
Scrubbing Up: Multi-Scale Investigation of Woody Encroachment in a Southern African Savannah

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
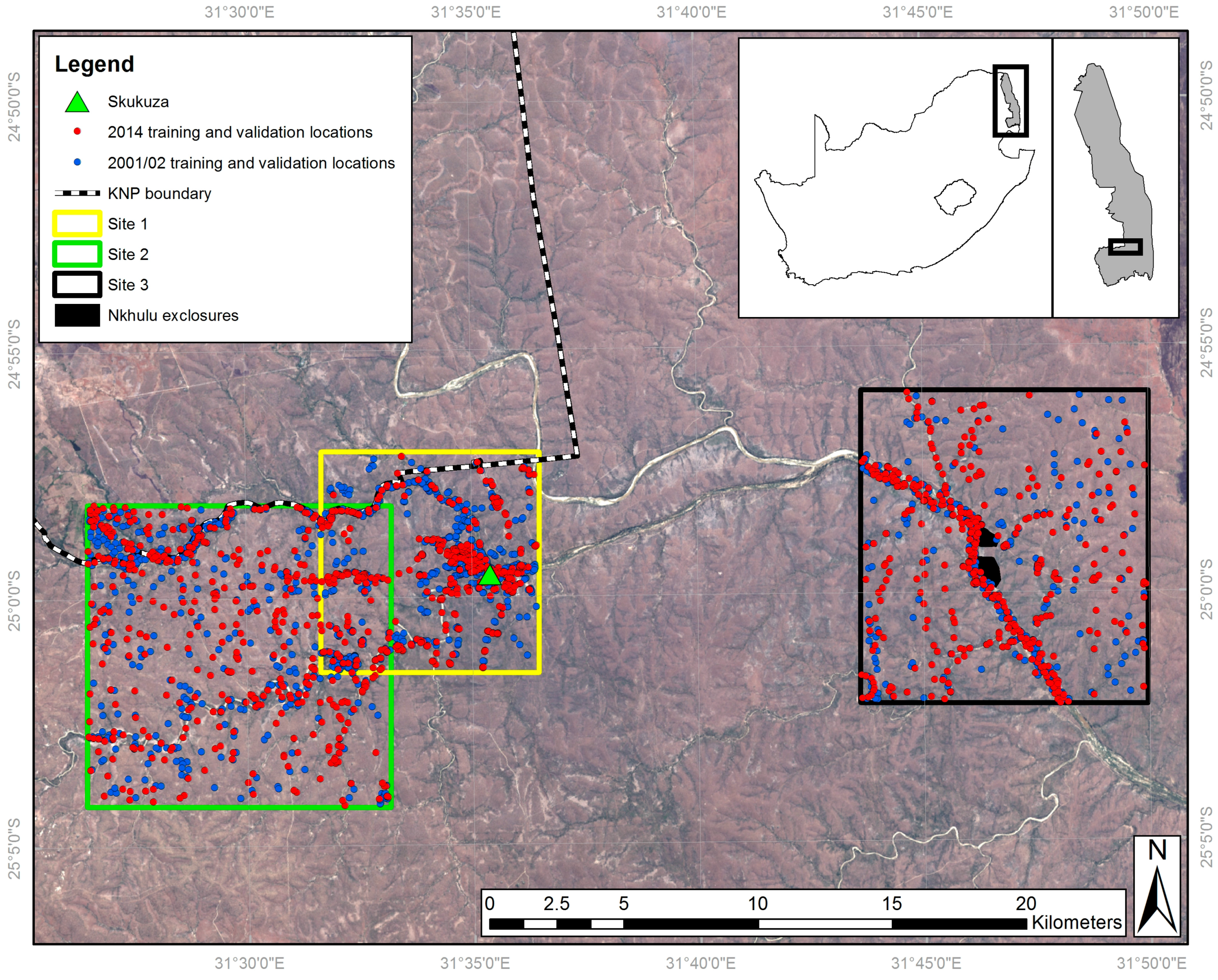
2.1. Study Area
2.2. Land Cover Field Survey
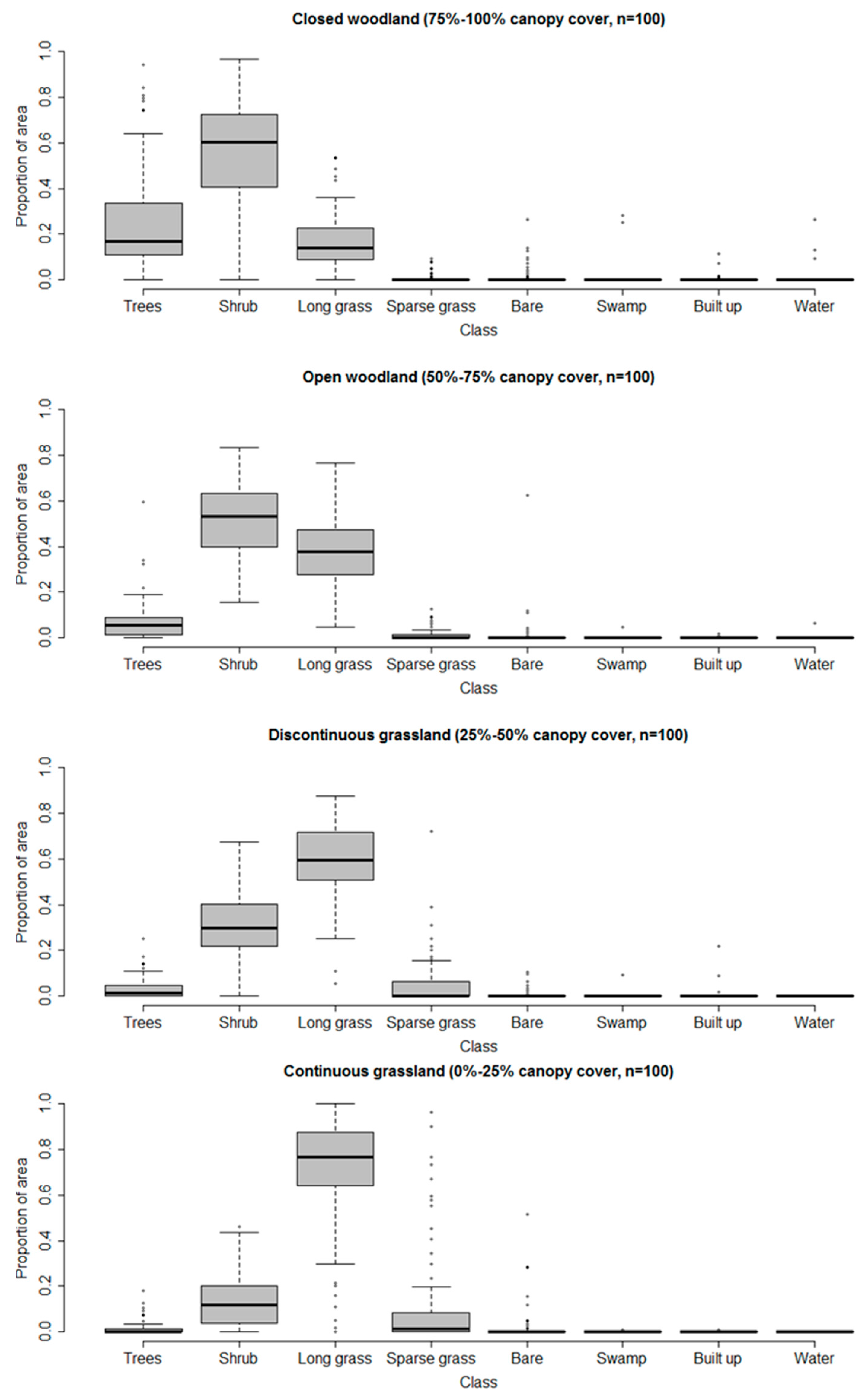
2.3. Classification Nomenclature
2.4. Image Analysis
2.5. Multi-Scale Comparison
2.6. Environmental Drivers of Land Cover Change
3. Results
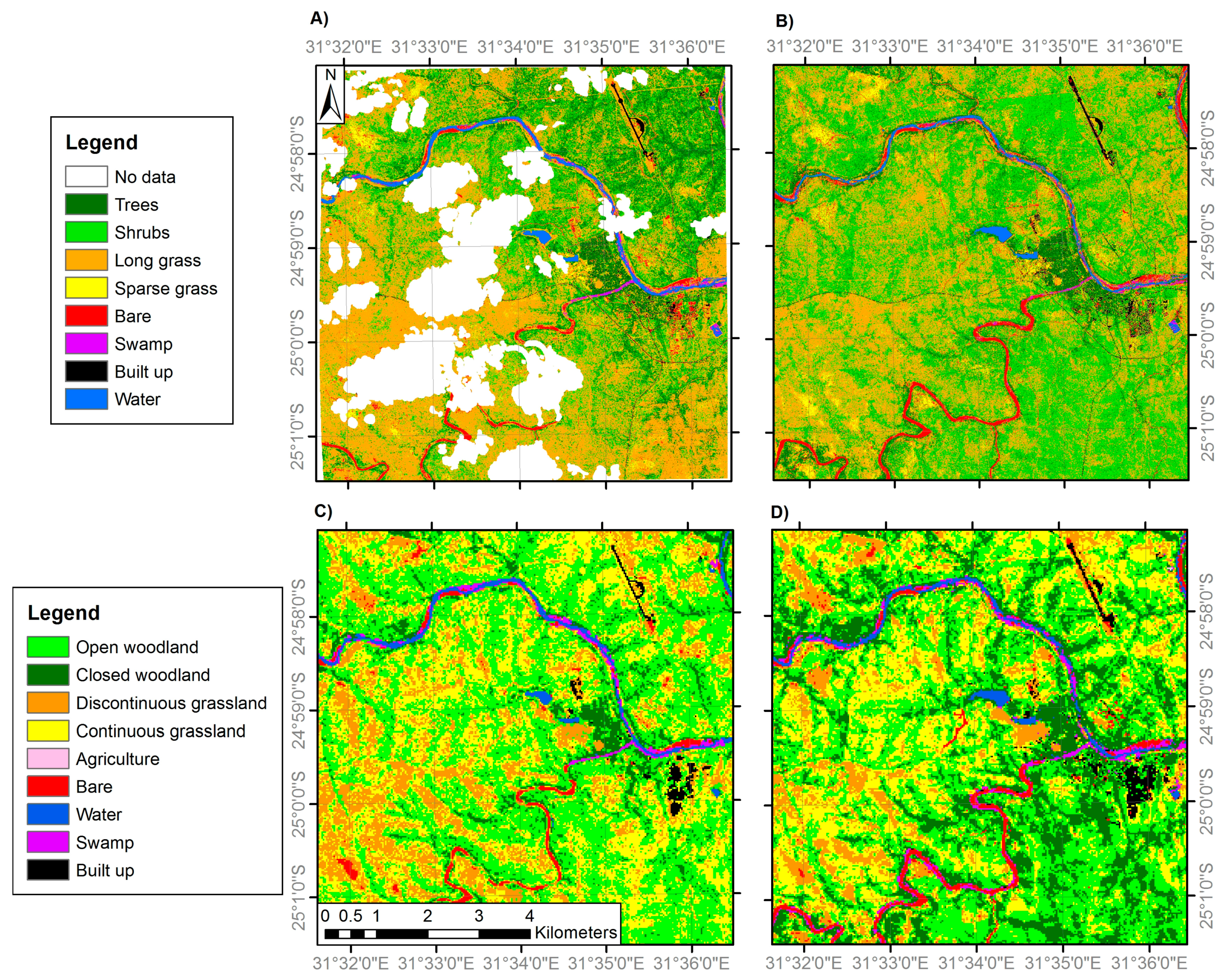
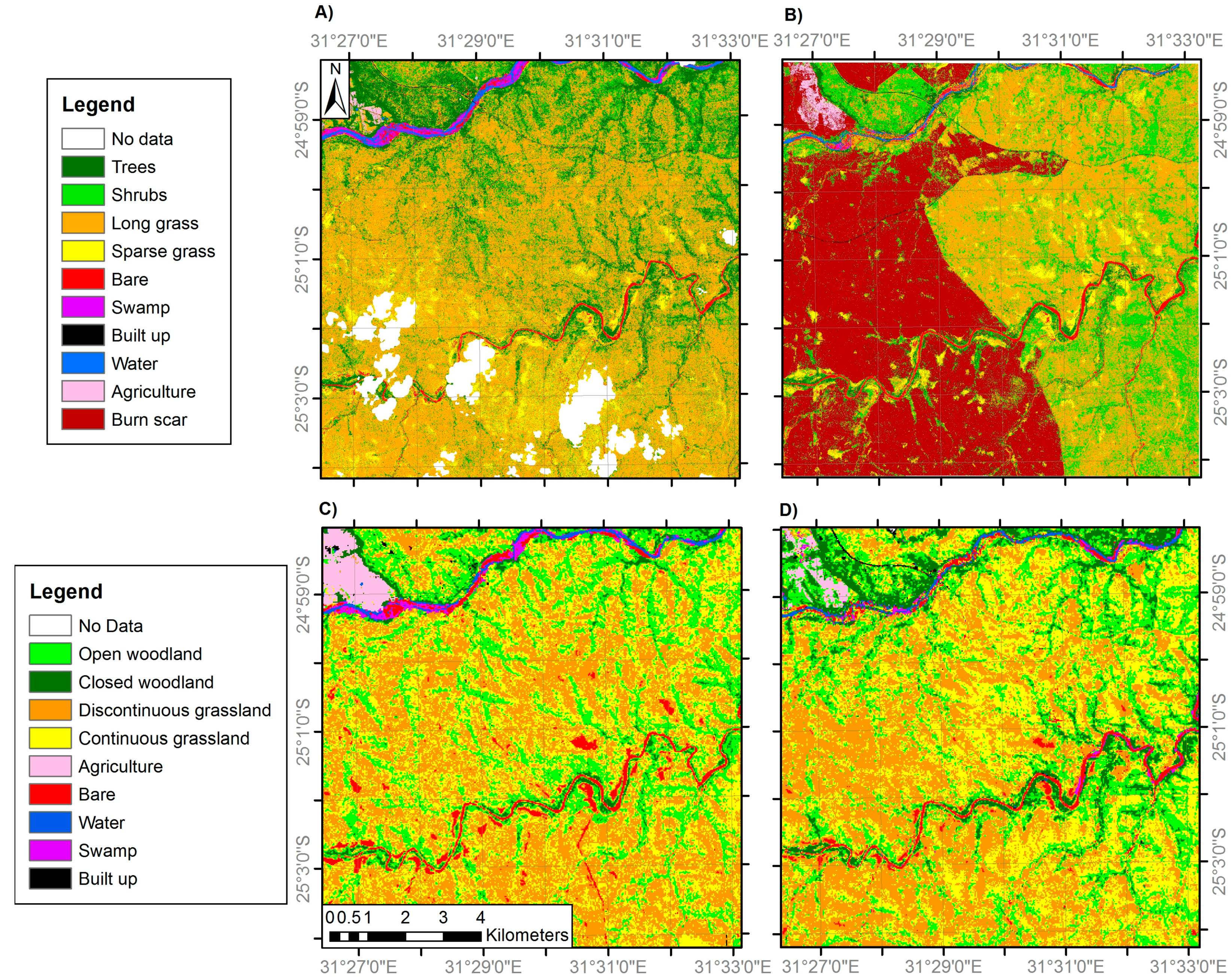
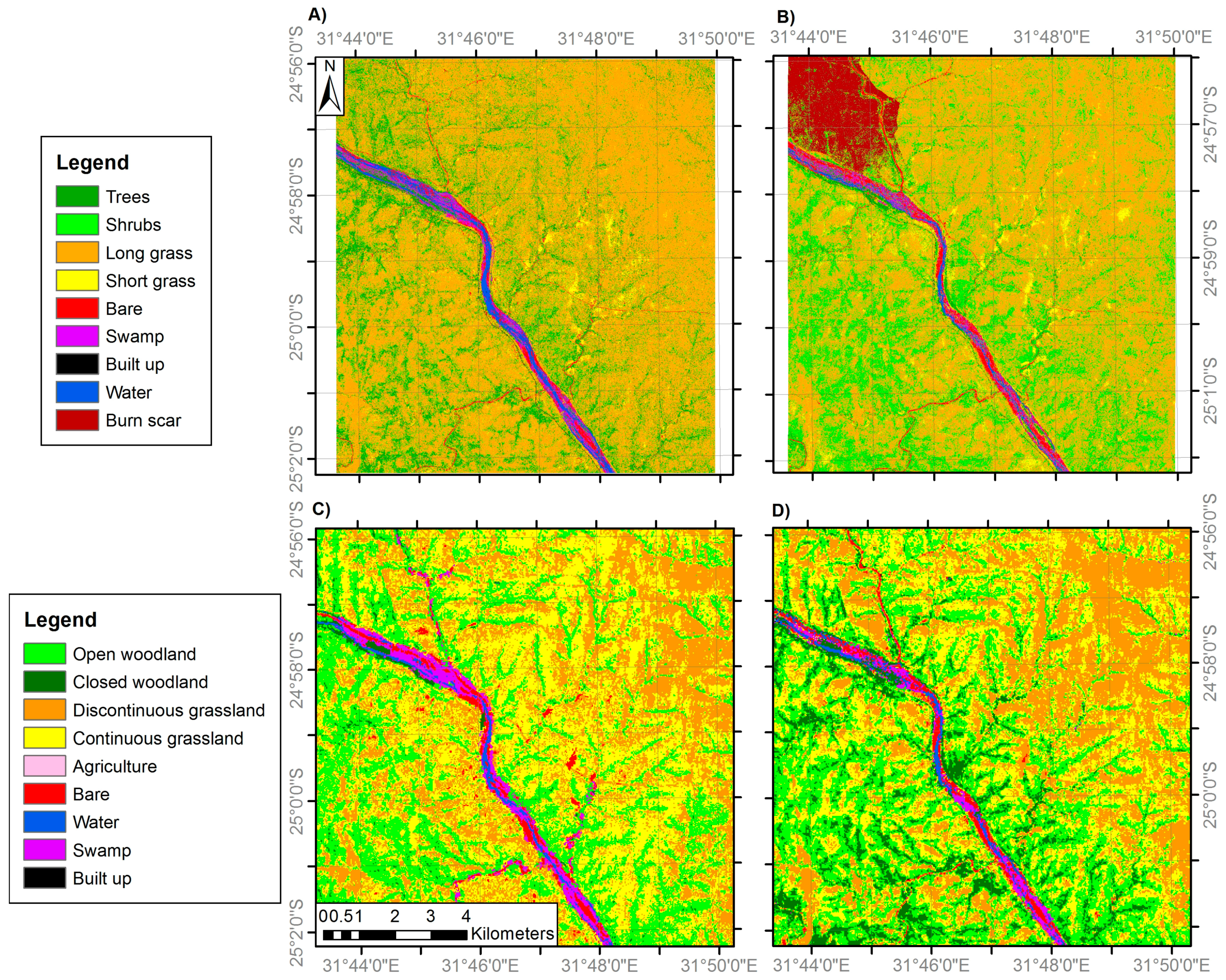
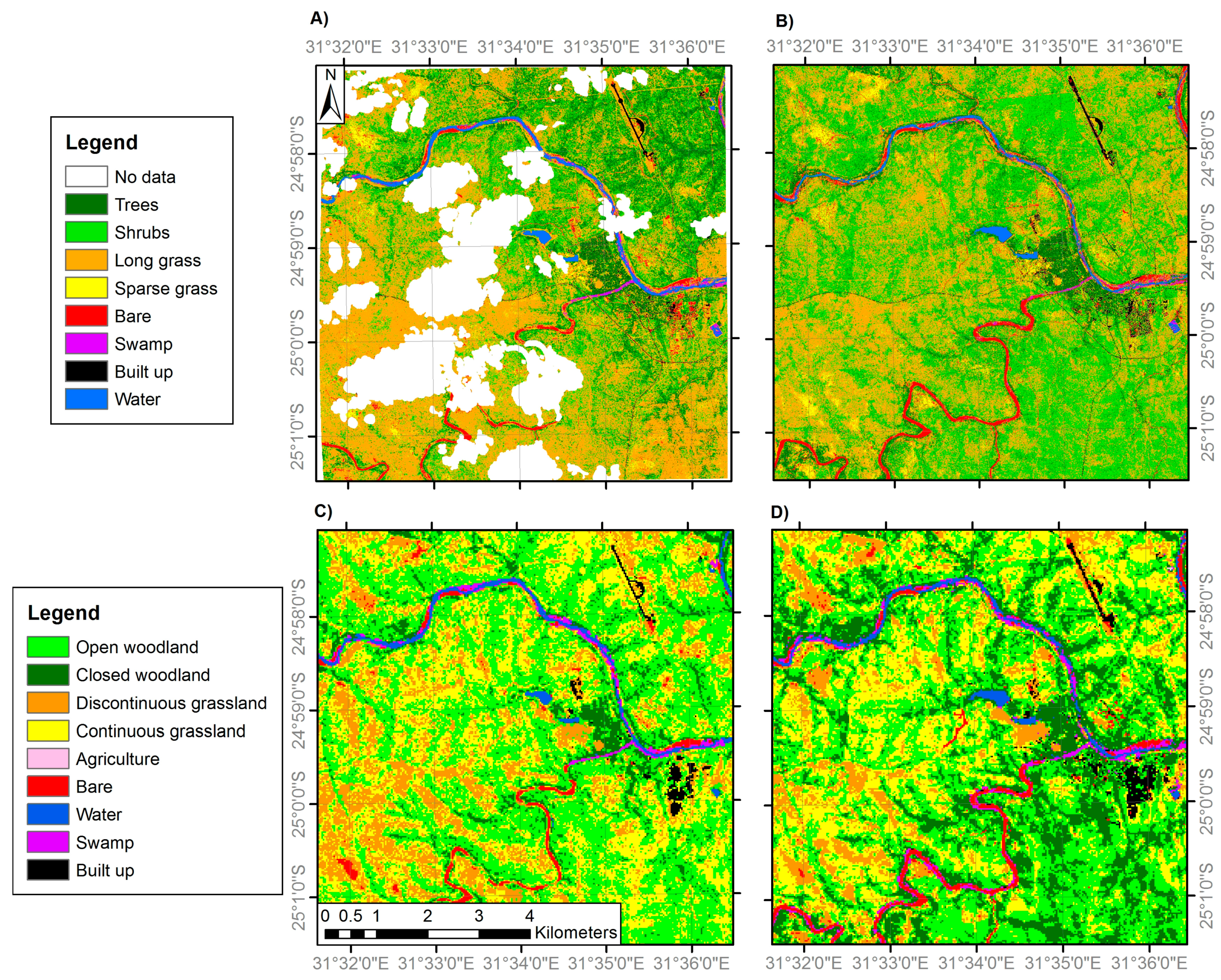
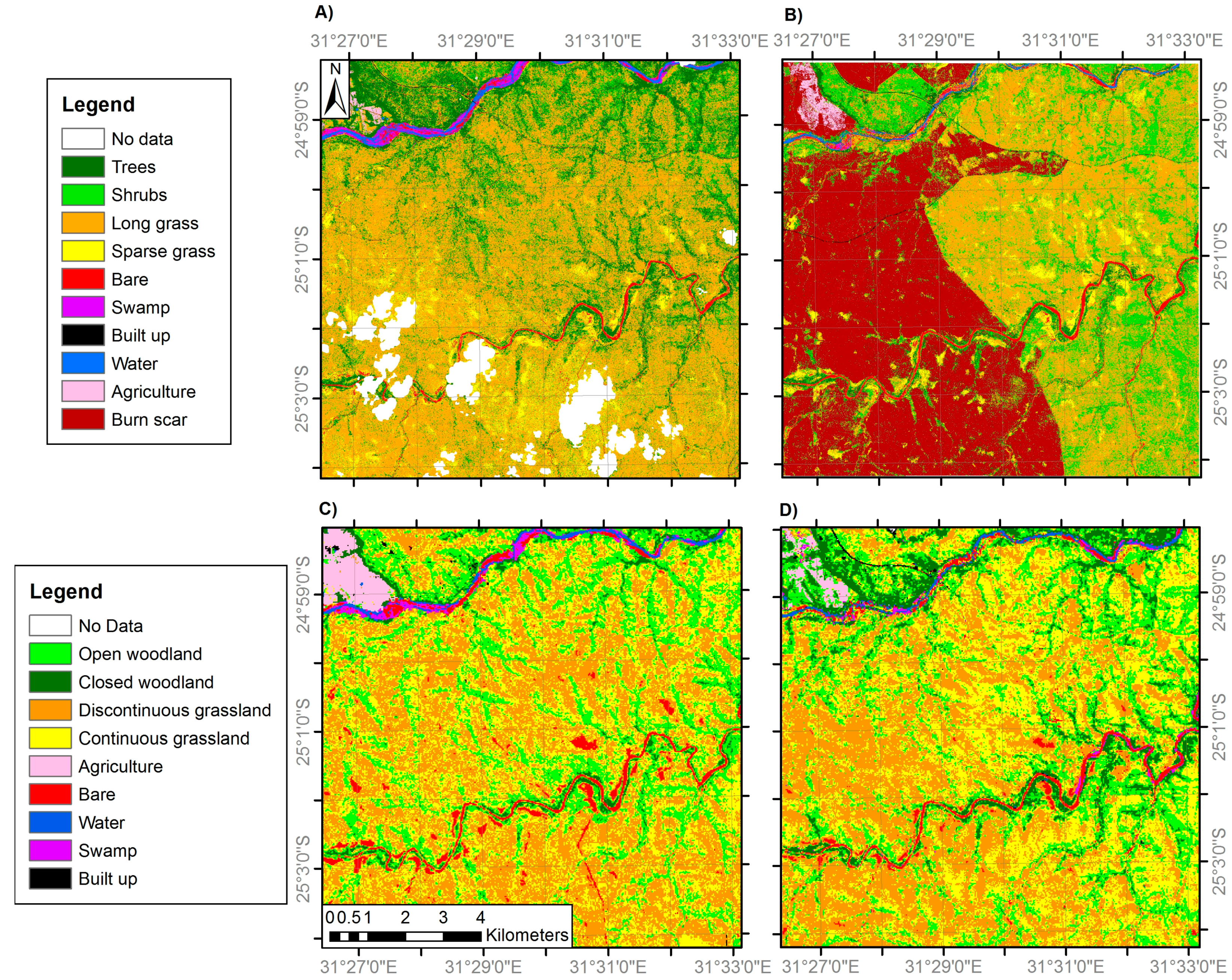
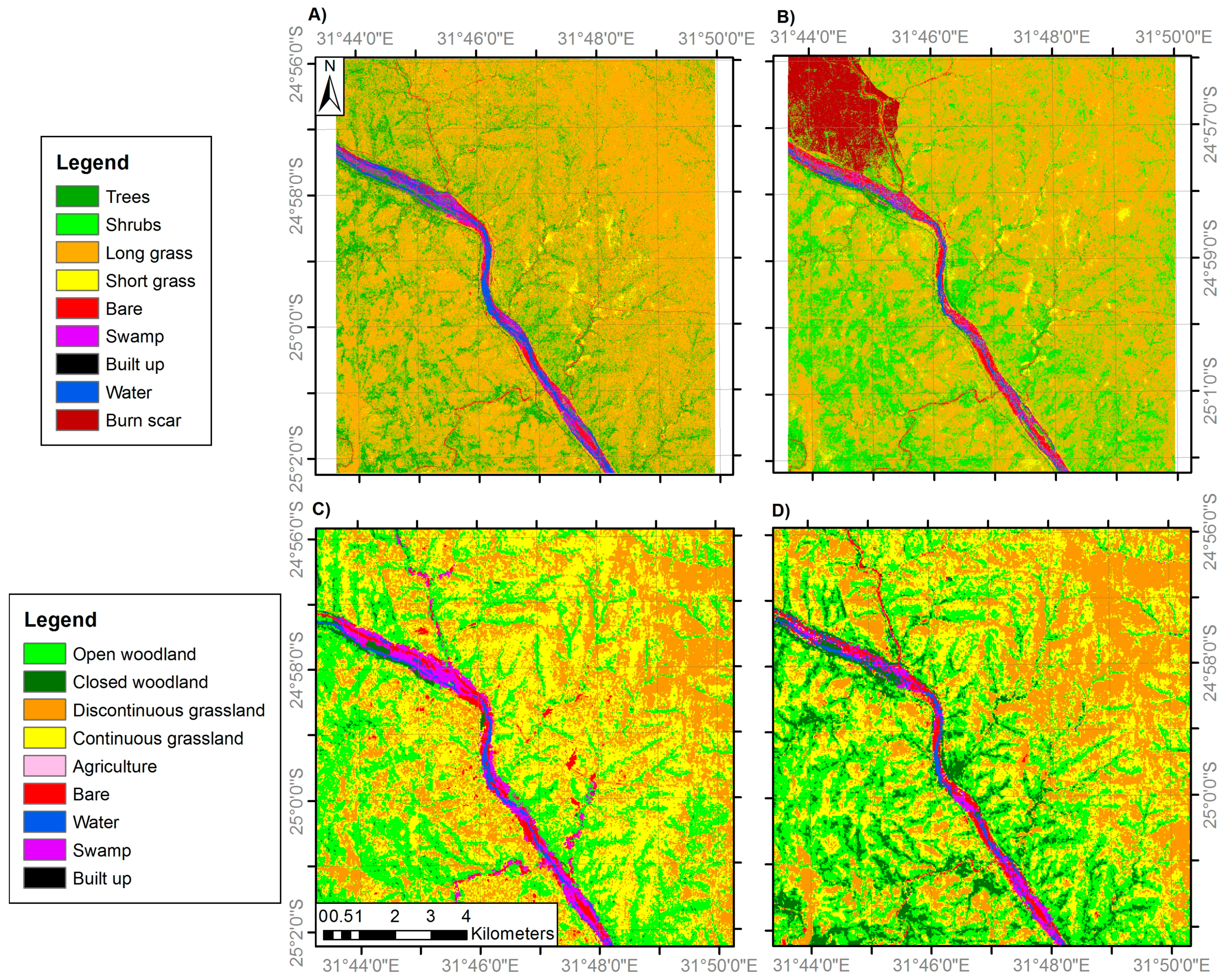
3.1. Land Cover Change
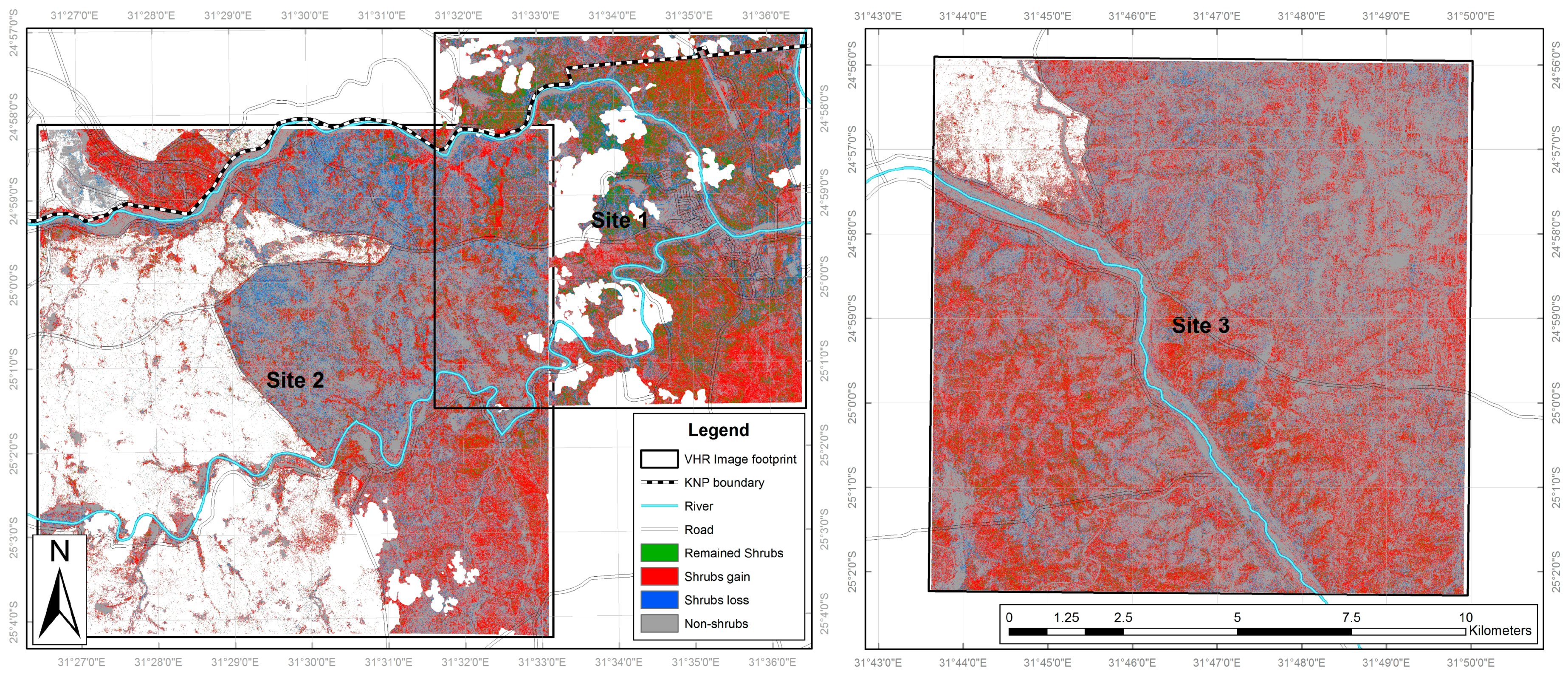
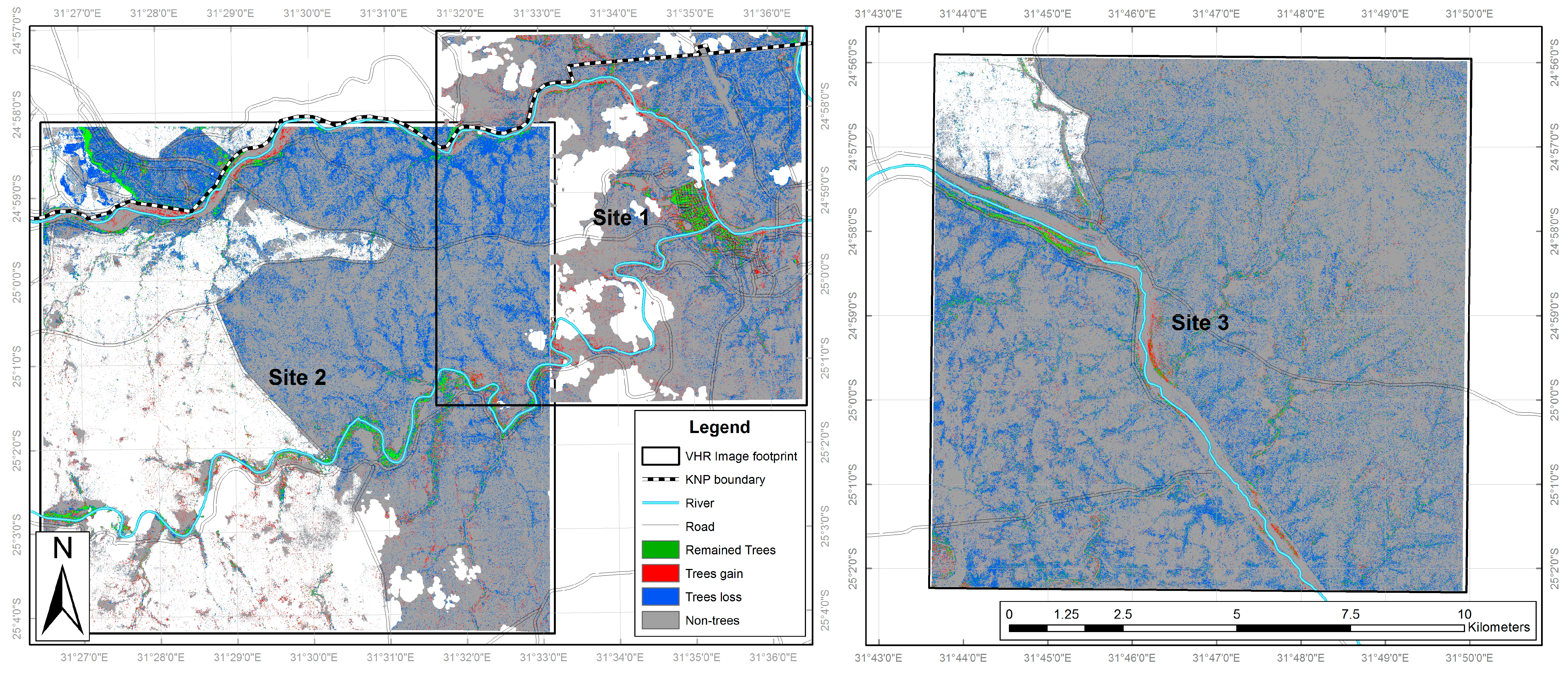
3.2. Nkhulu Exclosure Area
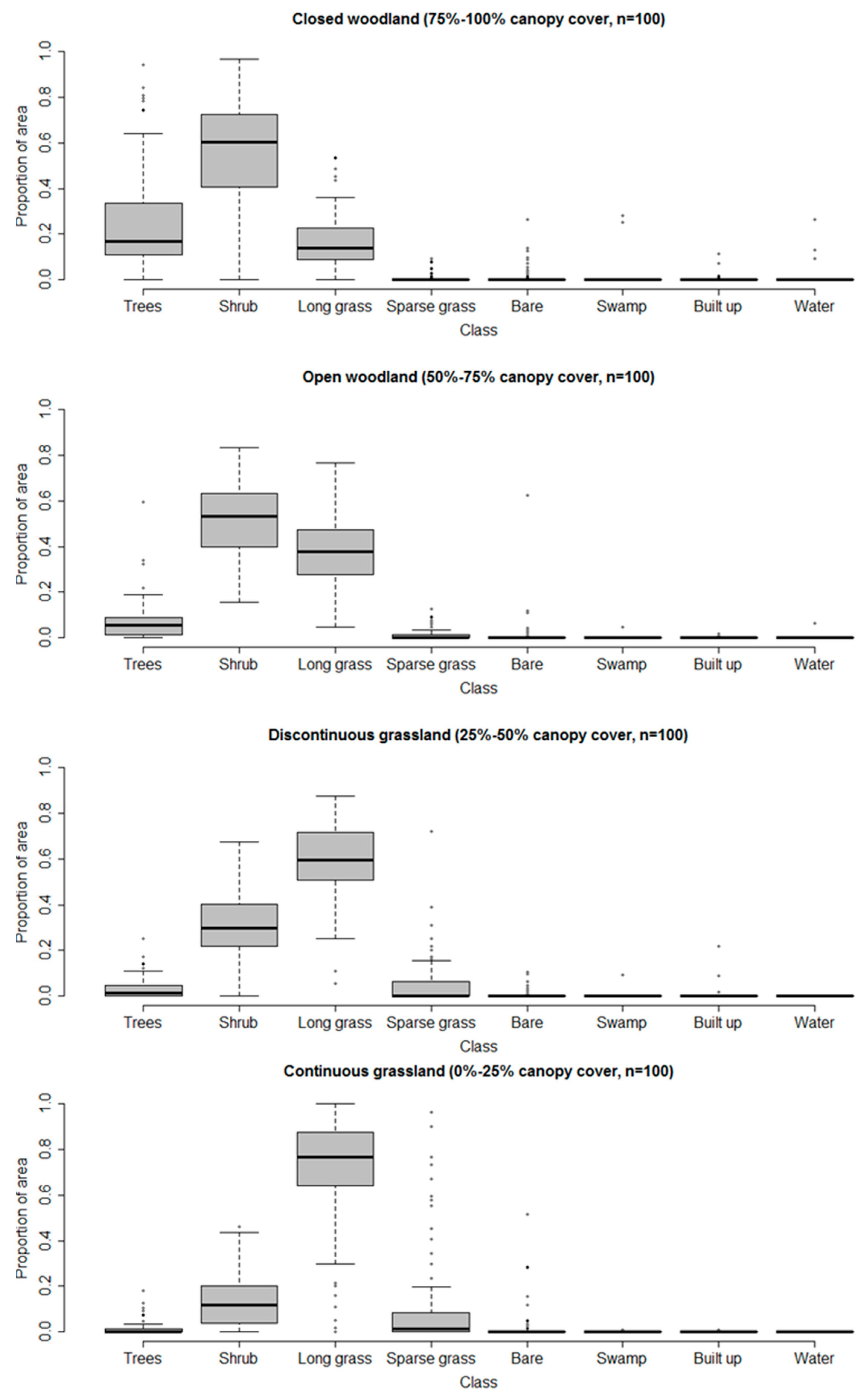
3.3. Multi-Scale Comparison
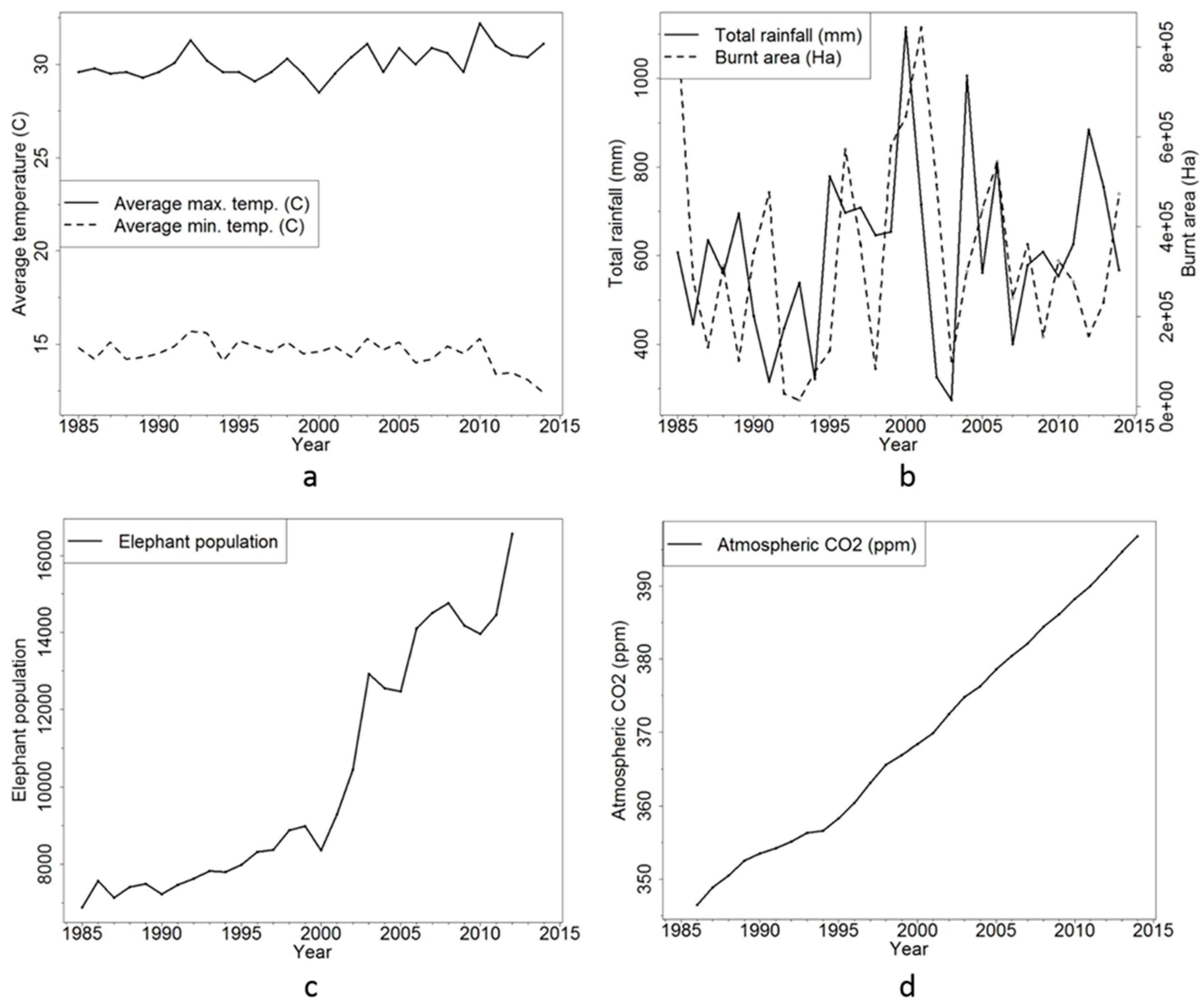
3.4. Environmental Drivers of Land Cover Change
4. Discussion
4.1. Multi-Scale Monitoring of Land Cover Change
4.2. Land Management and Ecological Impacts of Land Cover Change
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Devine, A.P.; Stott, I.; McDonald, R.A.; Maclean, I.M.D. Woody cover in wet and dry African savannas after six decades of experimental fires. J. Ecol. 2015, 103, 473–478. [Google Scholar] [CrossRef]
- Shorrocks, B.; Bates, W. The Biology of African Savannahs, 2nd ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Torello-Raventos, M.; Feldpausch, T.R.; Veenendaal, E.; Schrodt, F.; Saiz, G.; Domingues, T.F.; Djagbletey, G.; Ford, A.; Kemp, J.; Marimon, B.S.; et al. On the delineation of tropical vegetation types with an emphasis on forest/savannah transitions. Plant Ecol. Divers. 2013, 6, 101–137. [Google Scholar] [CrossRef]
- Daskin, J.H.; Stalmans, M.; Pringle, R.M. Ecological legacies of civil war: 35-year increase in savanna tree cover following wholesale large-mammal declines. J. Ecol. 2016, 104, 79–89. [Google Scholar] [CrossRef]
- Buitenwerf, R.; Bond, W.J.; Stevens, N.; Trollope, W.S.W. Increased tree densities in South African savannahs: >50 years of data suggests CO2 as a driver. Glob. Chang. Biol. 2012, 18, 675–684. [Google Scholar] [CrossRef]
- Hirota, M.; Holmgren, M.; Nes, E.H.V.; Scheffer, M. Global resilience of tropical forest and savanna to critical transitions. Science 2011, 334, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Anadon, J.D.; Sala, O.E.; Maestre, F.T. Climate change will increase savannas at the expense of forests and treeless vegetation in tropical and subtropical Americas. J. Ecol. 2014, 102, 1363–1373. [Google Scholar] [CrossRef]
- Zelazowski, P.; Malhi, Y.; Huntingford, C.; Sitch, S.; Fisher, J.B. Changes in the potential distribution of humid tropical forests on a warmer planet. Philos. Trans. R. Soc. A 2011, 369, 137–160. [Google Scholar] [CrossRef] [PubMed]
- Devine, A.P.; McDonald, R.A.; Quaife, T.; Maclean, I.M.D. Determinants of woody encroachment and cover in African savannas. Oecologia 2017, 183, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.M.J.S.; Murphy, B.; Banfai, D.S. Has global environmental change caused monsoon rainforests to expand in the Australian monsoon tropics? Landsc. Ecol. 2010, 25, 1247–1260. [Google Scholar] [CrossRef]
- Fensham, R.J.; Fairfax, R.J. Preliminary assessment of gidgee (Acacia cambagei) woodland thickening in the Longreach district, Queensland. Rangeland J. 2005, 27, 159–168. [Google Scholar] [CrossRef]
- Stevens, N.; Lehmann, C.E.R.; Murphy, B.P.; Durigan, G. Savanna woody encroachment is widespread across three continents. Glob. Chang. Biol. 2017, 23, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Bucini, G.; Hanan, N.P.; Boone, R.B.; Smit, I.P.J.; Saatchi, S.; Lefsky, M.A.; Asner, G.P. Woody fractional cover in Kruger National Park, South Africa: Remote-sensing-based maps and ecological insights. In Ecosystem Function in Savannas: Measurement and Modeling at Landscape to Global Scales; Hill, M.J., Hanan, N.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 219–238. [Google Scholar]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Bucini, G.; Hanan, N.P. A continental-scale analysis of tree cover in African savannas. Glob. Ecol. Biogeogr. 2007, 16, 593–605. [Google Scholar] [CrossRef]
- Trollope, W.S.W.; Tainton, N.M. Effect of fire intensity on the grass and bush components of the Eastern Cape Thornveld. J. Grassl. Soc. S. Afr. 1986, 3, 37–42. [Google Scholar] [CrossRef]
- Bond, W.J. What limits trees in C4 grasslands and savannas? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Bond, W.; Zaloumis, N.P. The deforestation story: Testing for anthropogenic origins of Africa’s flammable grassy biomes. Philos. Trans. R. Soc. B 2016, 371, 20150170. [Google Scholar] [CrossRef] [PubMed]
- Munyati, C.; Sinthumule, N.I. Change in woody cover at representative sites in the Kruger National Park, South Africa, based on historical imagery. SpringerPlus 2016, 5, 1417. [Google Scholar] [CrossRef] [PubMed]
- Smit, I.P.J.; Asner, G.P.; Govender, N.; Vaughn, N.R.; van Wilgen, B.W. An examination of the potential efficacy of high-intensity fires for reversing woody encroachment in savannas. J. Appl. Ecol. 2016, 53, 1623–1633. [Google Scholar] [CrossRef]
- Trollope, W.S.W. Fire in savanna. In Ecological Effects of Fire in South African Ecosystems; Booysen, P.V., Tainton, N.M., Eds.; Springer: Berlin, Germany, 1984; pp. 151–175. [Google Scholar]
- Higgins, S.I.; Bond, W.J.; Trollope, W.S.W. Fire, resprouting and variability: A recipe for grass-tree coexistence in savanna. J. Ecol. 2000, 88, 213–229. [Google Scholar] [CrossRef]
- O’Connor, T.G.; Puttick, J.R.; Hoffman, M.T. Bush encroachment in southern Africa: Changes and causes. Afr. J. Range Forage Sci. 2014, 31, 67–88. [Google Scholar] [CrossRef]
- Laws, R.M. Elephants as agents of habitat and landscape change in East Africa. Oikos 1970, 21, 1–15. [Google Scholar] [CrossRef]
- Asner, G.P.; Levick, S.R.; Kennedy-Bowdoin, T.; Knapp, D.E.; Emerson, R.; Jacobson, J.; Colgan, M.S.; Martin, R.E. Large-scale impacts of herbivores on the structural diversity of African savannas. Proc. Natl. Acad. Sci. USA 2009, 106, 4947–4952. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Levick, S.R. Landscape-scale effects of herbivores on treefall in African savannas. Ecol. Lett. 2012, 15, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.A.; Holdo, R.M.; Anderson, T.M. Elephant damage, not rain or rainfall, explains mortality of overstorey trees in Serengeti. J. Ecol. 2016, 104, 409–418. [Google Scholar] [CrossRef]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, A.T. Grazing systems, ecosystem responses, and global change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Midgley, J.J.; Lawes, M.J.; Chamaillé-Jammes, S. Savanna woody plant dynamics: The role of fire and herbivory, separately and together. Aust. J. Bot. 2010, 58, 1–11. [Google Scholar] [CrossRef]
- Staver, A.C.; Archibald, S.; Levin, S. Tree cover in sub-Saharan Africa: Rainfall and fire constrain forest and savanna as alternative stable states. Ecology 2011, 92, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, M.; Ratnam, J.; Hanan, N.P. Woody cover in African savannas: The role of resources, fire and herbivory. Glob. Ecol. Biogeogr. 2008, 17, 236–245. [Google Scholar] [CrossRef]
- Scholes, R.J.; Archer, S.R. Tree–Grass Interactions in Savannas. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Archer, S.; Schimel, D.S.; Holland, E.A. Mechanisms of shrubland expansion: Land use change, climate or CO2? Clim. Chang. 1995, 29, 91–99. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, G.F. Carbon dioxide and the uneasy interactions of trees and savannah grasses. Philos. Trans. R. Soc. B 2012, 367, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; Midgley, G.F. A proposed CO2-controlled mechanism of woody plant invasion in grasslands and savannas. Glob. Chang. Biol. 2000, 6, 865–869. [Google Scholar] [CrossRef]
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef]
- Stevens, N.; Erasmus, B.F.N.; Archibald, S.; Bond, W.J. Woody encroachment over 70 years in South African savannahs: Overgrazing, global change or extinction aftershock? Philos. Trans. R. Soc. B 2016, 371, 20150437. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.; Hoffman, M.T.; Collocott, S.J. A century of woody plant encroachment in the dry Kimberley savanna of South Africa. Afr. J. Range Forage Sci. 2014, 31, 107–121. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Polley, H.W. Implications of rising atmospheric carbon dioxide concentration for rangelands. J. Range Manag. 1997, 50, 562–577. [Google Scholar] [CrossRef]
- Idso, K.E.; Idso, S.B. Plant responses to atmospheric CO2 enrichment in the face of environmental constraints: A review of the past 10 years’ research. Agr. For. Meteorol. 1994, 69, 153–203. [Google Scholar] [CrossRef]
- Idso, S.B. Shrubland expansion in the American southwest. Clim. Chang. 1992, 22, 85–86. [Google Scholar] [CrossRef]
- Brandt, M.; Rasmussen, K.; Peñuelas, J.; Tian, F.; Schurgers, G.; Verger, A.; Mertz, O.; Palmer, J.R.B.; Fensholt, R. Human population growth offsets climate-driven increase in woody vegetation in sub-Saharan Africa. Nat. Ecol. Evol. 2017, 1, 0081. [Google Scholar] [CrossRef]
- Parr, C.L.; Gray, E.F.; Bond, W.J. Cascading biodiversity and functional consequences of a global change–induced biome switch. Divers. Distrib. 2012, 18, 493–503. [Google Scholar] [CrossRef]
- Sirami, C.; Monadjem, A. Changes in bird communities in Swaziland savannas between 1998 and 2008 owing to shrub encroachment. Divers. Distrib. 2012, 18, 390–400. [Google Scholar] [CrossRef]
- Tambling, C.J.; Minnie, L.; Adendorff, J.; Kerley, G.I. Elephants facilitate impact of large predators on small ungulate prey species. Basic Appl. Ecol. 2013, 14, 694–701. [Google Scholar] [CrossRef]
- Ford, A.T.; Goheen, J.R.; Otieno, T.O.; Bidner, L.; Isbell, L.A.; Palmer, T.M.; Ward, D.; Woodroffe, R.; Pringle, R.M. Large carnivores make savanna tree communities less thorny. Science 2014, 346, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Joffre, R.; Rambal, S. How tree cover influences the water balance of Mediterranean rangelands. Ecology 1993, 74, 570–582. [Google Scholar] [CrossRef]
- Coetsee, C.; Gray, E.F.; Wakeling, J.; Wigley, B.J.; Bond, W.J. Low gains in ecosystem carbon with woody plant encroachment in a South African savanna. J. Trop. Ecol. 2013, 29, 49–60. [Google Scholar] [CrossRef]
- Gray, E.F.; Bond, W.J. Will woody plant encroachment impact on the visitor experience and economy of conservation areas? Koedoe 2013, 55, 9. [Google Scholar] [CrossRef]
- Lindsey, P.A.; Alexander, R.; Mills, M.G.L.; Romañach, S.; Woodroffe, R. Wildlife viewing preferences of visitors to protected areas in South Africa: Implications for the role of ecotourism in conservation. J. Ecotour. 2007, 6, 19–33. [Google Scholar] [CrossRef]
- Bucini, G.; Saatchi, S.; Hanan, N.; Boone, R.B.; Smit, I. Woody cover and heterogeneity in the Savannas of the Kruger National Park, South Africa. In Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009; Volume 4, pp. 334–337. [Google Scholar]
- Helman, D.; Lensky, I.M.; Tessler, N.; Osem, Y. A phenology-based method for monitoring woody and herbaceous vegetation in Mediterranean forests from NDVI time series. Remote Sens. 2015, 7, 12314–12335. [Google Scholar] [CrossRef]
- Holdo, R.M.; Mack, M.C. Functional attributes of savanna soils: Contrasting effects of tree canopies and herbivores on bulk density, nutrients and moisture dynamics. J. Ecol. 2014, 102, 1171–1182. [Google Scholar] [CrossRef]
- Kaszta, Z.; Van De Kerchove, R.; Ramoelo, A.; Cho, M.A.; Madonsela, S.; Mathieu, R.; Wolff, E. Seasonal separation of African savanna components using Worldview-2 imagery: A comparison of pixel and object-based approaches and selected classification algorithms. Remote Sens. 2016, 8, 763. [Google Scholar] [CrossRef]
- Whiteside, T.G.; Boggs, G.S.; Maier, S.W. Comparing object-based and pixel-based classifications for mapping savannas. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 884–893. [Google Scholar] [CrossRef]
- Aplin, P. Remote sensing: Base mapping. Prog. Phys. Geog. 2003, 27, 275–283. [Google Scholar] [CrossRef]
- Sankey, T.; Glenn, N. Landsat-5 TM and Lidar Fusion for Sub-pixel Juniper Tree Cover Estimates in a Western Rangeland. Photogramm. Eng. Remote Sens. 2011, 12, 1241–1248. [Google Scholar] [CrossRef]
- Brandt, M.; Hiernaux, P.; Tagesson, T.; Verger, A.; Rasmussen, K.; Diouf, A.A.; Mbow, C.; Mougin, E.; Fensholt, R. Woody plant cover estimation in drylands from Earth Observation based seasonal metrics. Remote Sens. Environ. 2016, 172, 28–38. [Google Scholar] [CrossRef]
- Dye, D.G.; Middleton, B.R.; Vogel, J.M.; Wu, Z.; Velasco, M. Exploiting differential vegetation phenology for satellite-based mapping of semiarid grass vegetation in the Southwestern United States and Northern Mexico. Remote Sens. 2016, 8, 889. [Google Scholar] [CrossRef]
- Brandt, M.; Tappan, G.; Diouf, A.A.; Beye, G.; Mbow, C.; Fensholt, R. Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel. Remote Sens. 2017, 9, 39. [Google Scholar] [CrossRef]
- Kaptué, A.T.; Prihodko, L.; Hanan, N.P. On regreening and degradation in Sahelian watersheds. Proc. Natl. Acad. Sci. USA 2015, 112, 12133–12138. [Google Scholar] [CrossRef] [PubMed]
- Gessner, U.; Klein, D.; Conrad, C.; Schmidt, M.; Dech, S. Towards an automated estimation of vegetation cover fractions on multiple scales: Examples of Eastern and Southern Africa. In Proceedings of the 33rd International Symposium on Remote Sensing of Environment, Stresa, Italy, 4–8 May 2009. [Google Scholar]
- Scanlon, T.M.; Albertson, J.D.; Caylor, K.K.; Williams, C.A. Determining land surface fractional cover from NDVI and rainfall time series for a savanna ecosystem. Remote Sens. Environ. 2002, 82, 376–388. [Google Scholar] [CrossRef]
- Guldemond, R.; Van Aarde, R. A meta-analysis of the impact of African elephants on savannah vegetation. J. Wildl. Manag. 2008, 72, 892–899. [Google Scholar] [CrossRef]
- Wigley, B.J.; Fritz, H.; Coetsee, C.; Bond, W.J. Herbivores shape woody plant communities in the Kruger National Park: Lessons from three long-term exclosures. Koedoe 2014, 56, 12. [Google Scholar] [CrossRef]
- Du Toit, J.T.; Rogers, K.H.; Biggs, H.C. The Kruger Experience; Ecology and Management of Savanna Heterogeneity; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Pickett, S.T.A.; Cadenasso, M.L.; Benning, T.A. Biotic and abiotic variability as key determinants of savanna heterogeneity at multiple spatiotemporal scales. In The Kruger Experience; Ecology and Management of Savanna Heterogeneity; Du Toit, J.T., Rogers, K.H., Biggs, H.C., Eds.; Island Press: Washington, DC, USA, 2003; pp. 22–40. [Google Scholar]
- Mayaux, P.; Bartholom, E.; Cabral, A.; Cherlet, M.; Defourny, P.; Di Gregorio, A.; Diallo, O.; Massart, M.; Nonguierma, A.; Pekel, J.F.; et al. The Land Cover Map for Africa in the Year 2000. GLC2000 Database, European Commission Joint Research Centre, 2003. Available online: http://www-gem.jrc.it/glc2000 (accessed on 18 April 2015).
- Jenkins, T.A.R.; Mackie, E.D.; Matthews, R.W.; Miller, G.; Randle, T.J.; White, M.E. FC Woodland Carbon Code: Carbon Assessment Protocol. Forestry Commission; 2011. Available online: http://www.forestry.gov.uk/pdf/Carbon_Assessment_Protocol_v1_0_main_22Jul2011.pdf/$file/Carbon_Assessment_Protocol_v1_0_main_22Jul2011.pdf (accessed on 18 April 2015).
- Bezdek, J.C. Fuzzy Mathematics in Pattern Classification. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 1973. [Google Scholar]
- Loveland, T.R.; Reed, B.C.; Brown, J.F.; Ohlen, D.O.; Zhu, Z.; Yang, L.; Merchant, J.W. Development of a global land cover characteristics database and IGBP DISCover from 1 km AVHRR data. Int. J. Remote Sens. 2000, 21, 1303–1330. [Google Scholar] [CrossRef]
- O’Regan, H.J.; Wilkinson, D.M.; Marston, C.G. Hominin home ranges and habitat variability: Exploring modern African analogues using remote sensing. J. Archaeol. Sci. Rep. 2016, 9, 238–248. [Google Scholar] [CrossRef]
- Wayman, J.P.; Wynne, R.H.; Scrivani, J.A.; Reams, G.A. Landsat TM-based forest area estimation using iterative guided spectral class rejection. Photogramm. Eng. Remote. Sens. 2001, 67, 1155–1166. [Google Scholar]
- Ozesmi, S.L.; Bauer, M.E. Satellite remote sensing of wetlands. Wetl. Ecol. Manag. 2002, 10, 381–402. [Google Scholar] [CrossRef]
- Martin, R.; Brabyn, L.; Beard, C. Effects of class granularity and cofactors on the performance of unsupervised classification of wetlands using multi-spectral aerial photography. J. Spat. Sci. 2014, 59, 269–282. [Google Scholar] [CrossRef]
- Jiang, H.; Strittholt, J.R.; Frost, P.A.; Slosser, N.C. The classification of seral forest in the Pacific Northwest USA using Landsat ETM+ imagery. Remote Sens. Environ. 2004, 91, 320–331. [Google Scholar] [CrossRef]
- Baraldi, A.; Puzzolo, V.; Blonda, P.; Bruzzone, L.; Tarantino, C. Automatic spectral rule-based preliminary mapping of calibrated Landsat TM and ETM+ images. IEEE Trans. Geosci. Remote Sens. 2006, 44, 2563–2586. [Google Scholar] [CrossRef]
- Dlugokencky, E.J.; Lang, P.M.; Masarie, K.A.; Crotwell, A.M.; Crotwell, M.J. Atmospheric Carbon Dioxide Dry Air Mole Fractions from the NOAA ESRL Carbon Cycle Cooperative Global Air Sampling Network, 1968–2014. 2015. Available online: ftp://aftp.cmdl.noaa.gov/data/trace_gases/co2/flask/surface/ (accessed on 18 December 2015).
- Eckhardt, H.C.; van Wilgen, B.W.; Biggs, H.C. Trends in woody vegetation cover in the Kruger National Park, South Africa, between 1940 and 1998. Afr. J. Ecol. 2000, 38, 108–115. [Google Scholar] [CrossRef]
- Sankaran, M.; Augustine, D.J.; Ratnam, J. Native ungulates of diverse body sizes collectively regulate long-term woody plant demography and structure of a semi-arid savanna. J. Ecol. 2013, 101, 1389–1399. [Google Scholar] [CrossRef]
- Altamirano, A.; Field, R.; Cayuela, L.; Aplin, P.; Lara, A.; Rey-Benayas, J.M. Woody species diversity in temperate Andean forests: The need for new conservation strategies. Biol. Conserv. 2010, 143, 2080–2091. [Google Scholar] [CrossRef]
- Belsky, A.J.; Canham, C.D. Forest gaps and isolated savanna trees. Bioscience 1994, 44, 77–84. [Google Scholar] [CrossRef]
- Valeix, M.; Fritz, H.; Sabatier, R.; Murindagomo, F.; Cumming, D.; Duncan, P. Elephant-induced structural changes in the vegetation and habitat selection by large herbivores in an African savanna. Biol. Conserv. 2011, 144, 902–912. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
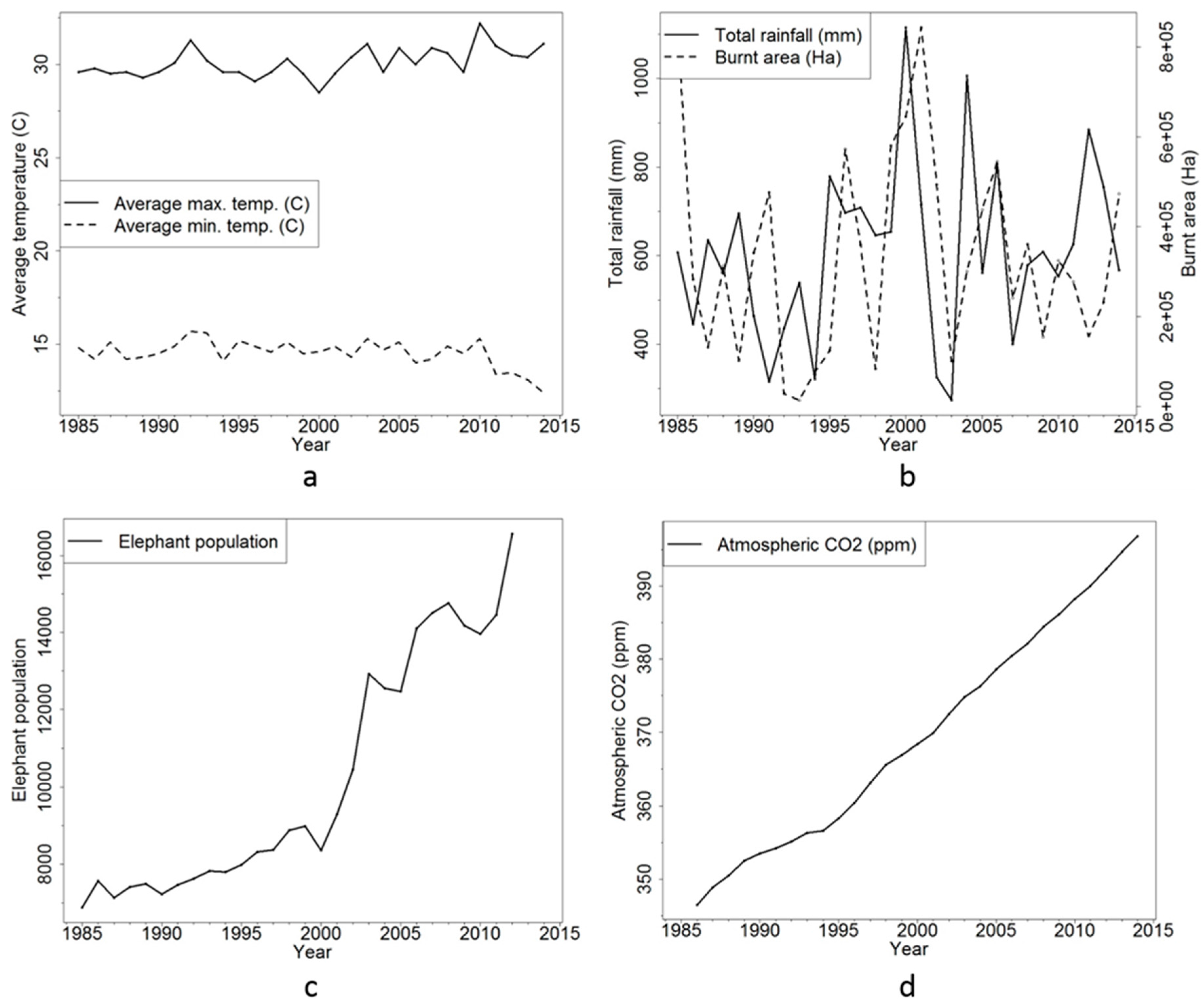{kind=link}
| General Habitat | Medium Resolution Land Cover Classes | VHR Land Cover Classes |
|---|---|---|
| Woodland | 1. Closed woodland (75–100% canopy cover) | 1. Trees (main trunk >7 cm diameter) |
| 2. Open woodland (50–75% canopy cover) | 2. Shrubs (main trunk <7 cm diameter) | |
| Grassland | 3. Discontinuous grassland (25–50% canopy cover) | 3. Long grass |
| 4. Continuous grassland (0–25% canopy cover) | 4. Sparse grass | |
| Anthropogenic classes | 5. Agriculture | 5. Agriculture |
| 6. Built-up | 6. Built-up | |
| Bare | 7. Bare | 7. Bare |
| Water | 8. Water | 8. Water |
| 9. Swamp | 9. Swamp |
| Sensor | Image Acquisition Date | Sites | Spatial Resolution (m) | Spectral Bands | Overall Classification Accuracy (%) |
|---|---|---|---|---|---|
| QuickBird | 9 December 2002 | 1 | 2.8 | B, G, R, NIR | 91.0 |
| IKONOS | 7 March 2002 | 2 | 4 | B, G, R, NIR | 90.6 |
| IKONOS | 28 April 2001 | 3 | 4 | B, G, R, NIR | 87.7 |
| WorldView-2 | 14 October 2014 | 1 | Resampled to 2.8 | CB, B, G, Y, R, RE, NIR1, NIR2 | 90.2 |
| IKONOS | 27 September 2014 | 2 | 4 | B, G, R, NIR | 92.5 |
| IKONOS | 27 September 2014 | 3 | 4 | B, G, R, NIR | 90.6 |
| Landsat ETM+ | 18 June 2002 | 1, 2 and 3 | 30 | B, G, R, NIR, SWIR1, SWIR2 | 75.2 |
| Landsat OLI | 27 June 2014 | 1, 2 and 3 | 30 | CA, B, G, R, NIR, SWIR1, SWIR2, C | 75.2 |
| Class | Site 1 | Site 2 | Site 3 | Sites 1–3 Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | 2001 cover (Ha) | 2014 cover (Ha) | Cover change (%) | February 2001 cover (Ha) | 2014 cover (Ha) | Cover change (%) | |
| Trees | 814.3 | 550.4 | −5.3 | 1801.5 | 502.6 | −17.1 | 1864.3 | 482.3 | −11.8 | 4421.8 | 1299.3 | −13.5 |
| Shrubs | 1440.8 | 2238.7 | +16.1 | 1380.0 | 2652.0 | +16.7 | 1436.6 | 3609.6 | +18.6 | 4029.7 | 8222.7 | +18.1 |
| Long grass | 2272.4 | 1637.4 | −12.8 | 3629.1 | 3585.0 | −0.6 | 7496.9 | 6751.2 | −6.4 | 12,738.1 | 11,556.8 | −5.1 |
| Sparse grass | 164.7 | 215.1 | +1.0 | 424.7 | 466.9 | +0.6 | 400.3 | 426.2 | +0.2 | 955.9 | 1057.1 | +0.4 |
| Bare | 127.3 | 174.7 | +1.0 | 170.2 | 240.8 | +0.9 | 172.2 | 221.4 | +0.4 | 440.7 | 600.0 | +0.7 |
| Swamp | 29.4 | 21.7 | −0.2 | 79.9 | 19.2 | −0.8 | 142.2 | 106.5 | −0.3 | 249.2 | 148.4 | −0.4 |
| Built up | 37.6 | 60.8 | +0.5 | 17.5 | 18.4 | 0.0 | 7.8 | 6.4 | 0.0 | 62.2 | 85.2 | +0.1 |
| Water | 68.6 | 56.4 | −0.3 | 77.3 | 41.5 | −0.5 | 153.0 | 69.6 | −0.7 | 287.0 | 160.0 | −0.5 |
| Agriculture | - | - | - | 29.1 | 84.3 | +0.7 | - | - | - | 29.1 | 84.3 | +0.2 |
| Class | Site 1 | Site 2 | Site 3 | Sites 1–3 Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | 2002 cover (Ha) | 2014 cover (Ha) | Cover change (%) | |
| Closed woodland | 370.3 | 1250.7 | +13.6 | 300.1 | 918.0 | +4.9 | 114.2 | 979.6 | +7.0 | 730.4 | 2960.9 | +7.4 |
| Open woodland | 2713.0 | 2477.4 | −3.6 | 2446.2 | 2605.5 | +1.3 | 3107.2 | 3684.3 | +4.6 | 7820.4 | 8301.7 | +1.6 |
| Discontinuous grassland | 2065.8 | 1696.4 | −5.7 | 4331.7 | 4275.2 | −0.4 | 5249.2 | 3961.8 | −10.4 | 11,106.5 | 9390.9 | −5.7 |
| Continuous grassland | 1044.2 | 651.5 | −6.0 | 4794.9 | 4327.7 | −3.7 | 3299.2 | 3335.2 | +0.3 | 8720.6 | 8074.2 | −2.1 |
| Bare | 107.6 | 154.0 | +0.7 | 390.1 | 273.8 | −0.9 | 244.3 | 216.3 | −0.2 | 705.2 | 608.7 | −0.3 |
| Swamp | 56.8 | 99.8 | +0.7 | 86.8 | 72.1 | −0.1 | 328.6 | 142.0 | −1.5 | 466.6 | 292.5 | −0.6 |
| Built up | 50.2 | 78.3 | 0.4 | 10.7 | 21.8 | +0.1 | 0.1 | 1.1 | 0.0 | 61.1 | 101.3 | +0.1 |
| Water | 85.0 | 84.3 | 0.0 | 67.5 | 67.6 | 0.0 | 85.9 | 108.3 | +0.2 | 226.3 | 245.9 | +0.1 |
| Agriculture | - | - | - | 228.2 | 94.3 | −1.1 | - | - | - | 234.5 | 95.0 | −0.5 |
| Class | Nkhulu Exclosure | Site 3 Excluding Nkhulu Exclosure | ||||
|---|---|---|---|---|---|---|
| 2001 Cover (Ha) | 2014 Cover (Ha) | Cover Change (%) | 2001 Cover (Ha) | 2014 Cover (Ha) | Cover Change (%) | |
| Trees | 21.6 | 24.8 | +2.8 | 1839.7 | 465.4 | −11.9 |
| Shrubs | 16.7 | 48.2 | +27.4 | 1420.3 | 3552.7 | +18.4 |
| Long grass | 61.1 | 36.7 | −21.2 | 7438.2 | 6715.1 | −6.3 |
| Sparse grass | 10.5 | 4.4 | −5.3 | 389.9 | 421.9 | +0.3 |
| Bare | 3.0 | 0.6 | −2.2 | 169.2 | 220.8 | +0.4 |
| Swamp | 1.5 | 0.0 | −1.3 | 140.7 | 106.5 | −0.3 |
| Built up | 0.0 | 0.0 | 0.0 | 7.8 | 6.4 | 0.0 |
| Water | 0.3 | 0.0 | −0.3 | 152.7 | 69.6 | −0.7 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marston, C.G.; Aplin, P.; Wilkinson, D.M.; Field, R.; O’Regan, H.J. Scrubbing Up: Multi-Scale Investigation of Woody Encroachment in a Southern African Savannah. Remote Sens. 2017, 9, 419. https://doi.org/10.3390/rs9050419
Marston CG, Aplin P, Wilkinson DM, Field R, O’Regan HJ. Scrubbing Up: Multi-Scale Investigation of Woody Encroachment in a Southern African Savannah. Remote Sensing. 2017; 9(5):419. https://doi.org/10.3390/rs9050419
Chicago/Turabian StyleMarston, Christopher G., Paul Aplin, David M. Wilkinson, Richard Field, and Hannah J. O’Regan. 2017. "Scrubbing Up: Multi-Scale Investigation of Woody Encroachment in a Southern African Savannah" Remote Sensing 9, no. 5: 419. https://doi.org/10.3390/rs9050419
APA StyleMarston, C. G., Aplin, P., Wilkinson, D. M., Field, R., & O’Regan, H. J. (2017). Scrubbing Up: Multi-Scale Investigation of Woody Encroachment in a Southern African Savannah. Remote Sensing, 9(5), 419. https://doi.org/10.3390/rs9050419