
Fluorescence-Based Approach to Estimate the Chlorophyll-A Concentration of a Phytoplankton Bloom in Ardley Cove (Antarctica)



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Apparent Optical Properties
2.2. In Situ Chlorophyll-a Concentration
2.3. In Vivo Fluorescence
2.4. Satellite/In Situ Match-Ups
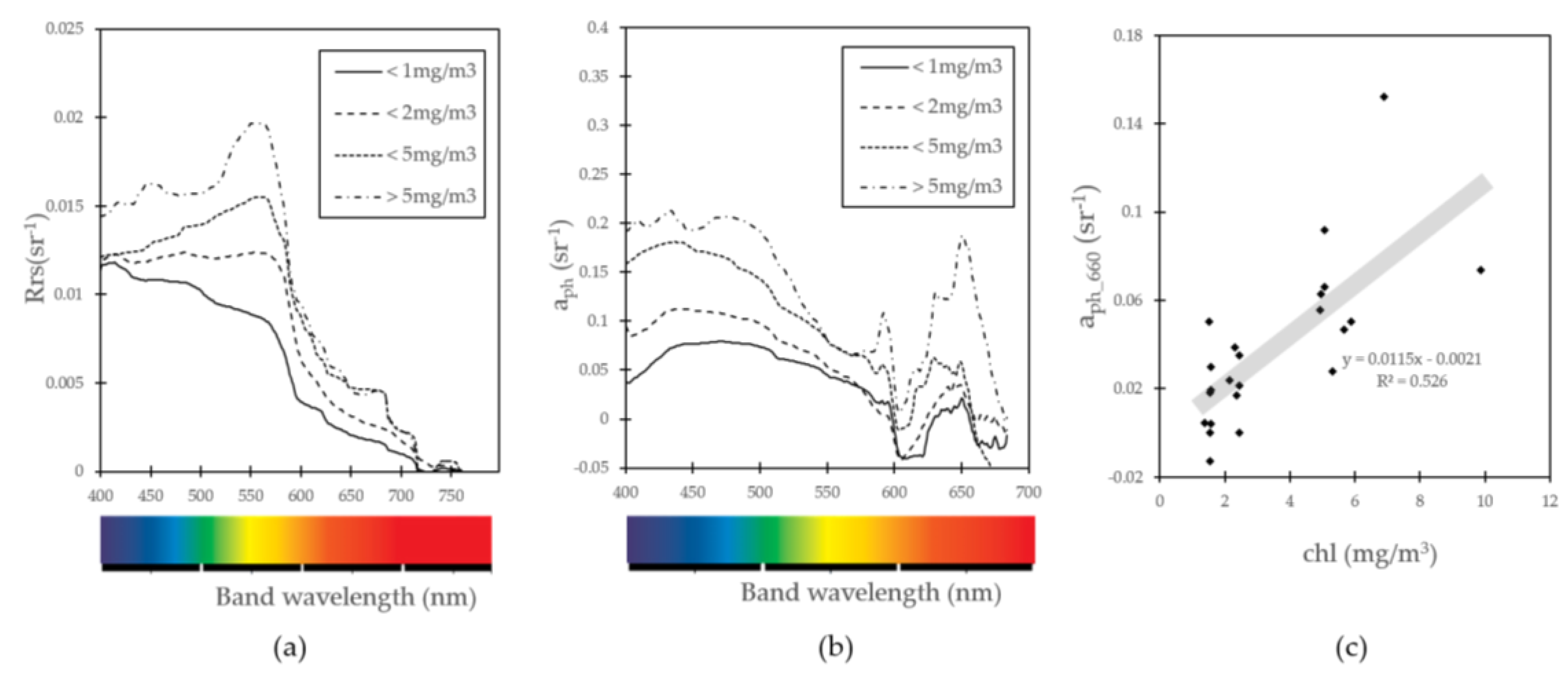
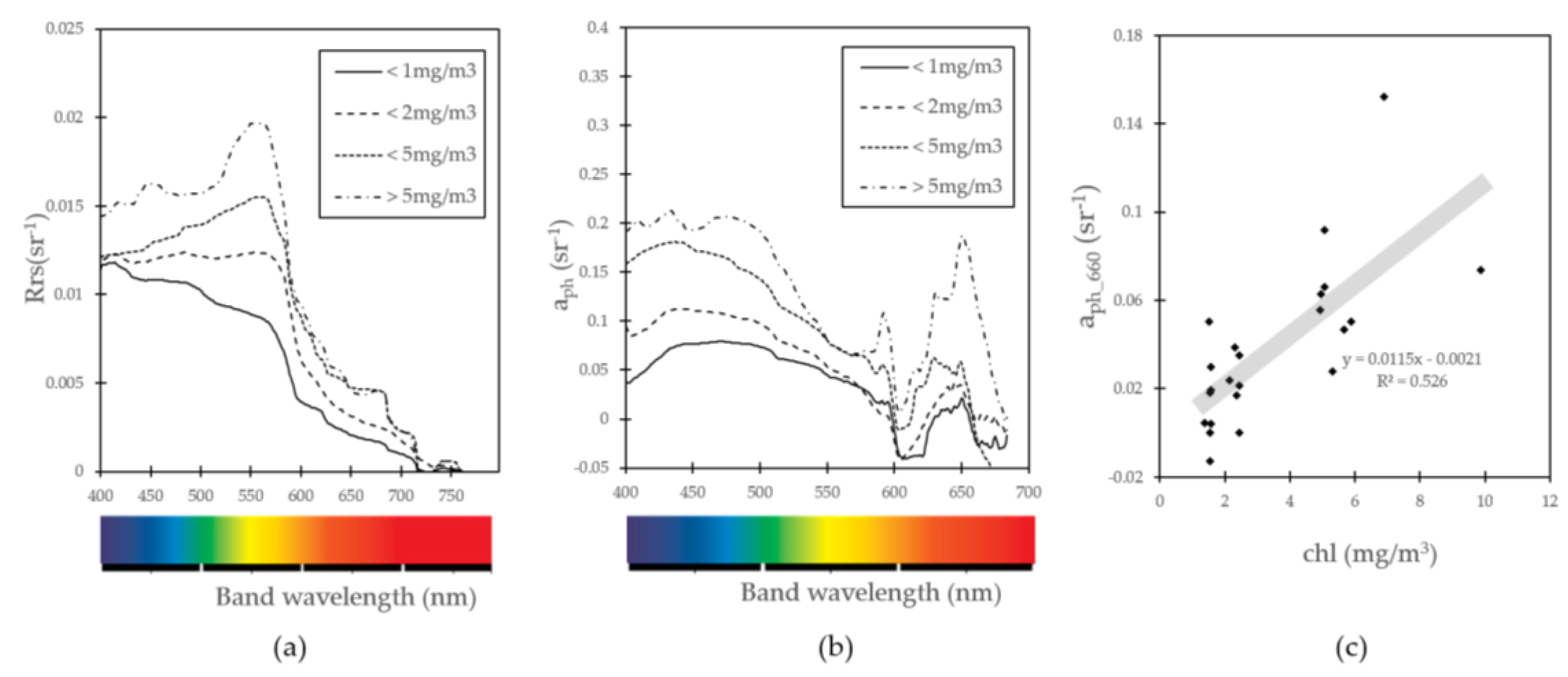
2.5. Phytoplankton Absorption Coefficent Spectrum
3. Results
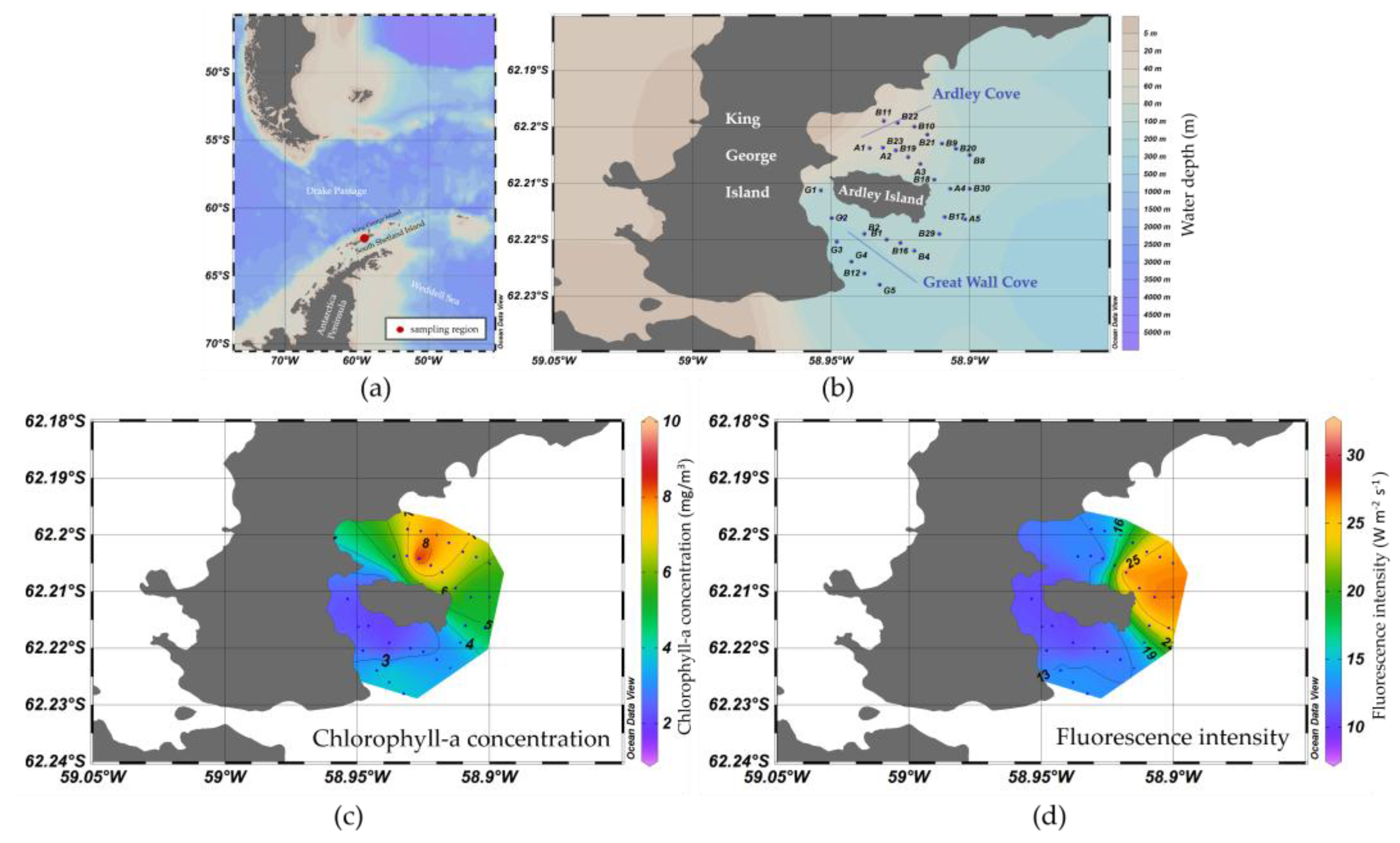
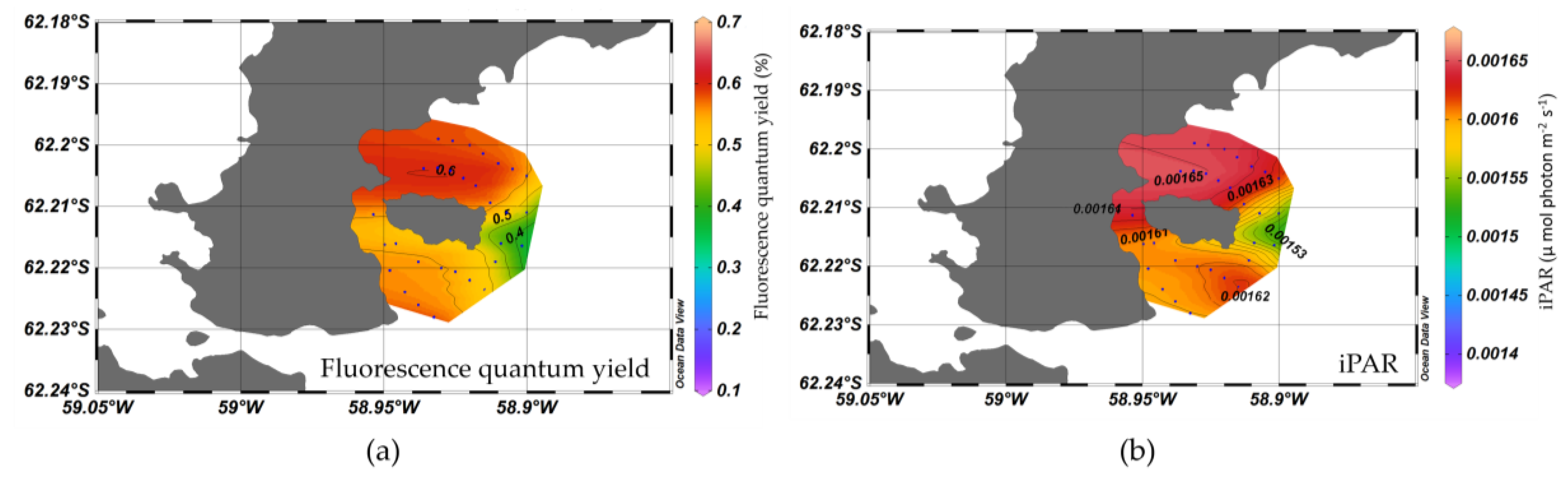
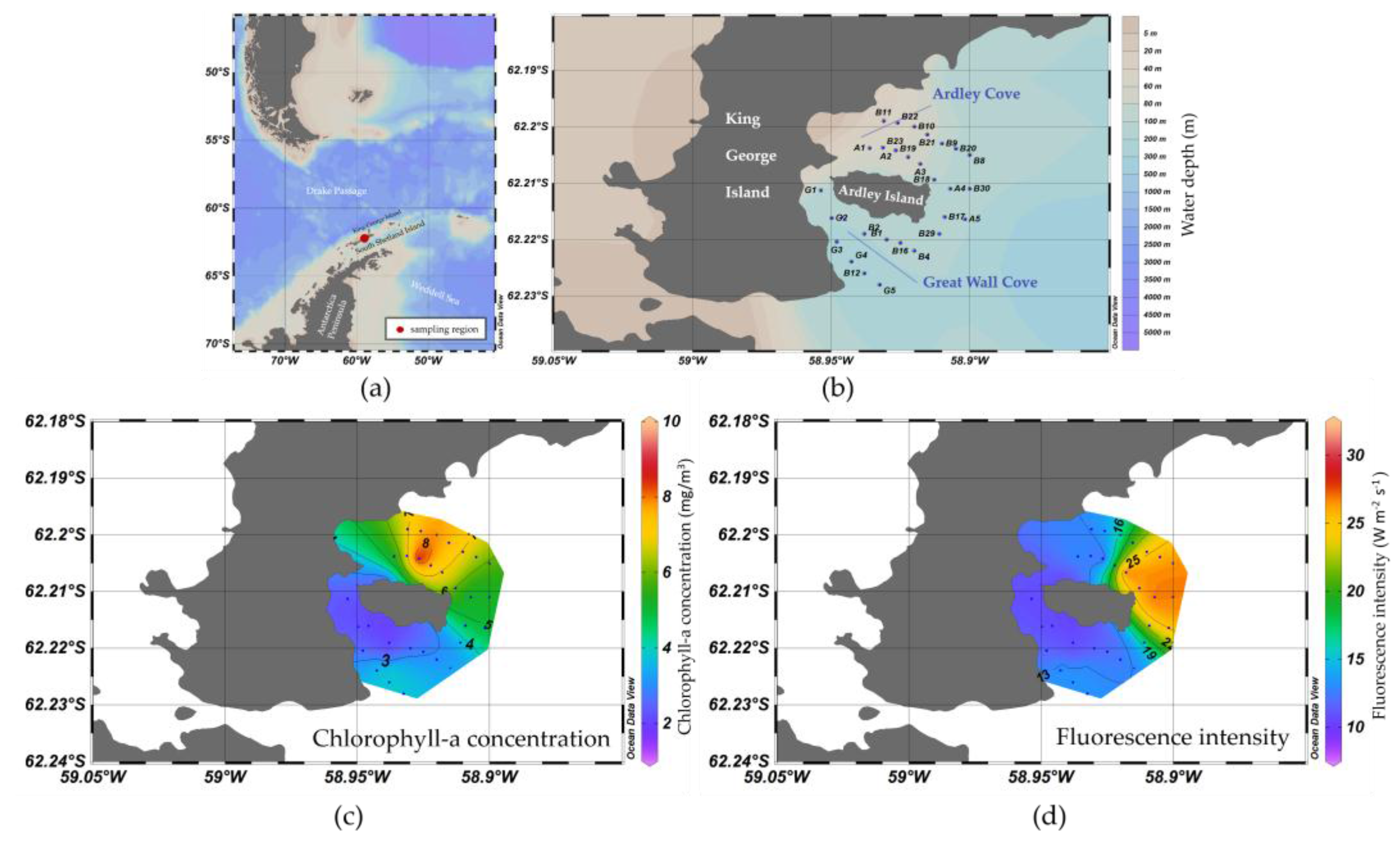
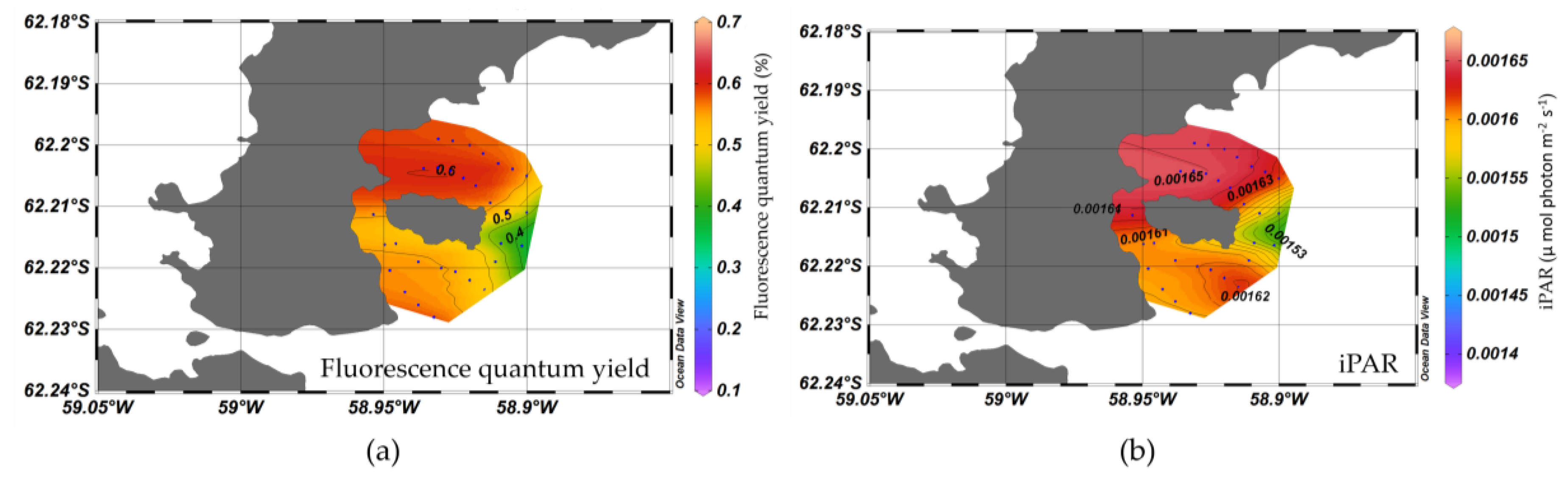
3.1. Spatial Distribution of Water Optical Properties
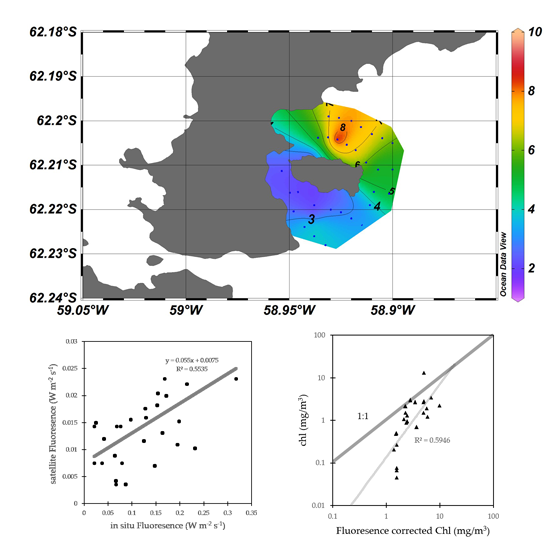
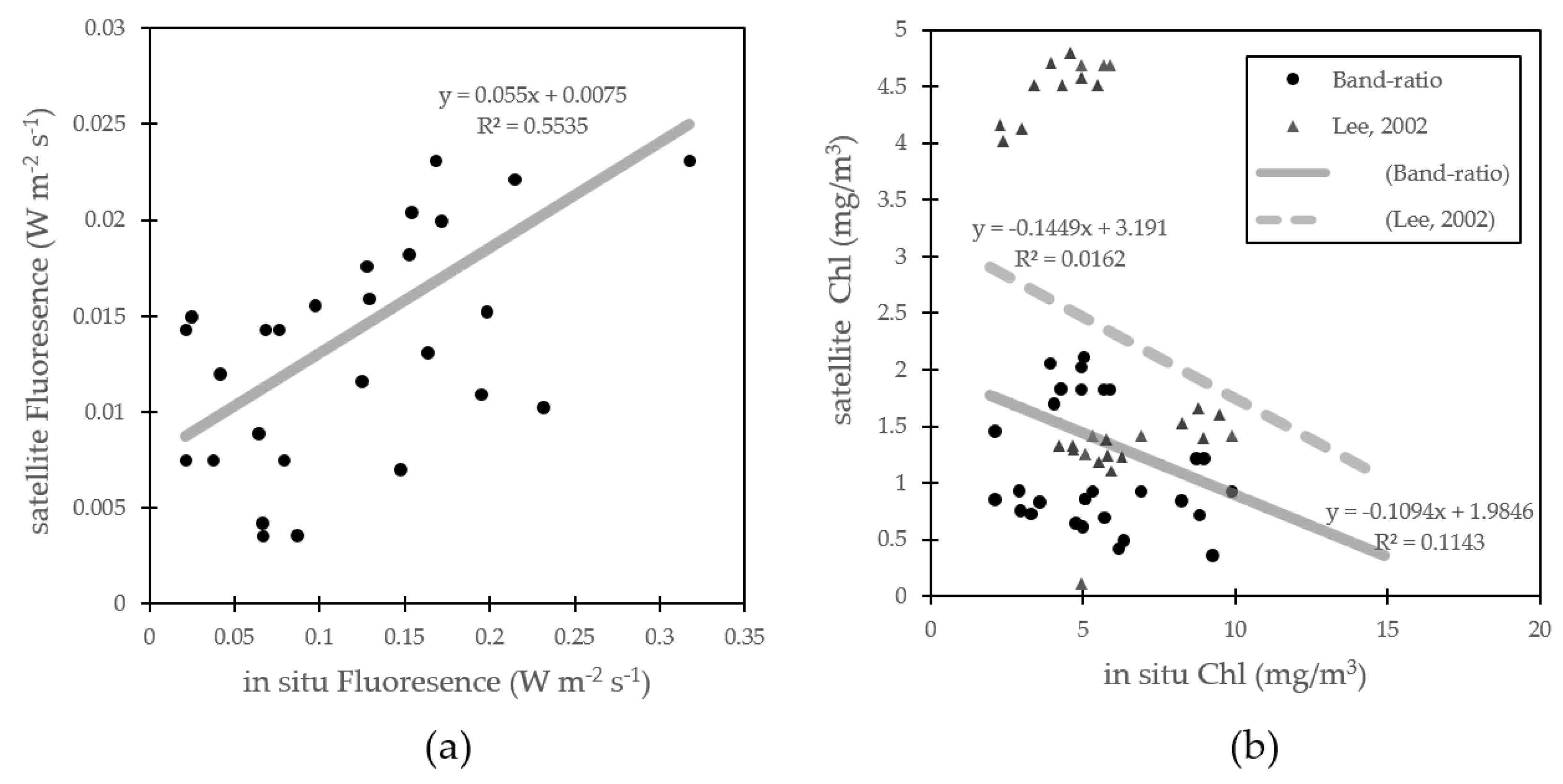
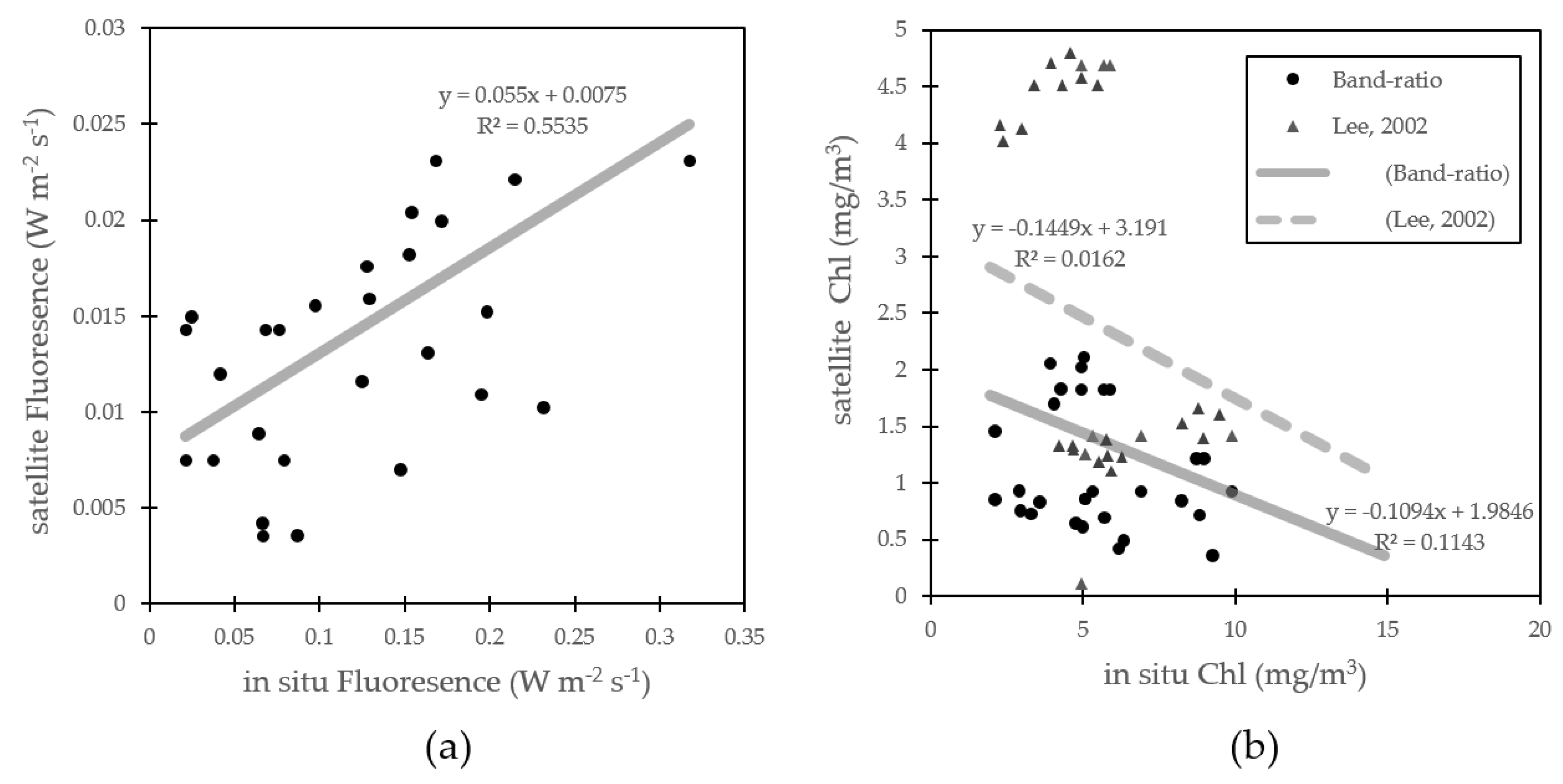
3.2. Algorithim Chlorophyll-a Estimation and In Situ/Satellite Match-Ups
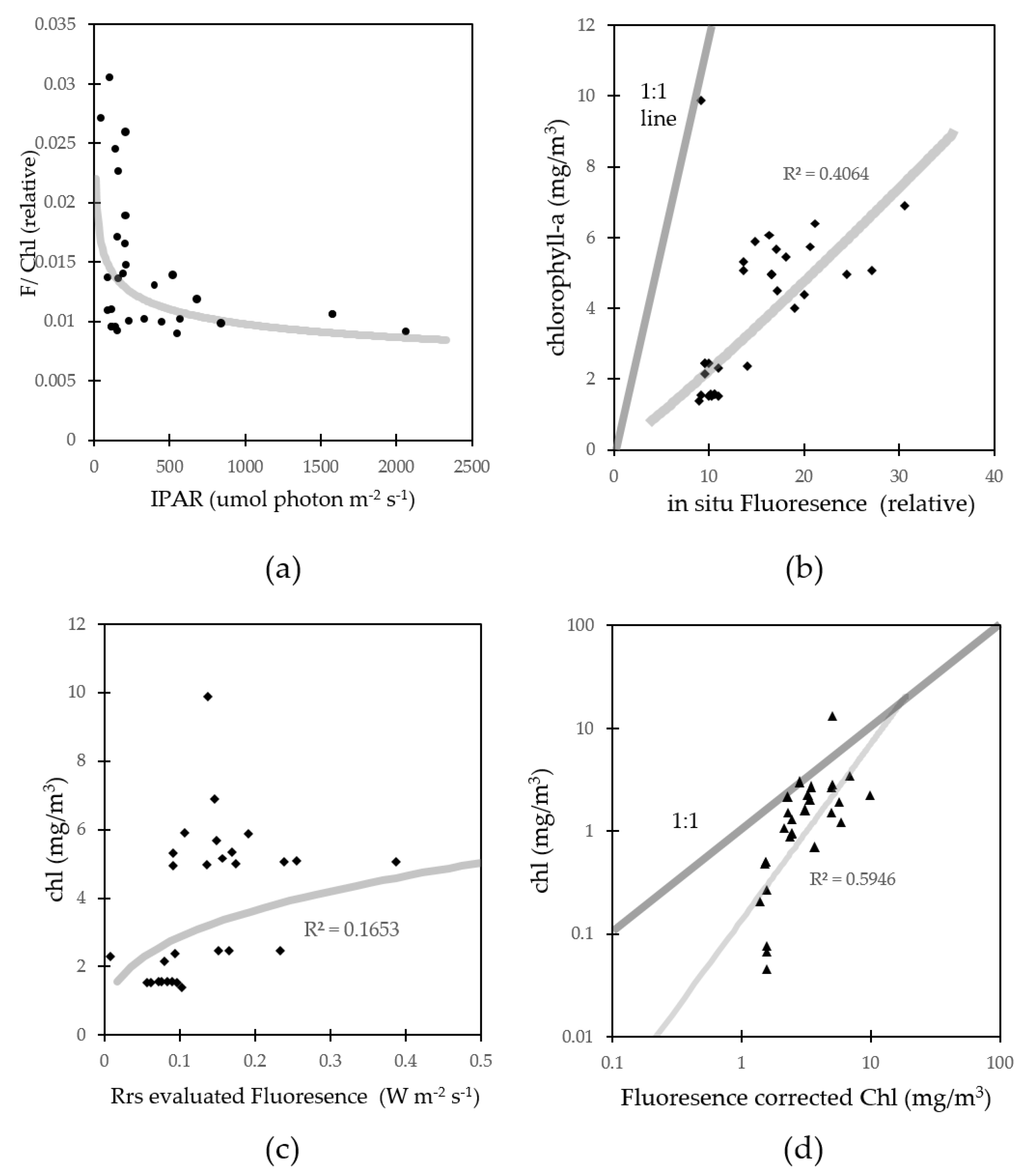
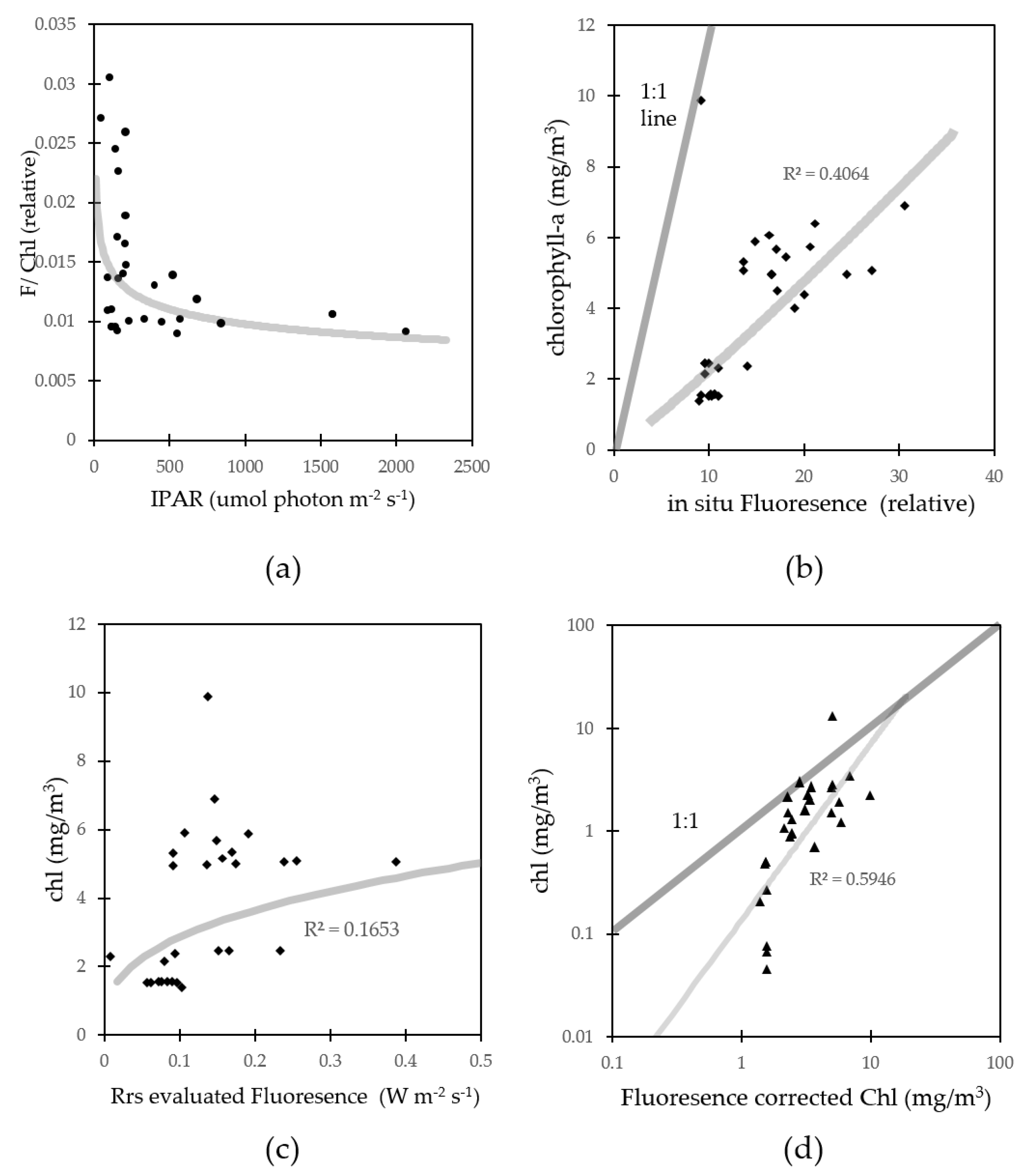
3.3. Fluorescence Approach Estimation for Chlorophyll-a
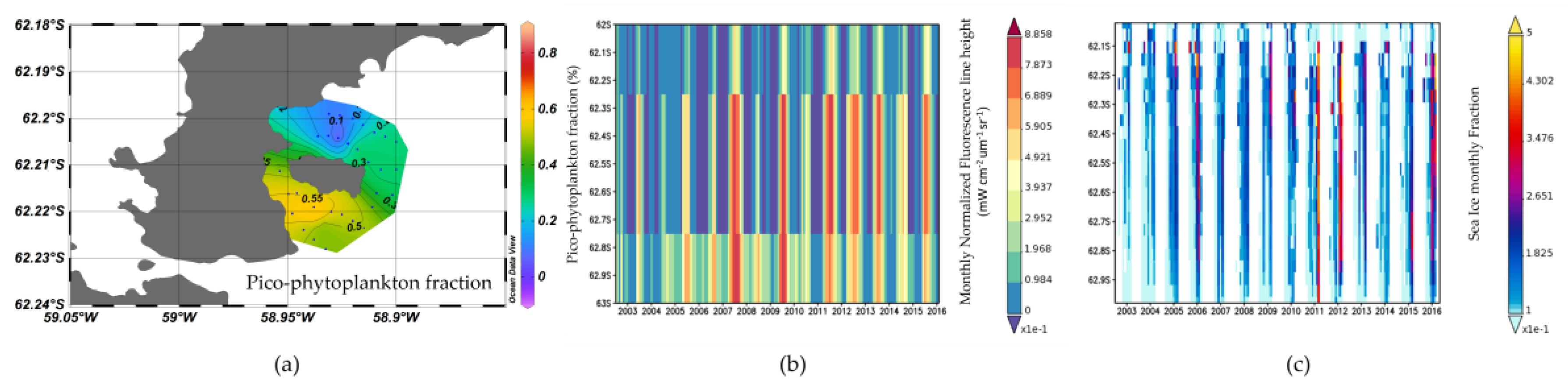
3.4. Nutrient, Light, and Phytoplankton Composition
4. Discussion
4.1. Fluorescence Approach Estimation for Chlorophyll-a
4.2. Factors on Phytoplankton Bloom in KGI
4.3. Errors for Fluorescence Approach Estimating Chlorophyll-a
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol and Description | Equation and Process |
|---|---|
| , below-surface remote-sensing reflectance (sr−1); , Above-surface remote-sensing reflectance (sr−1) | |
| , ratio of backscattering coefficient to the sum of absorption and backscattering coefficients, | , |
| , absorption coefficient of pure water (m−1); 443, 490, 555, 667, band wavelength (nm) | , |
| , backscattering coefficient of pure water (m−1); , Backscattering coefficient of suspended particles (m−1) | |
| , spectral power of particle backscattering coefficient | |
| , all band wavelength (nm) | |
| , absorption coefficient of gelbstoff and detritus (m−1); S, Spectral slope for gelbstoff absorption coefficient | , , |
| , spectral exponential coefficient for gelbstoff and detritus | |
| , absorption coefficient of phytoplankton |
References
- Falkowski, P.G. The power of plankton. Nature 2012. [Google Scholar] [CrossRef] [PubMed]
- Pope, A.; Wagner, P.; Johnson, R.; Baeseman, J.; Newman, L. Community review of Southern Ocean satellite data needs. Antarct. Sci. 2016. [Google Scholar] [CrossRef]
- Babin, M.; Arrigo, K.; Bélanger, S.; Forge, M.-H. Ocean Colour Remote Sensing in Polar Seas; IOCCG Report Series, No. 16; International Ocean Colour Coordinating Group: Dartmouth, NS, Canada, 2015. [Google Scholar]
- Reynolds, R.A.; Stramski, D.; Mitchell, B.G. A chlorophyll-a-dependent semi-analytical reflectance model derived from field measurements of absorption and backscattering coefficients within the Southern Ocean. J. Geophys. Res. 2001, 106, 7125–7138. [Google Scholar] [CrossRef]
- Marrari, M.; Hu, C.; Daly, K. Validation of SeaWiFS chlorophyll-a a concentrations in the Southern Ocean: A revisit. Remote Sens. Environ. 2006, 105, 367–375. [Google Scholar] [CrossRef]
- Moore, J.K.; Abbott, M.R. Phytoplankton chlorophyll-a distributions and primary production in the Southern Ocean. J. Geophys. Res. Ocean. 2000, 105, 28709–28722. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Smith, R.C. Bio-optical properties and remote sensing ocean color algorithms for Antarctic Peninsula waters. J. Geophys. Res. 2000, 105, 26301–26312. [Google Scholar] [CrossRef]
- Gregg, W.W.; Casey, N.W. Global and regional evaluation of the SeaWiFS chlorophyll-a data set. Remote Sens. Environ. 2004, 93, 463–479. [Google Scholar] [CrossRef]
- Kwok, R.; Comiso, J.C. Spatial patterns of variability in Antarctic surface temperature: Connections to the southern hemisphere annular mode and the southern oscillation. Geophys. Res. Lett. 2002. [Google Scholar] [CrossRef]
- Gabric, A.J.; Shephard, J.M.; Knight, J.M.; Jones, G.; Trevena, A.J. Correlations between the satellite-derived seasonal cycles of phytoplankton biomass and aerosol optical depth in the Southern Ocean: Evidence for the influence of sea ice. Global Biogeochem. Cycles. 2005, 19, 1–10. [Google Scholar] [CrossRef]
- Friedrichs, M.A.; Carr, M.E.; Barber, R.T.; Scardi, M.; Antoine, D.; Armstrong, R.A.; Asanuma, I.; Behrenfeld, M.J.; Buitenhuis, E.T.; Chai, F.; et al. Assessing the uncertainties of model estimates of primary productivity in the tropical Pacific Ocean. J. Mar. Syst. 2009, 76, 113–133. [Google Scholar] [CrossRef]
- Zeng, C.; Xu, H.; Fischer, A. M. Chlorophyll-a estimation around the Antarctica peninsula using satellite algorithms: hints from field water leaving reflectance. Sensors 2016. [Google Scholar] [CrossRef] [PubMed]
- Szeto, M.; Werdell, P.J.; Moore, T.S.; Campbell, J.W. Are the world’s oceans optically different? J. Geophys Res. 2011. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L.; Bushinsky, S. Primary production in the Southern Ocean, 1997–2006. J. Geophys. Res. 2008. [Google Scholar] [CrossRef]
- Bélanger, S.; Ehn, J.K.; Babin, M. Impact of sea ice on the retrieval of water-leaving reflectance, chlorophyll-a a concentration and inherent optical properties from satellite ocean color data. Remote Sens. Environ. 2007, 111, 51–68. [Google Scholar] [CrossRef]
- Wang, M.; Shi, W. Detection of ice and mixed ice–water pixels for MODIS ocean color data processing. IEEE Trans. Geosci. Remote Sens. 2009, 47, 2510–2518. [Google Scholar] [CrossRef]
- McFarquhar, G.M.; Wood, R.; Bretherton, C.S.; Alexander, S.; Jakob, C.; Marchand, R.; Protat, A.; Quinn, P.; Siems, S.T.; Weller, R.A. The southern ocean clouds, radiation, aerosol transport experimental study (SOCRATES): An observational campaign for determining role of clouds, aerosolsand radiation in climate system. In Proceedings of the 2014 AGU Fall Meeting, San Francisco, CA, USA, 15–19 December 2014.
- Kay, J.E.; Medeiros, B.; Hwang, Y.T.; Gettelman, A.; Perket, J.; Flanner, M.G. Processes controlling Southern Ocean shortwave climate feedbacks in CESM. Geophys. Res. Lett. 2014, 41, 616–622. [Google Scholar] [CrossRef]
- Maritorena, S.; Siegel, D.A.; Peterson, A. Optimization of a semi-analytical ocean color model for global scale applications. Appl. Opt. 2002, 41, 2705–2714. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.; Carder, K.L.; Arnone, R.A. Deriving inherent optical properties from water color: A multiband quasi-analytical algorithm for optically deep waters. Appl. Opt. 2002, 41, 5755–5772. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Claustre, H.; Blain, S.; D’Ortenzio, F.; Antoine, D.; Ras, J.; Guinet, C. Quenching correction for in vivo chlorophyll-a fluorescence acquired by autonomous platforms: A case study with instrumented elephant seals in the Kerguelen region (Southern Ocean). Limnol. Oceanogr. Method. 2012, 10, 483–495. [Google Scholar] [CrossRef]
- Palmer, S.C.; Hunter, P.D.; Lankester, T.; Hubbard, S.; Spyrakos, E.; Tyler, A.N.; Presing, M.; Horvath, H.; Lamb, A.; Balzter, H.; et al. Validation of Envisat MERIS algorithms for chlorophyll-a retrieval in a large, turbid and optically-complex shallow lake. Remote Sens. Environ. 2015, 157, 158–169. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Westberry, T.K.; Boss, E.; O’Malley, R.T.; Siegel, D.A.; Wiggert, J.D.; Franz, B.A.; McClain, C.R.; Feldman, G.C.; Doney, S.C.; et al. Satellite-detected fluorescence reveals global physiology of ocean phytoplankton. Biogeosciences 2009, 6, 779–794. [Google Scholar] [CrossRef]
- Schloss, I.R.; Abele, D.; Moreau, S.; Demers, S.; Bers, A.V.; González, O.; Ferreyra, G.A. Response of phytoplankton dynamics to 19-year (1991–2009) climate trends in Potter Cove (Antarctica). J. Marine Syst. 2012, 92, 53–66. [Google Scholar] [CrossRef]
- Schloss, I.R.; Wasilowska, A.; Dumont, D.; Almandoz, G.O.; Hernando, M.P.; Michaud-Tremblay, C.A.; Saravia, L.; Rzepecki, M.; Monien, P.; Monien, D.; et al. On the phytoplankton bloom in coastal waters of southern King George Island (Antarctica) in January 2010: An exceptional feature? Limnol. Oceanogr. 2014. [Google Scholar] [CrossRef]
- Mueller, J.L.; Fargion, G.S.; McClain, C.R.; Pegau, S.; Zanefeld, J.R.V.; Mitchell, B.G.; Kahru, M.; Wieland, J.; Stramska, M. Ocean Optics Protocols for Satellite Ocean Color Sensor Validation, Revision 4, Volume Iv: Radiometric Measurements and Data Analysis Protocols; Goddard Space Flight Space Center: Greenbelt, MD, USA, 2003. [Google Scholar]
- Tang, J.W.; Tian, G.L.; Wang, X.Y.; Wang, X.M.; Song, Q.J. The methods of water spectra measurement and analysis i: Above-water method. J. Remote Sens. Beijing 2004, 8, 37–44. [Google Scholar]
- Mobley, C. Overview of Optical Oceanography. Available online: http://www.oceanopticsbook.info/view/overview_of_optical_oceanography/reflectances (accessed on 25 June 2015).
- Kohavi, R. A study of cross-validation and bootstrap for accuracy estimation and model selection. In Proceedings of the 14th International Joint Conference on Artificial Intelligence, Montreal, QB, Canada, 20–25 August 1995.
- Lee, Z.P.; Lubac, B.; Werdell, J.; Arnone, R. An update of the Quasi-Analytical Algorithm (QAA_v5). Available online: http://www.ioccg.org/groups/Software_OCA/QAA_v5.pdf (accessed on 28 July 2009).
- Pope, R.M.; Fry, E.S. Absorption spectrum (380–700 nm) of pure water. II. Integrating cavity measurements. Appl. Opt. 1997, 36, 8710–8723. [Google Scholar] [CrossRef] [PubMed]
- Ciotti, Á.; Lewis, M.R.; Cullen, J.J. Assessment of the relationships between dominant cell size in natural phytoplankton communities and the spectral shape of the absorption coefficient. Limnol. Oceanogr. 2002, 2, 404–417. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Kahru, M. Bio-optical algorithms for ADEOS-2 GLI. J. Remote Sens. Soc. Jpn. 2009, 29, 80–85. [Google Scholar]
- Behrenfeld, M. J.; Halsey, K.; Milligan, A. J. Evolved physiological responses of phytoplankton to their integrated growth environment. Phil. Trans. Royal Soc. B 2008, 363, 2687–2703. [Google Scholar] [CrossRef] [PubMed]
- Babin, M.; Morel, A.; Gentili, B. Remote sensing of sea surface sun-induced chlorophyll-a fluorescence: consequences of natural variations in the optical characteristics of phytoplankton and the quantum yield of chlorophyll-a a fluorescence. Int. J. Remote Sens. 1996, 17, 2417–2448. [Google Scholar] [CrossRef]
- Gower, J. On the use of satellite-measured chlorophyll fluorescence for monitoring coastal waters. Int. J. Remote Sens. 2016, 37, 2077–2086. [Google Scholar] [CrossRef]
- Ortega-Retuerta, E.; Frazer, T.K.; Duarte, C.M.; Ruiz-Halpern, S.; Tovar-Sánchez, A.; Arrieta López de Uralde, J.M.; Reche, I. Biogeneration of chromophoric dissolved organic matter by bacteria and krill in the Southern Ocean. Limnol. Oceanogr. 2009, 54, 1941–1950. [Google Scholar] [CrossRef]
- Bony, S.; Stevens, B.; Frierson, D.M.; Jakob, C.; Kageyama, M.; Pincus, R.; Shepherd, T.G.; Sherwood, S.C.; Siebesma, A.P.; Sobel, A.H.; et al. Clouds, circulation and climate sensitivity. Nat. Geosci. 2015, 8, 261–268. [Google Scholar] [CrossRef]
- Browning, T.J.; Bouman, H.A.; Moore, C.M. Satellite-detected fluorescence: Decoupling nonphotochemical quenching from iron stress signals in the South Atlantic and Southern Ocean. Glob. Biogeochem. Cycles 2014, 28, 510–524. [Google Scholar] [CrossRef]
- Sathyendranath, S.; Platt, T. Spectral effects in bio-optical control on the ocean system. Oceanologia. 2007, 49, 5–39. [Google Scholar]
- Boyd, P.W.; Arrigo, K.R.; Strzepek, R.; Dijken, G.L. Mapping phytoplankton iron utilization: Insights into Southern Ocean supply mechanisms. J. Geophys. Res. Ocean. 2012, 117, 304–315. [Google Scholar] [CrossRef]
- Boyd, P.W.; Watson, A.J.; Law, C.S.; Abraham, E.R.; Trull, T.; Murdoch, R.; Bakker, D.C.; Bowie, A.R.; Buesseler, K.O.; Chang, H.; et al. A mesoscale phytoplankton bloom in the polar Southern Ocean stimulated by iron fertilization. Nature 2000, 407, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Hewes, C.D.; Reiss, C.S.; Holm-Hansen, O. A quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region. Deep-Sea Res. Part I. Oceanogr. Res. Pap. 2009, 56, 1230–1241. [Google Scholar] [CrossRef]
- Tripathy, S.C.; Pavithran, S.; Sabu, P.; Naik, R.K.; Noronha, S.B.; Bhaskar, P.V.; Anilkumar, N. Is phytoplankton productivity in the Indian Ocean sector of Southern? Curr. Sci. 2014, 107, 1019–1026. [Google Scholar]
- Brody, E.; Mitchell, B.G.; Holm-Hansen, O.; Vernet, M. Species-dependent variations of the absorption coefficient in the Gerlache Strait. Antarct. J. USA 1992, 27, 160–162. [Google Scholar]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: cell size and elemental stoichiometry. J. Plankton Res. 2009, 32, 119–137. [Google Scholar] [CrossRef]
- Smith, W.O.; Nelson, D.M. Phytoplankton bloom produced by a receding ice edge in the Ross Sea: Spatial coherence with the density field. Science 1985, 227, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Boetius, A.; Albrecht, S.; Bakker, K.; Bienhold, C.; Felden, J.; Fernández-Méndez, M.; Hendricks, S.; Katlein, C.; Lalande, C.; Krumpen, T.; et al. Export of algal biomass from the melting Arctic sea ice. Science 2013, 339, 1430–1432. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, K.R.; Lowry, K.E.; van Dijken, G.L. Annual changes in sea ice and phytoplankton in polynyas of the Amundsen Sea, Antarctica. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2012, 71, 5–15. [Google Scholar] [CrossRef]
- Gordon, H.R. Removal of atmospheric effects from the satellite imagery of the oceans. Appl. Opt. 1978, 17, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Behrangi, A.; Stephens, G.; Adler, R.F.; Huffman, G.J.; Lambrigtsen, B.; Lebsock, M. An update on the oceanic precipitation rate and its zonal distribution in light of advanced observations from space. J. Clim. 2014, 27, 3957–3965. [Google Scholar] [CrossRef]
- Haynes, J.M.; L’Ecuyer, T.S.; Stephens, G.L.; Miller, S.D.; Mitrescu, C.; Wood, N.B.; Tanelli, S. Rainfall retrieval over the ocean with spaceborne W-band radar. J. Geophys. Res. 2009. [Google Scholar] [CrossRef]
- Meskhidze, N.; Nenes, A. Phytoplankton and cloudiness in the Southern Ocean. Science 2006, 314, 1419–1423. [Google Scholar] [CrossRef] [PubMed]
- Vermote, E.F.; Tanré, D.; Deuzé, J.L.; Herman, M.; Morcrette, J.J.; Kotchenova, S.Y. Second Simulation of A Satellite Signal in the Solar Spectrum-Vector (6SV), 6S User Guide Version 3. Available online: http://6s.ltdri.org/files/tutorial/6S_Manual_Part_1.pdf (accessed on 23 September 2016).
- Marchand, R.; Ackerman, T.; Smyth, M.; Rossow, W.B. A review of cloud top height and optical depth histograms from MISR, ISCCP, and MODIS. J. Geophys. Res. 2010. [Google Scholar] [CrossRef]
- Soppa, M.A.; Dinter, T.; Taylor, B.B.; Bracher, A. Satellite derived euphotic depth in the Southern Ocean: Implications for primary production modelling. Remote Sens. Environ. 2013, 137, 198–211. [Google Scholar] [CrossRef] [Green Version]
- Lee, Z.; Weidemann, A.; Kindle, J.; Arnone, R.; Carder, K.L.; Davis, C. Euphotic zone depth: Its derivation and implication to ocean-color remote sensing. J. Geophys. Res. 2007. [Google Scholar] [CrossRef]
- Schlitzer, R. Carbon export fluxes in the Southern Ocean: Results from inverse modeling and comparison with satellite-based estimates. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 1623–1644. [Google Scholar] [CrossRef]
- Uitz, J.; Claustre, H.; Griffiths, F.B.; Ras, J.; Garcia, N.; Sandroni, V. A phytoplankton class-specific primary production model applied to the Kerguelen Islands region (Southern Ocean). Deep-Sea Res. Part I. Oceanogr. Res. Pap. 2009, 56, 541–560. [Google Scholar] [CrossRef]
- Sullivan, C.W.; McClain, C.R.; Comiso, J.C.; Smith, W.O. Phytoplankton standing crops within an Antarctic ice edge assessed by satellite remote sensing. J. Geophys. Res. Ocean. 1988, 93, 12487–12498. [Google Scholar] [CrossRef]
| Model Algorithms | Relationship (R) with In situ Chlorophyll-a | Relative Error (%) | Regions | References |
|---|---|---|---|---|
| chl = 10(0.573 − 2.259X + 0.203X2 − 1.300X3) + 0.386; X = log((nlw 443 > nlw 460 > 520)/ nlw 545) | 0.269214 | 0.367748 | SO | Mitchell et.al. [33] |
| chl = 2.22X; chl < 1.5 mg/m3; X = log(nlw 440/ nlw 555) | 0.191404 | 0.990063 | WAP | Dierssen et.al. [7] |
| chl = 10(0.78 − 2.52X); chl > 1.5 mg/m3; X = log(nlw 520/ nlw 555) | 0.260399 | 0.632074 | WAP | Dierssen et.al. [7] |
| chl = 0.45 + 0.53X; chl > 1.5 mg/m3; X = log(nlw 520/ nlw 555) | 0.261184 | 1.160298 | WAP | Dierssen et.al. [7] |
| chl = 10 (0.641 − 2.058X − 0.442X2 − 1.140X3); X = log(rrs490/rrs555) | 0.28514 | 0.733532 | WAP | Dierssen et.al. [7] |
| chl = 10(0.3914 + 1.0176X − 0.3114X2 + 0.0186X3 + 0.0610X4); X = log(rrs490/rrs555) | 0.28981 | 0.811026 | WAP | Dierssen et.al. [7] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, C.; Zeng, T.; Fischer, A.M.; Xu, H. Fluorescence-Based Approach to Estimate the Chlorophyll-A Concentration of a Phytoplankton Bloom in Ardley Cove (Antarctica). Remote Sens. 2017, 9, 210. https://doi.org/10.3390/rs9030210
Zeng C, Zeng T, Fischer AM, Xu H. Fluorescence-Based Approach to Estimate the Chlorophyll-A Concentration of a Phytoplankton Bloom in Ardley Cove (Antarctica). Remote Sensing. 2017; 9(3):210. https://doi.org/10.3390/rs9030210
Chicago/Turabian StyleZeng, Chen, Tao Zeng, Andrew M. Fischer, and Huiping Xu. 2017. "Fluorescence-Based Approach to Estimate the Chlorophyll-A Concentration of a Phytoplankton Bloom in Ardley Cove (Antarctica)" Remote Sensing 9, no. 3: 210. https://doi.org/10.3390/rs9030210
APA StyleZeng, C., Zeng, T., Fischer, A. M., & Xu, H. (2017). Fluorescence-Based Approach to Estimate the Chlorophyll-A Concentration of a Phytoplankton Bloom in Ardley Cove (Antarctica). Remote Sensing, 9(3), 210. https://doi.org/10.3390/rs9030210