Effects of Different n6/n3 PUFAs Dietary Ratio on Cardiac Diabetic Neuropathy

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics
2.2. Experimental Design
2.3. Sample Collection
2.4. Immunohistochemistry
2.5. Data Acquisition and Analysis
2.6. Statistical Analyses
3. Results
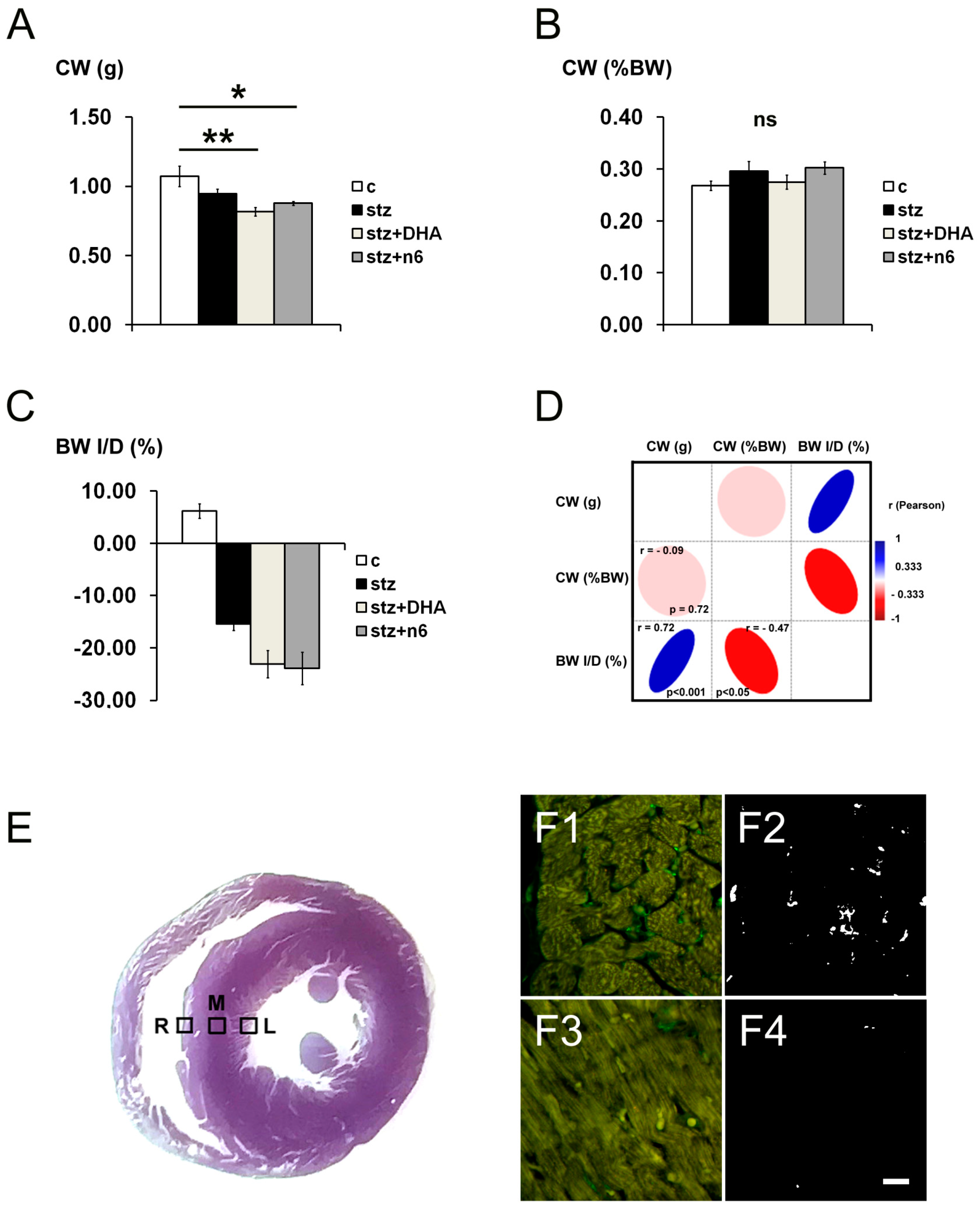
3.1. General Remarks
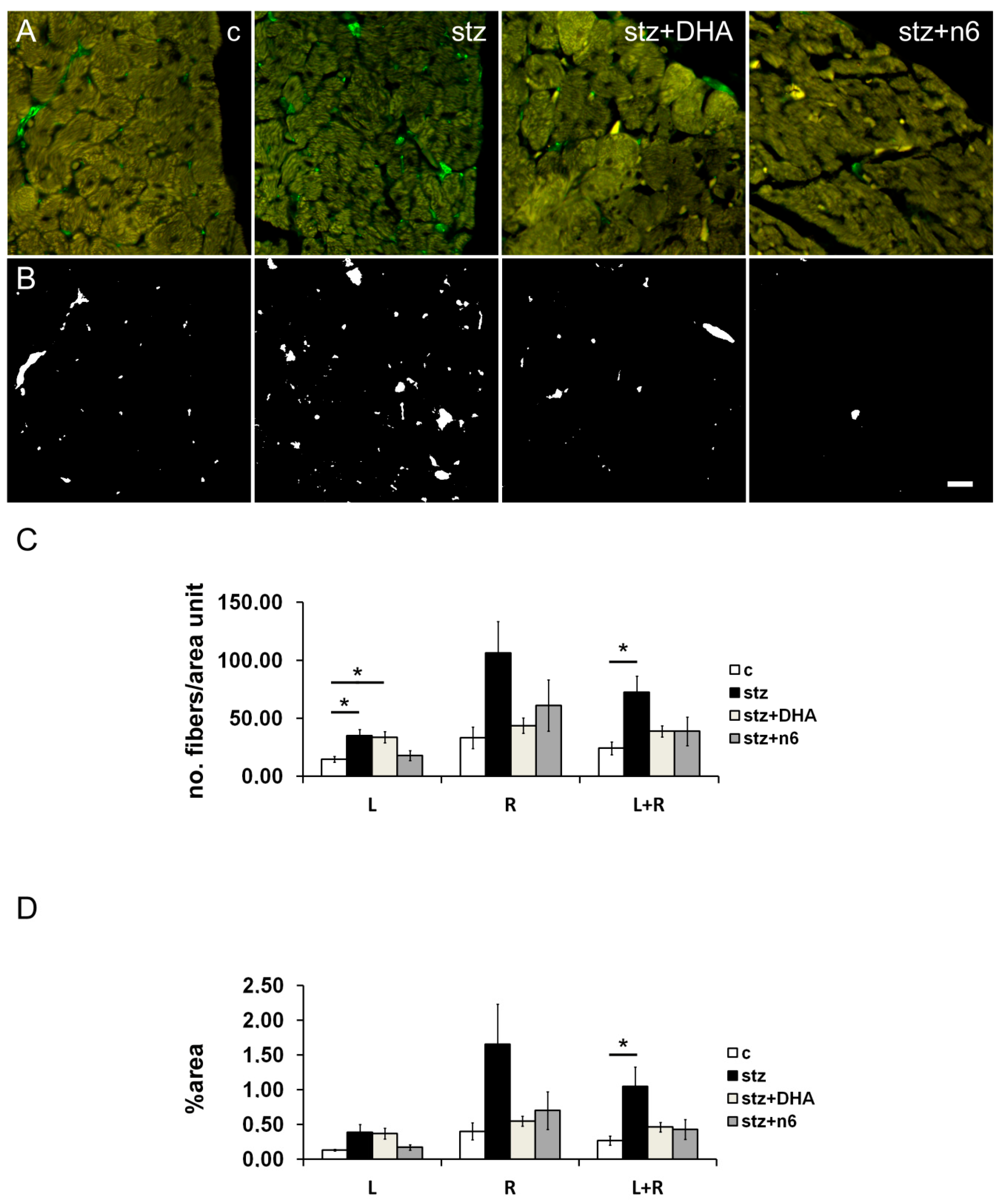
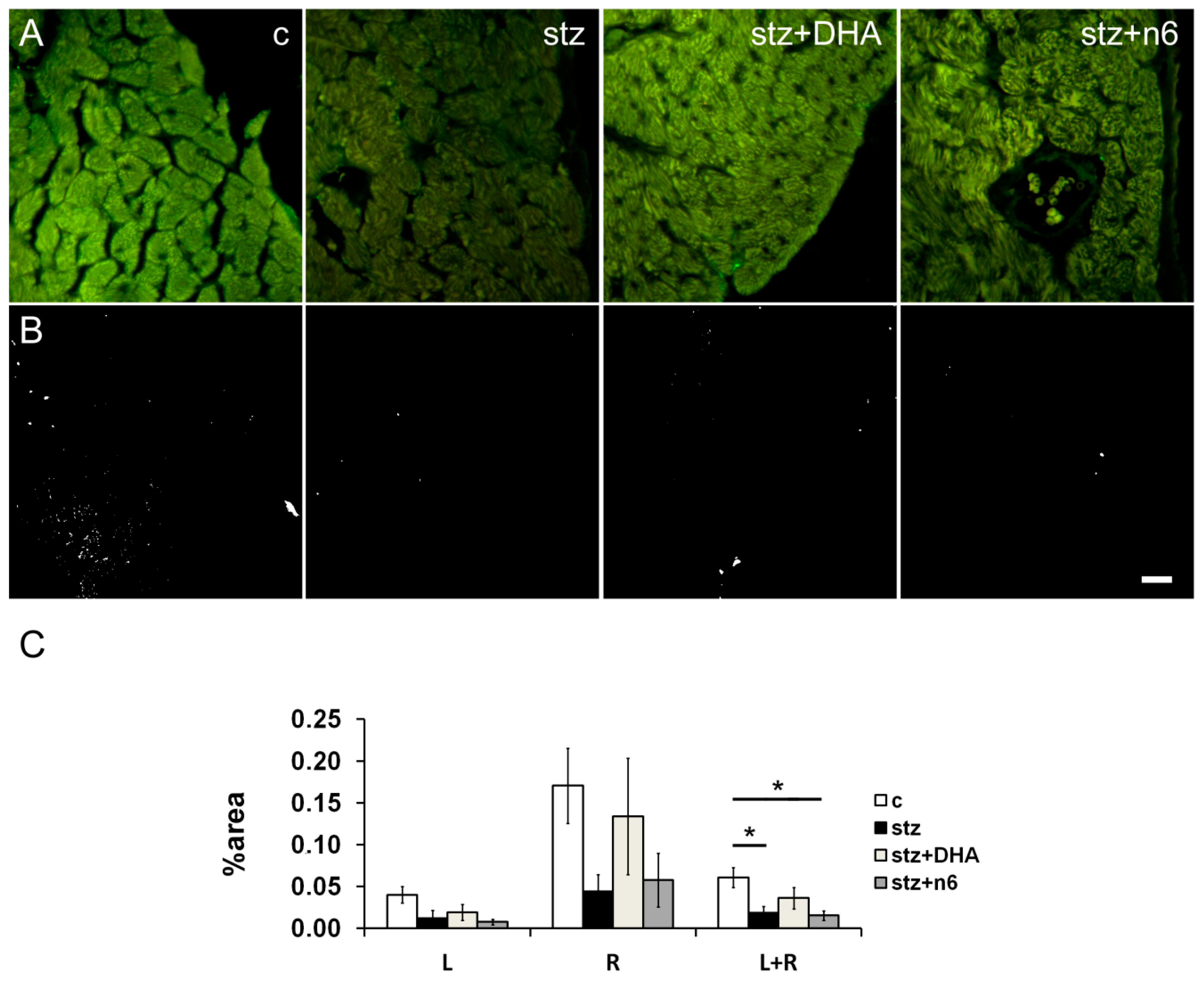
3.2. Protein g Product 9.5—Immunoreactive Fibers Density
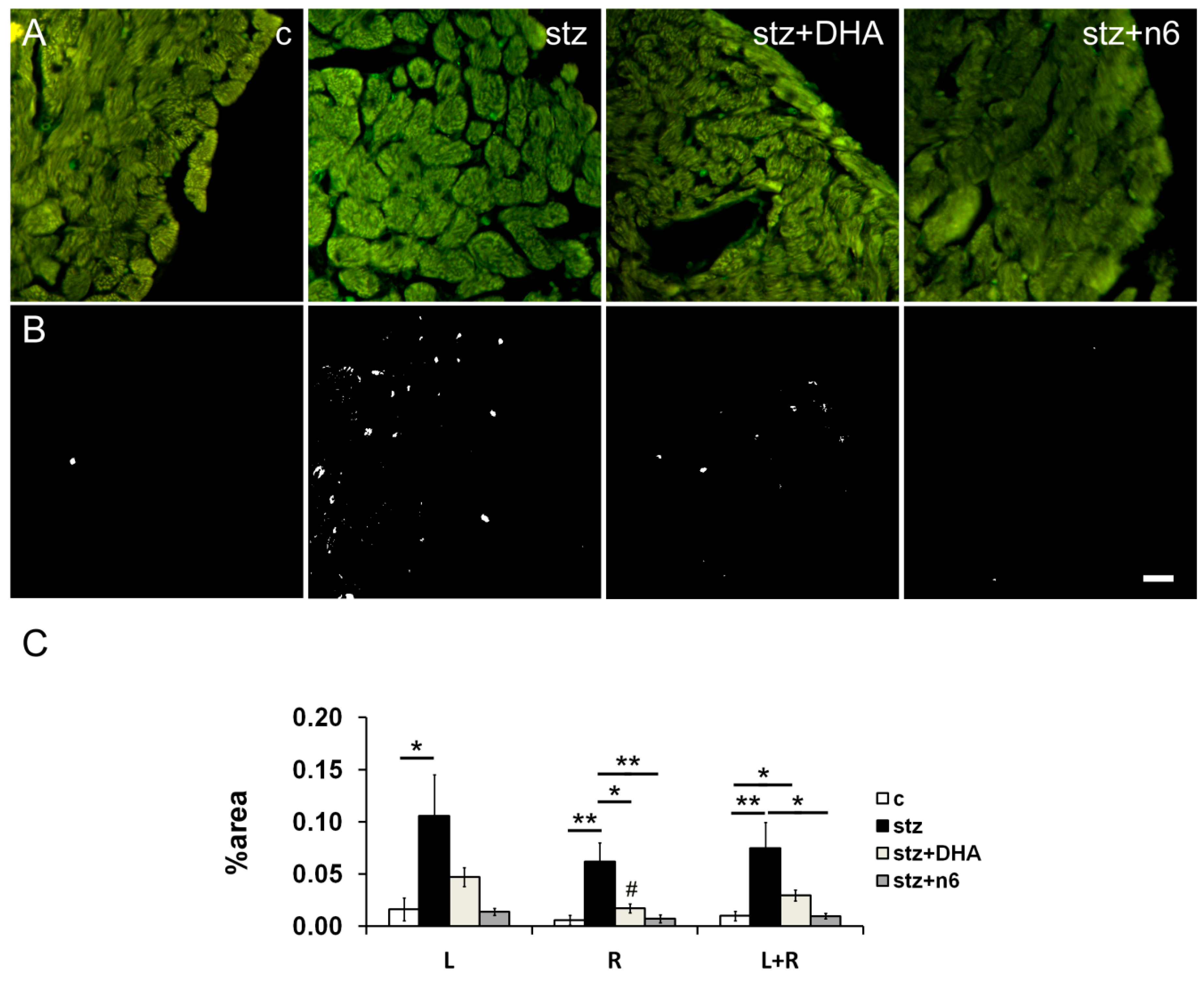
3.3. Neurofilament 200—Immunoreactive Fibers Density
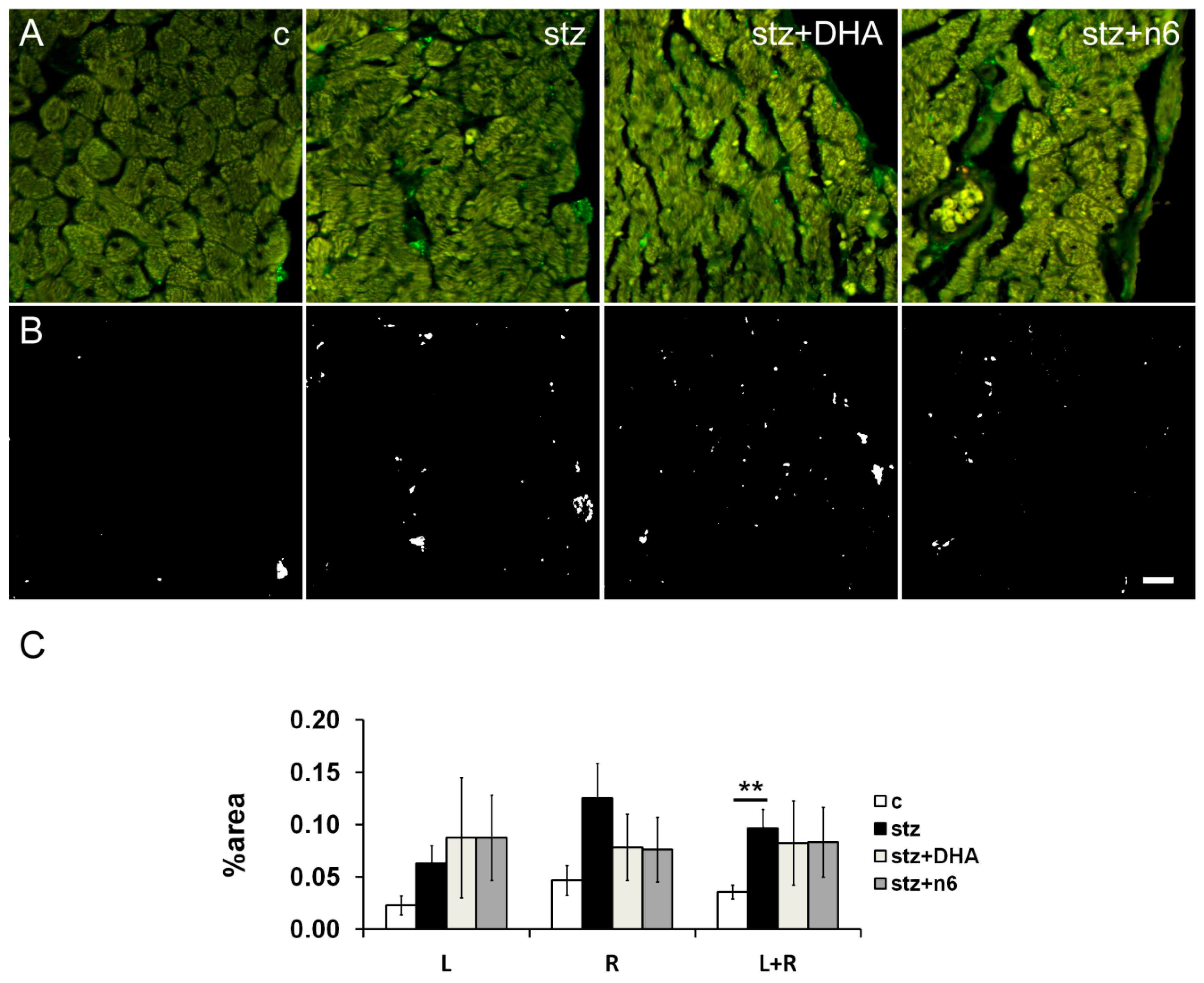
3.4. Calcitonin Gene-Related Peptide—Immunoreactive Fibers Density
3.5. Tyrosine Hydroxylase—Immunoreactive Fibers Density
3.6. Choline Acetyltransferase—Immunoreactive Fibers Density
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Verhulst, M.J.L.; Loos, B.G.; Gerdes, V.E.A.; Teeuw, W.J. Evaluating All Potential Oral Complications of Diabetes Mellitus. Front. Endocrinol. 2019, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.C.; Kuncova, J.; Pfeil, U.; McGregor, G.; Sviglerova, J.; Slavikova, J.; Kummer, W. Cardiomyopathy in streptozotocin-induced diabetes involves intra-axonal accumulation of calcitonin gene-related peptide and altered expression of its receptor in rats. Neuroscience 2005, 134, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Vinik, A.I.; Erbas, T.; Casellini, C.M. Diabetic cardiac autonomic neuropathy, inflammation and cardiovascular disease. J. Diabetes Investig. 2013, 4, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Mašek, T.; Filipovic, N.; Hamzić, L.F.; Puljak, L.; Starčević, K. Long-term streptozotocin diabetes impairs arachidonic and docosahexaenoic acid metabolism and ∆5 desaturation indices in aged rats. Exp. Gerontol. 2014, 60, 140–146. [Google Scholar] [CrossRef]
- Tosi, F.; Sartori, F.; Guarini, P.; Olivieri, O.; Martinelli, N. Delta-5 and Delta-6 Desaturases: Crucial Enzymes in Polyunsaturated Fatty Acid-Related Pathways with Pleiotropic Influences in Health and Disease. Adv. Exp. Med. Biol. 2014, 824, 61–81. [Google Scholar] [CrossRef]
- Takahashi, M.; Ando, J.; Shimada, K.; Nishizaki, Y.; Tani, S.; Ogawa, T.; Yamamoto, M.; Nagao, K.; Hirayama, A.; Yoshimura, M.; et al. The ratio of serum n-3 to n-6 polyunsaturated fatty acids is associated with diabetes mellitus in patients with prior myocardial infarction: A multicenter cross-sectional study. BMC Cardiovasc. Disord. 2017, 17, 41. [Google Scholar] [CrossRef]
- Bang, H.O.; Dyerberg, J.; Hjoorne, N. The composition of food consumed by Greenland Eskimos. Acta Med. Scand. 1976, 200, 69–73. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids in the prevention-management of cardiovascular disease. Can. J. Physiol. Pharmacol. 1997, 75, 234–239. [Google Scholar]
- Horrocksab, L.A.; Yeo, Y.K. Health Benefits of Docosahexaenoic Acid (DHA). Pharmacol. Res. 1999, 40, 211–225. [Google Scholar] [CrossRef]
- Yokoyama, M.; Origasa, H.; Matsuzaki, M.; Matsuzawa, Y.; Saito, Y.; Ishikawa, Y.; Oikawa, S.; Sasaki, J.; Hishida, H.; Itakura, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (JELIS): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar] [CrossRef]
- Shoji, T.; Kakiya, R.; Hayashi, T.; Tsujimoto, Y.; Sonoda, M.; Shima, H.; Mori, K.; Fukumoto, S.; Tahara, H.; Shioi, A.; et al. Serum n−3 and n−6 Polyunsaturated Fatty Acid Profile as an Independent Predictor of Cardiovascular Events in Hemodialysis Patients. Am. J. Kidney Dis. 2013, 62, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Harbige, L.S. Fatty acids, the immune response, and autoimmunity: A question of n−6 essentiality and the balance between n−6 and n−3. Lipids 2003, 38, 323–341. [Google Scholar] [CrossRef]
- Billman, G.E. The effects of omega-3 polyunsaturated fatty acids on cardiac rhythm: A critical reassessment. Pharmacol. Ther. 2013, 140, 53–80. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.F.; Thivierge, M.C.; Kidd, C.A.; McGeoch, S.C.; Abraham, P.; Pearson, D.W.M.; Horgan, G.W.; Holtrop, G.; Thiès, F.; Lobley, G.E. Fish oil supplemented for 9 months does not improve glycaemic control or insulin sensitivity in subjects with impaired glucose regulation: A parallel randomised controlled trial. Br. J. Nutr. 2015, 115, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.J.; Aung, T.; Armitage, J. Questioning the Associations of omega-3 Fatty Acid Supplement Use With Cardiovascular Disease Risks—Reply. JAMA Cardiol. 2018, 3, 781–782. [Google Scholar] [CrossRef] [PubMed]
- Vikøren, L.A.; Drotningsvik, A.; Bergseth, M.T.; Mjøs, S.A.; Austgulen, M.H.; Mellgren, G.; Gudbrandsen, O.A. Intake of Baked Cod Fillet Resulted in Lower Serum Cholesterol and Higher Long Chain n-3 PUFA Concentrations in Serum and Tissues in Hypercholesterolemic Obese Zucker fa/fa Rats. Nutrients 2018, 10, 840. [Google Scholar] [CrossRef]
- Aloud, B.M.; Raj, P.; O’Hara, K.; Shao, Z.; Yu, L.; Anderson, H.D.; Netticadan, T. Conjugated linoleic acid prevents high glucose–induced hypertrophy and contractile dysfunction in adult rat cardiomyocytes. Nutr. Res. 2016, 36, 134–142. [Google Scholar] [CrossRef]
- Anna, Z.; Angela, S.; Barbara, B.; Jana, R.; Tamara, B.; Csilla, V.; Victor, D.; Oleksiy, M.; Tribulova, N. Heart-protective effect of n-3 PUFA demonstrated in a rat model of diabetic cardiomyopathy. Mol. Cell. Biochem. 2013, 389, 219–227. [Google Scholar] [CrossRef]
- Bhandarkar, N.; Kumar, S.; Martin, J.M.; Brown, L.; Panchal, S.K. Attenuation of Metabolic Syndrome by EPA/DHA Ethyl Esters in Testosterone-Deficient Obese Rats. Mar. Drugs 2018, 16, 182. [Google Scholar] [CrossRef]
- Christensen, J.H.; Schmidt, E.B. Autonomic nervous system, heart rate variability and n-3 fatty acids. J. Cardiovasc. Med. 2007, 8, S19–S22. [Google Scholar] [CrossRef]
- Bowman, L.; Mafham, M.; Wallendszus, K.; Stevens, W.; Buck, G.; Barton, J.; Murphy, K.; Aung, T.; Haynes, R.; Cox, J.; et al. Effects of n-3 Fatty Acid Supplements in Diabetes Mellitus. N. Engl. J. Med. 2018, 379, 1540–1550. [Google Scholar]
- Billman, G.E.; Harris, W.S. Effect of dietary omega-3 fatty acids on the heart rate and the heart rate variability responses to myocardial ischemia or submaximal exercise. Am. J. Physiol. Circ. Physiol. 2011, 300, H2288–H2299. [Google Scholar] [CrossRef]
- Lundbye-Christensen, S. Omega-3 Polyunsaturated Fatty Acids and Heart Rate Variability. Front. Physiol. 2011, 2, 84. [Google Scholar] [CrossRef]
- Coste, T.C.; Gerbi, A.; Vague, P.; Pieroni, G.; Raccah, D. Neuroprotective effect of docosahexaenoic acid-enriched phospholipids in experimental diabetic neuropathy. Diabetes 2003, 52, 2578–2585. [Google Scholar] [CrossRef]
- Pham, T.; He, J.; Kakazu, A.H.; Jun, B.; Bazan, N.G.; Bazan, H.E.P. Defining a mechanistic link between pigment epithelium–derived factor, docosahexaenoic acid, and corneal nerve regeneration. J. Boil. Chem. 2017, 292, 18486–18499. [Google Scholar] [CrossRef]
- He, J.; Pham, T.; Kakazu, A.; Bazan, H.E. Recovery of Corneal Sensitivity and Increase in Nerve Density and Wound Healing in Diabetic Mice After PEDF Plus DHA Treatment. Diabetes 2017, 66, 2511–2520. [Google Scholar] [CrossRef]
- Starcevic, K.; Filipovic, N.; Galan, A.; Micek, V.; Gudan-Kurilj, A.; Masek, T. Hepatic Lipogenesis and Brain Fatty Acid Profile in Response to Different Dietary n6/n3 Ratios and DHA/EPA Supplementation in Streptozotocin Treated Rats. Mol. Nutr. Food Res. 2018, 62, e1701007. [Google Scholar] [CrossRef]
- Vitlov-Uljević, M.; Starčević, K.; Mašek, T.; Bočina, I.; Restović, I.; Kević, N.; Racetin, A.; Kretzschmar, G.; Grobe, M.; Vukojević, K.; et al. Dietary DHA/EPA supplementation ameliorates diabetic nephropathy by protecting from distal tubular cell damage. Cell Tissue Res. 2019, 378, 301–317. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Mead, R.; Gilmour, S.G.; Mead, A. Statistical Principles for the Design of Experiments: Applications to Real Experiments; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Bakovic, M.; Filipovic, N.; Hamzic, L.F.; Kunac, N.; Zdrilic, E.; Uljevic, M.V.; Kostić, S.; Puljak, L.; Vukojević, K. Changes in neurofilament 200 and tyrosine hydroxylase expression in the cardiac innervation of diabetic rats during aging. Cardiovasc. Pathol. 2018, 32, 38–43. [Google Scholar] [CrossRef]
- Bakovic, M.; Paic, M.J.; Zdrilic, E.; Vukojević, K.; Ferhatovic, L.; Marin, A.; Filipovic, N.; Grkovic, I.; Puljak, L. Changes in cardiac innervation during maturation in long-term diabetes. Exp. Gerontol. 2013, 48, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- van Acker, N.; Ragé, M.; Sluydts, E.; Knaapen, M.W.M.; de Bie, M.; Timmers, M.; Fransen, E.; Duymelinck, C.; de Schepper, S.; Anand, P.; et al. Automated PGP9.5 immunofluorescence staining: A valuable tool in the assessment of small fiber neuropathy? BMC Res. Notes 2016, 9, 280. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.J.; Grkovic, I.; Anderson, C.R. Immunohistochemical analysis of intracardiac ganglia of the rat heart. Cell Tissue Res. 2003, 314, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Vo, P.A.; Tomlinson, D.R. Effects of nerve growth factor on expression of GAP-43 in right atria after sympathectomy in diabetic rats. Diabetes Obes. Metab. 2001, 3, 350–359. [Google Scholar] [CrossRef]
- Porseva, V.V.; Smirnova, V.P.; Korzina, M.B.; Emanuilov, A.I.; Masliukov, P.M. Age-associated changes in sympathetic neurons containing neurofilament 200 kDa during chemical deafferentation. Bull. Exp. Boil. Med. 2013, 155, 268–271. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.V.; Nixon, R.A. Neurofilaments and Neurofilament Proteins in Health and Disease. Cold Spring Harb. Perspect. Boil. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Guidato, S.; Bajaj, N.P.; Miller, C.C. Cellular phosphorylation of neurofilament heavy-chain by cyclin-dependent kinase-5 masks the epitope for monoclonal antibody N52. Neurosci. Lett. 217, 157–160. [CrossRef]
- Guidato, S.; Tsai, L.H.; Woodgett, J.; Miller, C.C. Differential cellular phosphorylation of neurofilament heavy side-arms by glycogen synthase kinase-3 and cyclin-dependent kinase-5. J. Neurochem. 66, 1698–1706. [CrossRef]
- Restovic, I.; Bočina, I.; Vukojević, K.; Kero, D.; Filipovic, N.; Raonić, J.; Vučinić, J.; Vukmirović, F.; Vučković, L.; Saraga-Babic, M. Time course and expression pattern of the neuronal markers in the developing human spinal cord. Int. J. Dev. Neurosci. 2019, 74, 1–10. [Google Scholar] [CrossRef]
- Yoshimura, N.; Erdman, S.L.; Snider, M.W.; de Groat, W.C. Effects of spinal cord injury on neurofilament immunoreactivity and capsaicin sensitivity in rat dorsal root ganglion neurons innervating the urinary bladder. Neuroscience 1998, 83, 633–643. [Google Scholar] [CrossRef]
- Guić, M.M.; Košta, V.; Aljinović, J.; Sapunar, D.; Grković, I. Characterization of spinal afferent neurons projecting to different chambers of the rat heart. Neurosci. Lett. 2010, 469, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Košta, V.; Guić, M.M.; Aljinović, J.; Sapunar, D.; Grković, I. Immunohistochemical characteristics of neurons in nodose ganglia projecting to the different chambers of the rat heart. Auton. Neurosci. 2010, 155, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.; King, R.; Smillie, S.-J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, Y.-H.; Jiang, P.; Lin, H.-Q.; Yang, J.-L.; Ma, D.-F.; Wang, X.; Yang, C.-H. Analysis of Heart Rate Variability and Cardiac Autonomic Nerve Remodeling in Streptozotocin-induced Diabetic Rats. Exp. Clin. Endocrinol. Diabetes 2015, 123, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, J.-L.; Ma, D.-F.; Lin, H.-Q.; Xu, X.-D.; Yue, H.J. Effects of Different Compatibilities of Ramulus Cinnamomi and Peony in Guizhi Decoction on Diabetic Cardiac Autonomic Neuropathy. Zhongguo Zhong xi yi jie he za zhi Zhongguo Zhongxiyi jiehe zazhi Chin. J. Integr. Tradit. West. Med. 2015, 35, 741–745. [Google Scholar]
- Nandi, S.S.; Zheng, H.; Sharma, N.M.; Shahshahan, H.R.; Patel, K.P.; Mishra, P.K. Lack of miR-133a Decreases Contractility of Diabetic Hearts: A Role for Novel Cross Talk Between Tyrosine Aminotransferase and Tyrosine Hydroxylase. Diabetes 2016, 65, 3075–3090. [Google Scholar] [CrossRef]
- Xuan, Y.-L.; Wang, Y.; Xue, M.; Hu, H.-S.; Cheng, W.; Li, X.; Yin, J.; Yang, N.; Yan, S. In rats the duration of diabetes influences its impact on cardiac autonomic innervations and electrophysiology. Auton. Neurosci. 2015, 189, 31–36. [Google Scholar] [CrossRef]
- Oda, Y. Choline acetyltransferase: The structure, distribution and pathologic changes in the central nervous system. Pathol. Int. 1999, 49, 921–937. [Google Scholar] [CrossRef]
- Chessa, M.; Butera, G.; Lanza, G.A.; Bossone, E.; Delogu, A.; de Rosa, G.; Marietti, G.; Rosti, L.; Carminati, M. Role of Heart Rate Variability in the Early Diagnosis of Diabetic Autonomic Neuropathy in Children. Herz 2002, 27, 785–790. [Google Scholar] [CrossRef]
- Thayer, J.F.; Lane, R.D. The role of vagal function in the risk for cardiovascular disease and mortality. Boil. Psychol. 2007, 74, 224–242. [Google Scholar] [CrossRef]
- Ziegler, D.; Gries, F.A.; Muhlen, H.; Rathmann, W.; Spuler, M.; Lessmann, F. Prevalence and clinical correlates of cardiovascular autonomic and peripheral diabetic neuropathy in patients attending diabetes centers. The Diacan Multicenter Study Group. Diabete Metab. 1993, 19, 143–151. [Google Scholar] [PubMed]
- Lund, D.D.; Subieta, A.R.; Pardini, B.J.; Chang, K.S. Alterations in cardiac parasympathetic indices in STZ-induced diabetic rats. Diabetes 1992, 41, 160–166. [Google Scholar] [CrossRef]
- Coppey, L.; Davidson, E.; Shevalye, H.; Obrosov, A.; Yorek, M. Effect of Early and Late Interventions with Dietary Oils on Vascular and Neural Complications in a Type 2 Diabetic Rat Model. J. Diabetes Res. 2019, 2019, 5020465. [Google Scholar] [CrossRef] [PubMed]
- Yorek, M.A. The Potential Role of Fatty Acids in Treating Diabetic Neuropathy. Curr. Diabetes Rep. 2018, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X. Reduction of heart rate by omega-3 fatty acids and the potential underlying mechanisms. Front. Physiol. 2012, 3, 416. [Google Scholar] [CrossRef]
- Scarpini, E.; Bianchi, R.; Moggio, M.; Sciacco, M.; Fiori, M.G.; Scarlato, G. Decrease of nerve Na+, K(+)-ATPase activity in the pathogenesis of human diabetic neuropathy. J. Neurol. Sci. 1993, 120, 159–167. [Google Scholar] [CrossRef]
- Lorentz, C.U.; Woodward, W.R.; Tharp, K.; Habecker, B.A. Altered norepinephrine content and ventricular function in p75NTR-/- mice after myocardial infarction. Auton. Neurosci. 2011, 164, 13–19. [Google Scholar] [CrossRef]
- Huang, J.P.; Cheng, M.L.; Hung, C.Y.; Wang, C.H.; Hsieh, P.S.; Shiao, M.S.; Chen, J.K.; Li, D.E.; Hung, L.M. Docosapentaenoic acid and docosahexaenoic acid are positively associated with insulin sensitivity in rats fed high-fat and high-fructose diets. J. Diabetes 2017, 9, 936–946. [Google Scholar] [CrossRef]
- Ma, C.; Xu, Z.; Lv, H. Low n-6/n-3 PUFA ratio improves inflammation and myocardial ischemic reperfusion injury. Biochem. Cell. Biol. 2019, 97, 621–629. [Google Scholar] [CrossRef]
- Zarei, M.; Fakher, S.; Tabei, S.M.B.; Javanbakht, M.H.; Derakhshanian, H.; Farahbakhsh-Farsi, P.; Sadeghi, M.R.; Mostafavi, E.; Djalali, M. Effects of vitamin A, C and E, or omega-3 fatty acid supplementation on the level of paraoxonase and arylesterase activity in streptozotocin-induced diabetic rats: An investigation of activities in plasma, and heart and liver homogenates. Singap. Med. J. 2016, 57, 153–156. [Google Scholar] [CrossRef]
- Coppey, L.J.; Davidson, E.P.; Obrosov, A.; Yorek, M.A. Enriching the diet with menhaden oil improves peripheral neuropathy in streptozotocin-induced type 1 diabetic rats. J. Neurophysiol. 2015, 113, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Coppey, L.J.; Holmes, A.; Davidson, E.P.; Yorek, M.A. Partial Replacement with Menhaden Oil Improves Peripheral Neuropathy in High-Fat-Fed Low-Dose Streptozotocin Type 2 Diabetic Rat. J. Nutr. Metab. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y. Effects of a fatty acid deficiency on lipids of whole brain, microsomes, and myelin in the rat. J. Lipid Res. 1972, 13, 56–62. [Google Scholar] [PubMed]
- Barcelo-Coblijn, G.; Murphy, E.J.; Mills, K.; Winchester, B.; Jakobs, C.; Snead Iii, O.C.; Gibson, K.M. Lipid abnormalities in succinate semialdehyde dehydrogenase (Aldh5a1-/-) deficient mouse brain provide additional evidence for myelin alterations. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 556–562. [Google Scholar] [CrossRef][Green Version]
- Horrobin, D.F. The roles of essential fatty acids in the development of diabetic neuropathy and other complications of diabetes mellitus. Prostaglandins Leukot. Essent. Fat. Acids 1988, 31, 181–197. [Google Scholar]
- Ruiz-Gutierrez, V.; Stiefel, P.; Villar, J.; Acosta, D.; Carneado, J. Cell membrane fatty acid composition in Type 1 (insulin-dependent) diabetic patients: Relationship with sodium transport abnormalities and metabolic control. Diabetologia 1993, 36, 850–856. [Google Scholar] [CrossRef]
- Huang, Y.S.; Horrobin, D.F.; Manku, M.S.; Mitchell, J.; Ryan, M.A. Tissue phospholipid fatty acid composition in the diabetic rat. Lipids 1984, 19, 367–370. [Google Scholar] [CrossRef]
- Holman, R.T.; Johnson, S.B.; Gerrard, J.M.; Mauer, S.M.; Kupcho-Sandberg, S.; Brown, D.M. Arachidonic acid deficiency in streptozotocin-induced diabetes. Proc. Natl. Acad. Sci. USA 1983, 80, 2375–2379. [Google Scholar] [CrossRef] [PubMed]
- Duvernoy, C.S.; Raffel, D.M.; Swanson, S.D.; Jaiswal, M.; Mueller, G.; Ibrahim, E.-S.; Pennathur, S.; Plunkett, C.; Stojanovska, J.; Brown, M.B.; et al. Left ventricular metabolism, function, and sympathetic innervation in men and women with type 1 diabetes. J. Nucl. Cardiol. 2016, 23, 960–969. [Google Scholar] [CrossRef]
- Stevens, M.J.; Dayanikli, F.; Raffel, D.M.; Allman, K.C.; Sandford, T.; Feldman, E.L.; Wieland, D.M.; Corbett, J.; Schwaiger, M. Scintigraphic Assessment of Regionalized Defects in Myocardial Sympathetic Innervation and Blood Flow Regulation in Diabetic Patients with Autonomic Neuropathy. J. Am. Coll. Cardiol. 1998, 31, 1575–1584. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary | Antibody | Code no. | Host | Dilution | Source |
| PGP9.5 Monoclonal Antibody (BH7) | 480012 | Mouse | 1:500 | Invitrogen | |
| Anti-Neurofilament 200 kDa Antibody, clone N52 | MAB5266 | Mouse | 1:100 | Sigma-Aldrich | |
| Anti-CGRP | ab36001 | Goat | 1:1000 | Abcam | |
| Anti-Tyrosine Hydroxylase antibody | ab113 | Sheep | 1:500 | Abcam | |
| Anti-Choline Acetyltransferase Antibody | AB144P | Goat | 1:100 | Millipore | |
| Secondary | Alexa Fluor@4 88AffiniPure D onkey Anti- Mouse lgG (H+L) | 715-545-150 | Donkey | 1:400 | Jackson Immuno Research Laboratories |
| Alexa Fluor@4 88 AffiniPure D onkey Anti- Goat lgG (H+L) | 705-545-003 | Donkey | 1:400 | Jackson Immuno Research Laboratories | |
| Donkey Anti-Sheep IgG H+L (Alexa Fluor® 488) | ab150177 | 1:400 | Abcam |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urlić, M.; Urlić, I.; Urlić, H.; Mašek, T.; Benzon, B.; Vitlov Uljević, M.; Vukojević, K.; Filipović, N. Effects of Different n6/n3 PUFAs Dietary Ratio on Cardiac Diabetic Neuropathy. Nutrients 2020, 12, 2761. https://doi.org/10.3390/nu12092761
Urlić M, Urlić I, Urlić H, Mašek T, Benzon B, Vitlov Uljević M, Vukojević K, Filipović N. Effects of Different n6/n3 PUFAs Dietary Ratio on Cardiac Diabetic Neuropathy. Nutrients. 2020; 12(9):2761. https://doi.org/10.3390/nu12092761
Chicago/Turabian StyleUrlić, Marjan, Ivanka Urlić, Hrvoje Urlić, Tomislav Mašek, Benjamin Benzon, Marija Vitlov Uljević, Katarina Vukojević, and Natalija Filipović. 2020. "Effects of Different n6/n3 PUFAs Dietary Ratio on Cardiac Diabetic Neuropathy" Nutrients 12, no. 9: 2761. https://doi.org/10.3390/nu12092761
APA StyleUrlić, M., Urlić, I., Urlić, H., Mašek, T., Benzon, B., Vitlov Uljević, M., Vukojević, K., & Filipović, N. (2020). Effects of Different n6/n3 PUFAs Dietary Ratio on Cardiac Diabetic Neuropathy. Nutrients, 12(9), 2761. https://doi.org/10.3390/nu12092761