). Disproportionate Vitamin A Deficiency in Women of Specific Ethnicities Linked to Differences in Allele Frequencies of Vitamin A-Related Polymorphisms. Nutrients, 13(6), 1743. https://doi.org/10.3390/nu13061743