Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats

Abstract
Abbreviations Used
| EPA | eicosapentaenoic acid |
| HDL/LDL | high density lipoproteins/low density lipoproteins |
| TG | triglyceride |
| VLDL-C | very low density lipoprotein- cholesterol |
1. Introduction
2. Materials and Methods

{kind=link}
| Ingredient | AP (% dry weight) | Biomass ( % dry weight) | Pectin ( % dry weight) | Cellulose ( % dry weight) |
|---|---|---|---|---|
| Soluble fibers | 37 | 8.5 | 89 | - |
| Insoluble fibers | 8 | 27 | - | 98 |
| Total dietary fibers | 45 | 35.5 | 89 | 98 |
| Carbohydrate | 26.5 | 27 | - | - |
| Protein | 15 | 18 | 7 | 1.9 |
| Lipids | 2.5 | 6 | - | - |
| Ash | 11 | 13.5 | 4 | 0.1 |
| Total dietary fibers in diet ( %) | 6.5 | 6.5 | 6.5 | 2 |
| Ingredient | AP (g·kg-1) | Biomass (g·kg-1) | Pectin (g·kg-1) | Control (g·kg-1) |
|---|---|---|---|---|
| Casein | 178 | 171 | 196 | 200 |
| Sucrose | 232 | 232 | 232 | 232 |
| Corn starch | 325 | 290 | 374 | 420 |
| Corn oil | 67 | 59 | 70 | 70 |
| Fiber source | 140 | 190 | 70 | 20 |
| Vitamin mix* | 10 | 10 | 10 | 10 |
| Salt mix* | 35 | 35 | 35 | 35 |
| DL-methionine | 3 | 3 | 3 | 3 |
| Cholesterol | 10 | 10 | 10 | 10 |
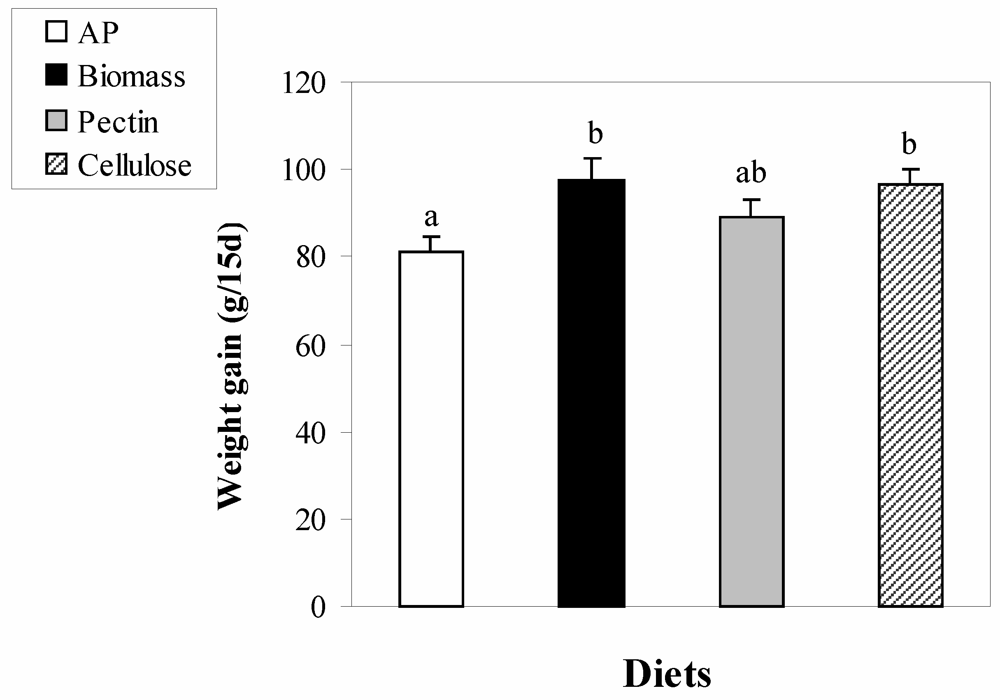
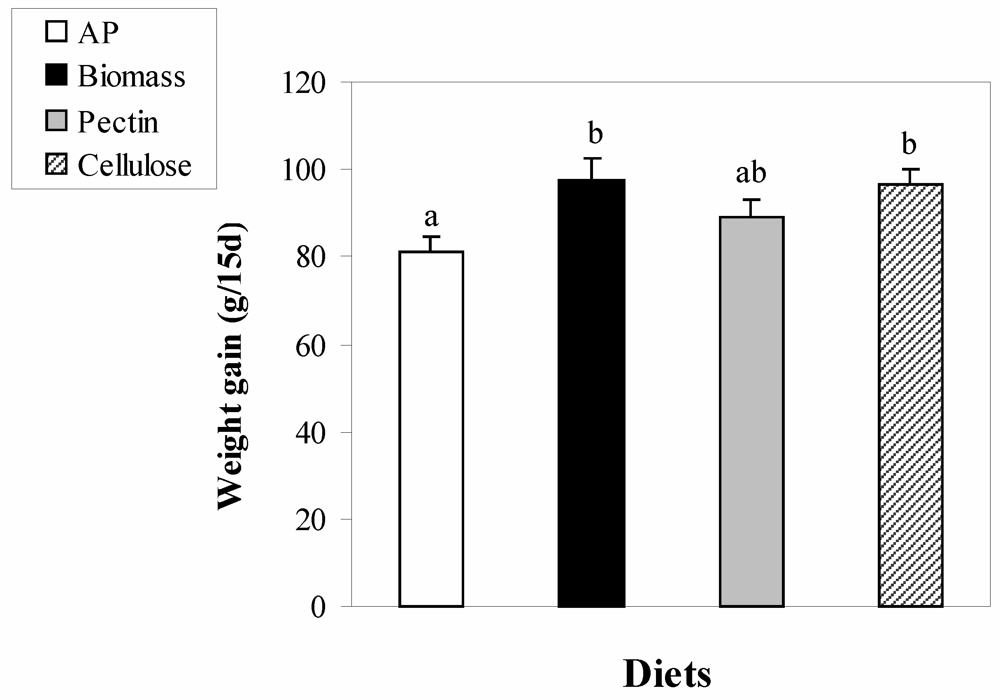
3. Results
| Study group | Plasma cholesterol (mg/dL) | Plasma triglycerides (mg /dL) | VLDL-C (mg/dL) | HDL/LDL | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| Polysaccharide | 103.17 | 5.28 a | 23.17 | 1.52 a | 46.7 | 2.9 a | 0.77 | 0.12 a |
| Biomass | 95.67 | 4.62 a | 26.67 | 1.71 a b | 53.3 | 3.4ab | 0.83 | 0.07 b |
| Pectin | 132.50 | 8.62b | 38.00 | 2.31 c | 76.0 | 4.6c | 0.53 | 0.08 bc |
| Control | 144.00 | 12.12 b | 30.17 | 3.06 b | 60.3 | 6.1b | 0.33 | 0.05 c |
| Study group | Fecal dry weight (g·day-1) | Fecal neutral sterol (g·day-1) | Fecal bile acids (g·day-1) | |||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | |
| Polysaccharide | 3.17 | 0.08 a | 1.93 | 0.03 a | 0.33 | 0.07 a |
| Biomass | 2.46 | 0.08 b | 1.19 | 0.03 b | 1.31 | 0.18 b |
| Pectin | 0.93 | 0.06 c | 0.82 | 0.04 bc | 0.59 | 0.07 a |
| Control | 1.07 | 0.05 c | 0.59 | 0.03 c | 0.61 | 0.08 a |
| Study group | Liver weight (g) | Liver weight (mg·g body weight-1) | Hepatic cholesterol (mg g-1) | |||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | |
| Polysaccharide | 7.65 | 0.28 a | 35.39 | 0.81 a | 1.89 | 0.15 a |
| Biomass | 8.90 | 0.35 b | 37.76 | 1.28 a | 2.14 | 0.05 a |
| Pectin | 8.69 | 0.37 ab | 38.08 | 0.88 a | 1.81 | 0.22 a |
| Control | 10.24 | 0.40 c | 42.86 | 0.85 b | 3.89 | 0.5 b |
4. Discussion
Acknowledgements
References
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar]
- Wu, S.H.; Wen, T.N.; Pan, C.L. Algal-oligosaccharide-lysates prepared by two bacterial agarases stepwise hydrolyzed and their anti-oxidative properties. Fisheries Sci. 2005, 71, 1149–1159. [Google Scholar]
- Dvir, I.; Chayoth, R.; Sod-Moriah, U.; Shany, S.; Nyska, A.; Stark, A.H.; Madar, Z.; Arad, S.M. Soluble polysaccharide and biomass of red microalga Porphyridium sp. alter intestinal morphology and reduce serum cholesterol in rats. Br. J. Nutr. 2000, 84, 469–476. [Google Scholar] [PubMed]
- Bocanegra, A.; Nieto, A.; Bastida, S.; Benedi, J.; Sánchez-Muniz, F.J.A. Nori but not a Konbu, dietary supplement decreases the cholesterolaemia, liver fat infiltration and mineral bioavailability in hypercholesterolaemic growing Wistar rats. Br. J. Nutr. 2008, 99, 272–280. [Google Scholar] [PubMed]
- Frestedt, J.L.; Kuskowski, M.A.; Zenk, J.L. A natural seaweed derived mineral supplement (Aquamin F) for knee osteoarthritis: a randomised, placebo controlled pilot study. Nutr. J. 2009, 2, 7. [Google Scholar]
- Michel, C.; Macfarlane, G.T. Digestive fates of soluble polysaccharide from marine macroalgae: involvement of the colonic microflora and physiological consequences for the host. J. Appl. Bacteriol. 1996, 80, 349–369. [Google Scholar]
- Tannin-Spitz, T.; Bergman, M.; van-Moppes, D.; Grossman, S.; Arad, M.S. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Matsui, S.M.; Muizzudin, N.; Arad, S.; Marenus, K. Sulfated polysaccharides from red microalgae anti-inflammatory properties in vitro and in vivo. Appl. Biochem. Biotechnol. 2003, 104, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Arad, M.S.; Ginzberg, A.; Huleihel, M. Biomaterials from Aquatic and Terrestrial Organisms; Fingerman, M., Nagabhushanam, R., Eds.; Science Publishers Inc: Enfield, NH, USA, 2006; pp. 37–62. [Google Scholar]
- Yaron, A.; Dvir, I.; Maislos, M.; Mokady, S.; Arad (Malis), S. Food Flavors: Generation, Analysis and Process Influence; Charalambous, G., Ed.; Elsevier: Amsterdam, The Netherlands, 1995; pp. 665–674. [Google Scholar]
- Jimenez-Escrig, A.; Goni Cambrodon, I. Nutritional evaluation and physiological effects of edible seaweeds. Arch. Latinoam. Nutr. 1999, 49, 114–120. [Google Scholar]
- Lahaye, M. Marine algae as sources of fibers: Determination of soluble and insoluble dietary fiber contents in some “sea vegetables”. J. Sci. Food Agr. 1991, 54, 587–594. [Google Scholar]
- Nishimune, T.; Sumimoto, T.; Yakusiji, T.; Kunita, N.; Ichikawa, T.; Doguchi, M.; Nakahara, S. Determination of total dietary fibers in Japanese foods. J. Assoc. Off. Anal. Chem. 1991, 74, 350–359. [Google Scholar]
- Lahaye, M.; Jegou, D. Chemical and physical-chemical characteristics of dietary fibers from Ulva lactuca (L.) Thuret and Enteromorpha compressa (L.) Grev. J. Appl. Physiol. 1993, 5, 195–200. [Google Scholar]
- Lairon, D. Dietary fibres: effects on lipid metabolism and mechanisms of action. Eur. J. Clin. Nutr. 1996, 50, 125–133. [Google Scholar]
- Eastwood, M.A.; Morris, E.R. Physical properties of dietary fiber that influence physiological function: a model for polymers among the gastrointestinal tract. Am. J. Clin. Nutr. 1992, 55, 436–442. [Google Scholar]
- Anderson, J.W.; Jones, A.E.; Riddell-Mason, S. Ten different dietary fibers have significantly different effect on serum and liver lipids of cholesterol-fed rats. J. Nutr. 1994, 124, 78–83. [Google Scholar]
- Ara, J.; Sultana, V.; Qasim, R.; Ahmad, V.U. Hypolipidaemic activity of seaweed from Karachi coast. Phytother. Res. 2002, 16, 479–483. [Google Scholar]
- Werman, M.J.; Sukenik, A.; Mokady, S. Effects of the marine unicellular alga Nannochloropsis sp. to reduce the plasma and liver cholesterol levels in male rats fed on diets with cholesterol. Biosci. Biotech. Bioch. 2003, 67, 2266–2268. [Google Scholar] [CrossRef]
- Southgate, D.A.T. Dietary Fiber: Chemical and Biological Aspects; Southgate, D.A.T., Waldron, K., Johnson, I.T., Fenwick, G.R., Eds.; The Royal Society of Chemistry: Cambridge, UK, 1990; pp. 10–19. [Google Scholar]
- Arad (Malis), S. Algal Biotechnology; Stadler, T., Mollion, J., Verdus, M.D., Karamanos, Y., Morvan, H., Christiaen, D., Eds.; Elsevier Applied Science: London, UK, 1988; pp. 65–87. [Google Scholar]
- Geresh, S.; Arad (Malis), S. The extracellular polysaccharide of red microalgae: chemistry and rheology. Bioresource Technol. 1991, 38, 195–201. [Google Scholar]
- Arad, S.; Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Publishing: Oxford, England, 2004; pp. 289–297. [Google Scholar]
- Bajpai, P.; Bajpai, P.K. Eicosapentaenoic acid (EPA) production from microorganisms: a review. Biotechnology 1993, 30, 161–183. [Google Scholar]
- Nuutila, A.M.; Aura, A.M.; Kiesvaara, M.; Kauppienen, V. The effect of salinity, nitrate concentration, pH and temperature on eicosapentaenoic acid (EPA) production by red unicellular alga Porphyridium purpureum. Biotechnol. 1997, 55, 55–63. [Google Scholar]
- Ginzberg, A.; Cohen, M.; Sod-Moriah, U.A.; Shany, S.; Rosenshtrauch, A.; Arad (Malis), S. Chickens fed with biomass of the red microalga Porphyridium sp. have reduced blood cholesterol level and modified fatty acid composition in egg yolk. J. Appl. Phycol. 2000, 12, 325–330. [Google Scholar] [CrossRef]
- Jones, T.M.; Anderson, A.J.; Albersheim, P. Studies on the growth of the red alga Porphyridium cruentum. Plant Physiol. 1963, 16, 636–643. [Google Scholar] [CrossRef]
- Cohen, E.; Arad (Malis), S. A closed system for outdoor cultivation of Porphyridium. Biomass 1989, 18, 59–67. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rabers, P.A.; Smith, F. Colorimetric method for determination of sugar and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Kates, M. Techniques of Lipidology; Burdon, R.H., Van Knippenberg, P.H., Eds.; Elsevier: New York, NY, USA, 1986; p. 1. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.T.; Farr, A.I.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 275–365. [Google Scholar]
- Prosky, L.; Asp, N.; Schweizer, T.F.; DeVries, J.W.; Furda, I. Determination of insoluble, soluble and total dietary fiber in food and food products: Interlaboratory study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar]
- Allain, C.C.; Poon, L.S.; Chan, C.S.G.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar]
- Lopes-Virella, M.F.; Stone, P.; Ellis, S.; Colwella, J.A. Cholesterol determination in high-density lipoproteins separated by three different methods. Clin. Chem. 1977, 23, 882–884. [Google Scholar]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar]
- Searcy, R.L.; Bergquist, L.M. A new color reaction for quantitation of serum cholesterol. Clin. Chem. Acta 1960, 5, 192–199. [Google Scholar]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Sheltawy, M.J.; Losowsky, M.S. Determination of feacal bile acids by an enzymatic method. Clin. Chim. Acta 1975, 64, 127–132. [Google Scholar]
- Amano, H.; Kakinuma, M.; Coury, D.A.; Ohno, H.; Hara, T. Effects of a seaweed mixture on serum lipid level and platelet aggregation in rats. Fisheries Sci. 2005, 71, 1160–1166. [Google Scholar]
- Pengzhan, Y.; Ning, L.; Xiguang, L.; Gefei, Z.; Quanbin, Z.; Pengcheng, L. Antihyperlipidemic effects of different molecular weight sulfated polysaccharides from Ulva Pertusa (chlorophyta). Pharmacol. Res. 2003, 48, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Castro, I.A.; Barroso, L.P.; Sinnecker, P. Functional foods for coronary heart disease risk reduction: a meta-analysis using a multivariate approach. Am. J. Clin. Nutr. 2005, 82, 32–40. [Google Scholar] [PubMed][Green Version]
- Oakenfull, D. Handbook of Dietary Fibers; Cho, S.S., Dreher, M.D., Eds.; Marcel Dekker Inc: New York, NY, USA, 2001; pp. 195–206. [Google Scholar]
- Marlett, J. Handbook of Dietary Fibers; Cho, S.S., Dreher, M.D., Eds.; Marcel Dekker, Inc: New York, NY, USA, 2001; pp. 17–30. [Google Scholar]
- Glore, S.R.; Van Treeck, D.; Knehans, A.W.; Guild, M. Soluble fiber and serum lipids: a literature review. J. Am. Diet. Assoc. 1993, 94, 425–436. [Google Scholar]
- Yan, E.; Durazo, F.; Tong, M.; Hong, K. Nonalcoholic fatty liver disease: pathogenesis, identification, progression, and management. Nutr. Rev. 2007, 65, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Eteshola, E.; Karpasas, M.; Arad (Malis), S.; Gottlieb, M. Red microalga exopolysaccharides: 2. Study of the rheology, morphology and thermal gelation of aqueous preparations. Acta. Polym. 1998, 49, 549–556. [Google Scholar] [CrossRef]
- Eteshola, E.; Gottlieb, M.; Arad (Malis), S. Dilute solution viscosity of red microalga exopolysaccharide. Chem. Eng. Sci. 1996, 51, 1487–1494. [Google Scholar]
- Arad (Malis), S.; Rapoport, L.; Moshkovich, A.; van Moppes, D.; Karpasas, M.; Golan, R.; Golan, Y. A superior biolubricant froma species of red microalga. Langmuir 2006, 15, 7313–7317. [Google Scholar]
- Stark, A.; Nyska, A.; Madar, Z. Metabolic and morphometric changes in small and large intestine in rats fed high-fiber diets. Toxicol. Pathol. 1996, 24, 166–171. [Google Scholar]
- Wang, C.; Yang, G. Comparison of effects of two kinds of soluble algae polysaccharide on blood lipid, liver lipid, platelet aggregation and growth in rats. Zhonghua Yu Fang Yi Xue Za Zhi 1997, 31, 342–345. [Google Scholar] [PubMed]
- Schneeman, B.O.; Richter, D. Changes in plasma and hepatic lipids, small intestinal histology and pancreatic enzyme activity due to aging and dietary fibre in rats. J. Nutr. 1993, 123, 1328–1337. [Google Scholar]
- Lupton, J.R.; Turner, N.D. Dietary fiber and coronary disease: does the evidence support an association? Curr. Atheroscler. Rep. 2003, 5, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Chau, C.F.; Huang, Y.L. Effects of the insoluble fiber derived from Passiflora edulis seed on plasma and hepatic lipids and fecal output. Mol. Nutr. Food Res. 2005, 49, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Zunft, H.J.; Luder, W.; Harde, A.; Haber, B.; Graubaum, H.J.; Koebnick, C.; Grunwald, J. Carob pulp preparation rich in insoluble fibre lowers total and LDL cholesterol in hypercholesterolemic patients. Eur. J. Nutr. 2003, 24, 235–242. [Google Scholar]
- Nagaoka, S.; Shimizu, K.; Kaneko, H.; Shibayama, F.; Morikawa, K.; Kanamaru, Y.; Otsuka, A.; Hirahashi, T.; Kato, T. A novel protein C-phycocyanin plays a crucial role in the hypocholesterolemic action of Spirulina platensis concentrate in rats. J. Nutr. 2005, 135, 2425–2430. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients 2009, 1, 156-167. https://doi.org/10.3390/nu1020156
Dvir I, Stark AH, Chayoth R, Madar Z, Arad SM. Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients. 2009; 1(2):156-167. https://doi.org/10.3390/nu1020156
Chicago/Turabian StyleDvir, Irit, Aliza Hannah Stark, Reuven Chayoth, Zecharia Madar, and Shoshana Malis Arad. 2009. "Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats" Nutrients 1, no. 2: 156-167. https://doi.org/10.3390/nu1020156
APA StyleDvir, I., Stark, A. H., Chayoth, R., Madar, Z., & Arad, S. M. (2009). Hypocholesterolemic Effects of Nutraceuticals Produced from the Red Microalga Porphyridium sp. in Rats. Nutrients, 1(2), 156-167. https://doi.org/10.3390/nu1020156