
Prediction of Species-Specific Volume Using Different Inventory Approaches by Fusing Airborne Laser Scanning and Hyperspectral Data


Abstract
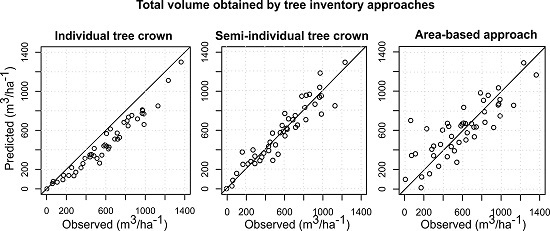
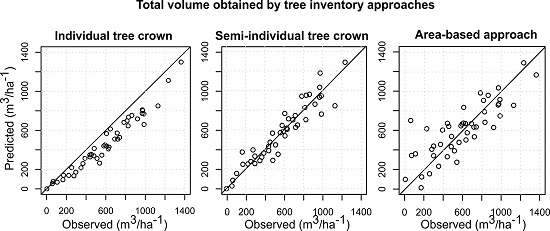{kind=link}
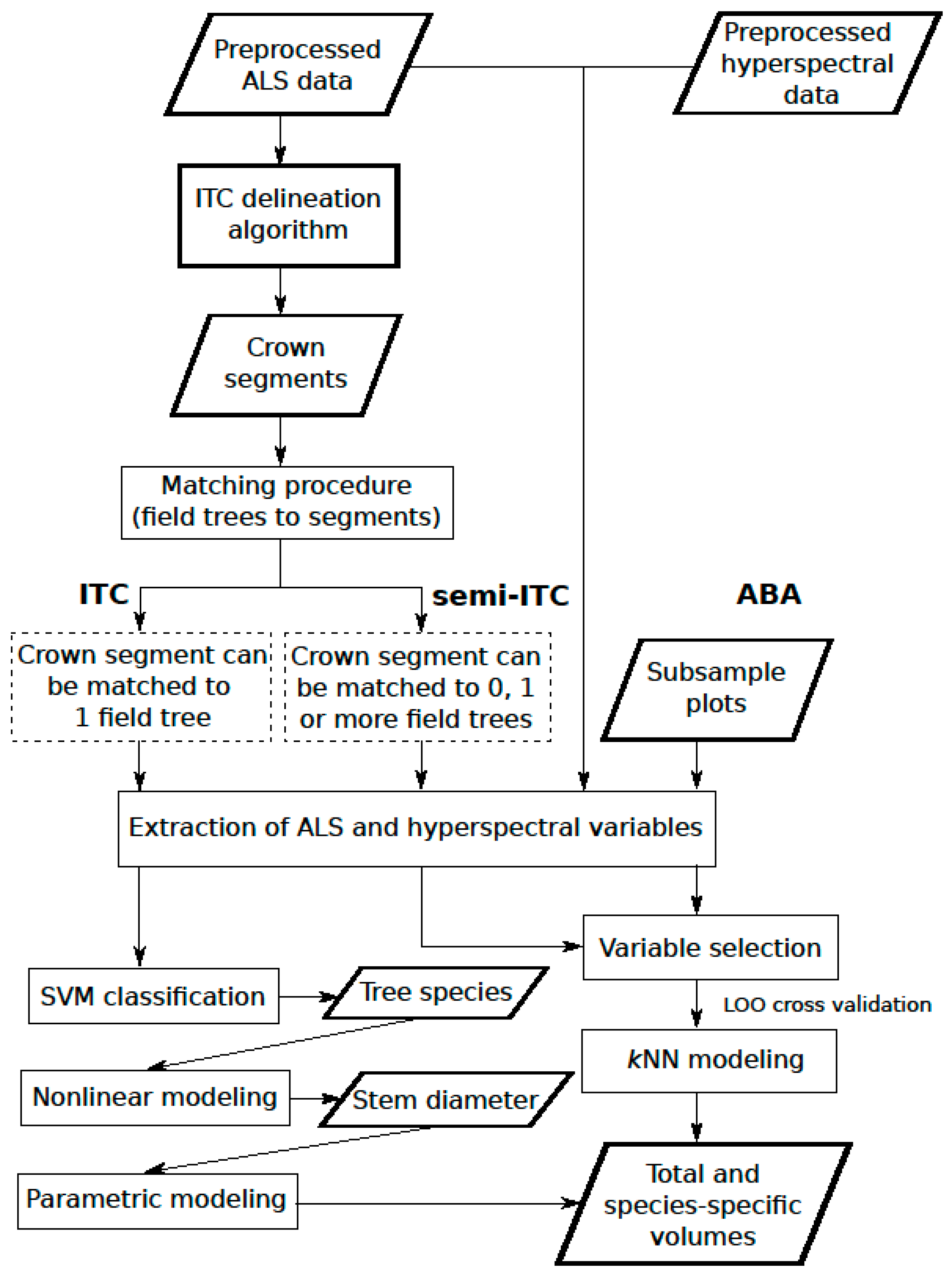{kind=link}
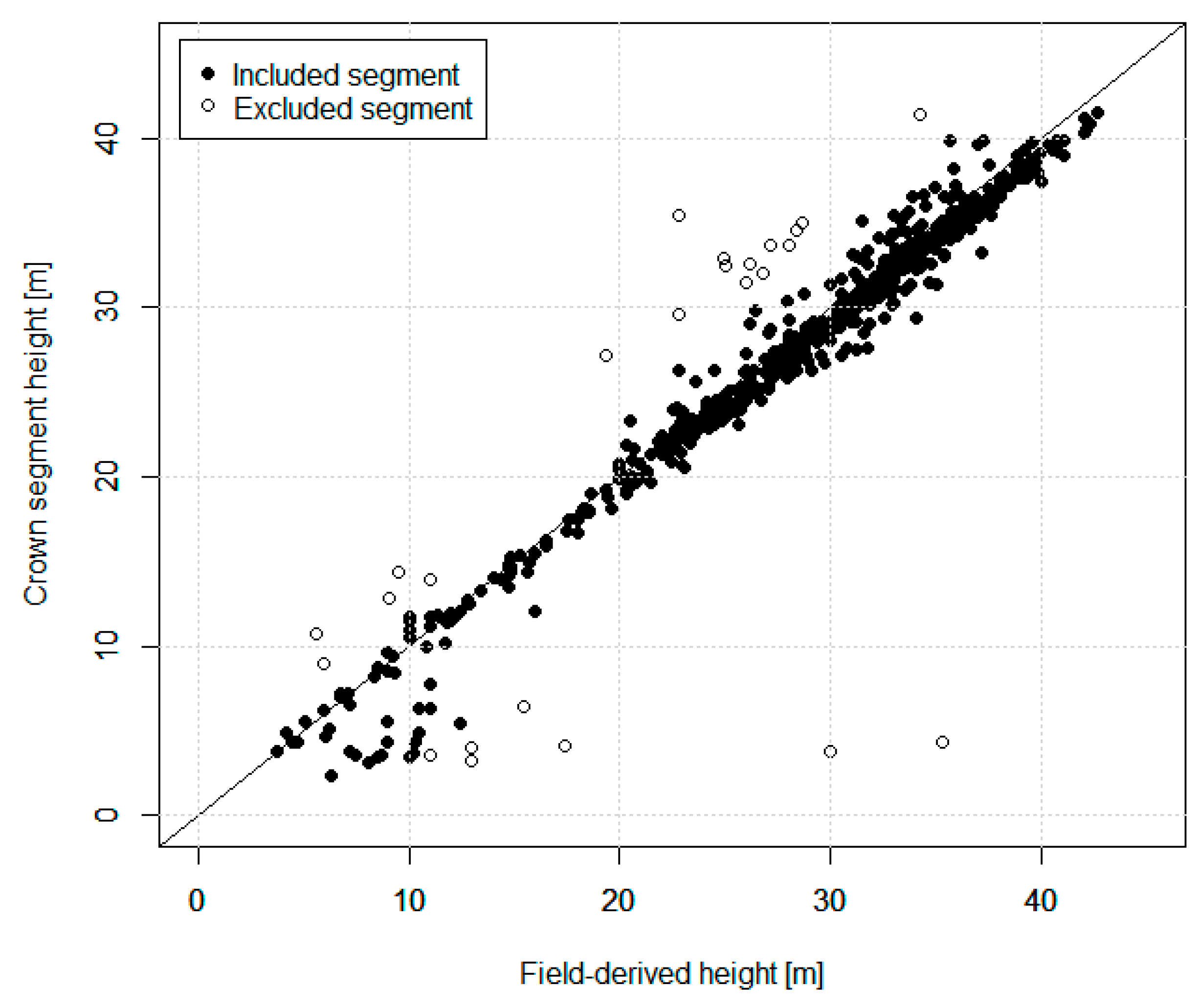{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Kandare, K.; Dalponte, M.; Ørka, H.O.; Frizzera, L.; Næsset, E. Prediction of Species-Specific Volume Using Different Inventory Approaches by Fusing Airborne Laser Scanning and Hyperspectral Data. Remote Sens. 2017, 9, 400. https://doi.org/10.3390/rs9050400
Kandare K, Dalponte M, Ørka HO, Frizzera L, Næsset E. Prediction of Species-Specific Volume Using Different Inventory Approaches by Fusing Airborne Laser Scanning and Hyperspectral Data. Remote Sensing. 2017; 9(5):400. https://doi.org/10.3390/rs9050400
Chicago/Turabian StyleKandare, Kaja, Michele Dalponte, Hans Ole Ørka, Lorenzo Frizzera, and Erik Næsset. 2017. "Prediction of Species-Specific Volume Using Different Inventory Approaches by Fusing Airborne Laser Scanning and Hyperspectral Data" Remote Sensing 9, no. 5: 400. https://doi.org/10.3390/rs9050400
APA StyleKandare, K., Dalponte, M., Ørka, H. O., Frizzera, L., & Næsset, E. (2017). Prediction of Species-Specific Volume Using Different Inventory Approaches by Fusing Airborne Laser Scanning and Hyperspectral Data. Remote Sensing, 9(5), 400. https://doi.org/10.3390/rs9050400