Robinia pseudoacacia L. Flower Analyzed by Using An Unmanned Aerial Vehicle (UAV)


 and
and
Abstract
1. Introduction
- (1)
- What is the best flying altitude (FA) for the UAVs?
- (2)
- How much flower surface (2D) and volume (3D) can be detected by a UAV?
- (3)
- How many flowers are present per hectare, and per tree?
- (1)
- What mass of nectar and honey (kg) can be produced by one hectare of eight-year-old R. pseudoacacia?
- (2)
- What population of honeybees can survive for one year from the flower production of a one-hectare black locust plantation?
2. Materials and Methods
2.1. Study Area
2.2. Flower Analysis
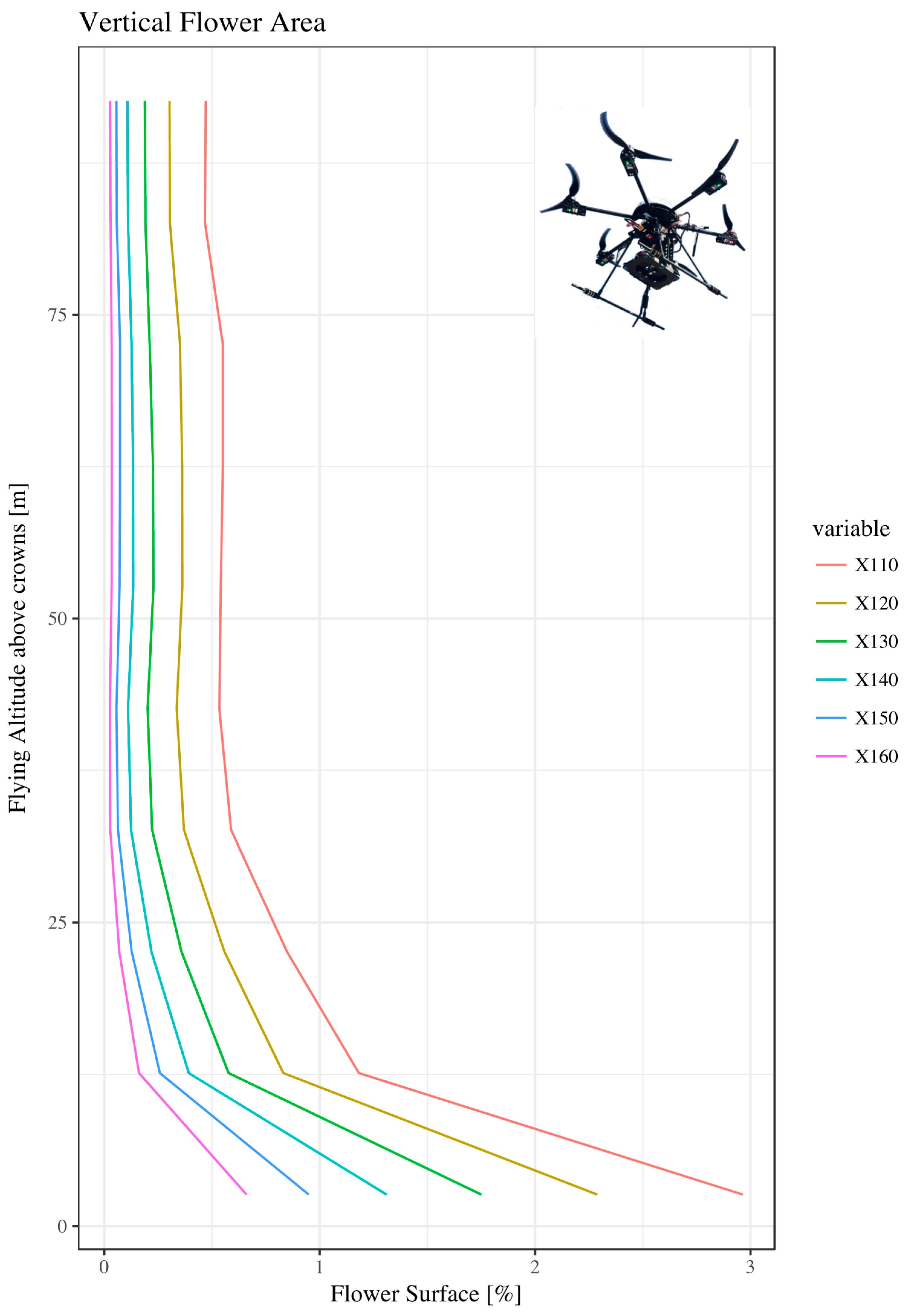
2.2.1. Vertical Analysis
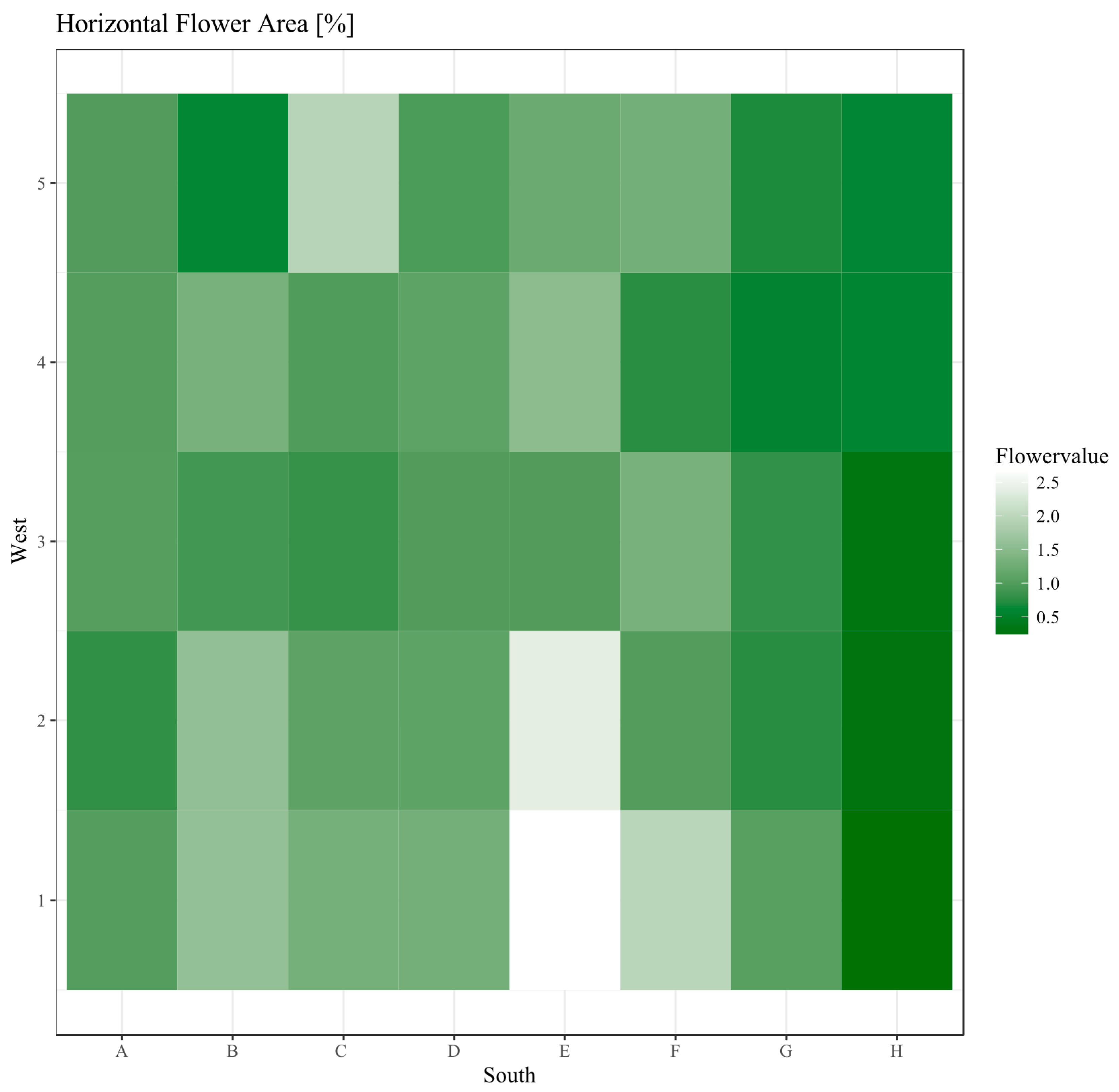
2.2.2. Horizontal Analysis
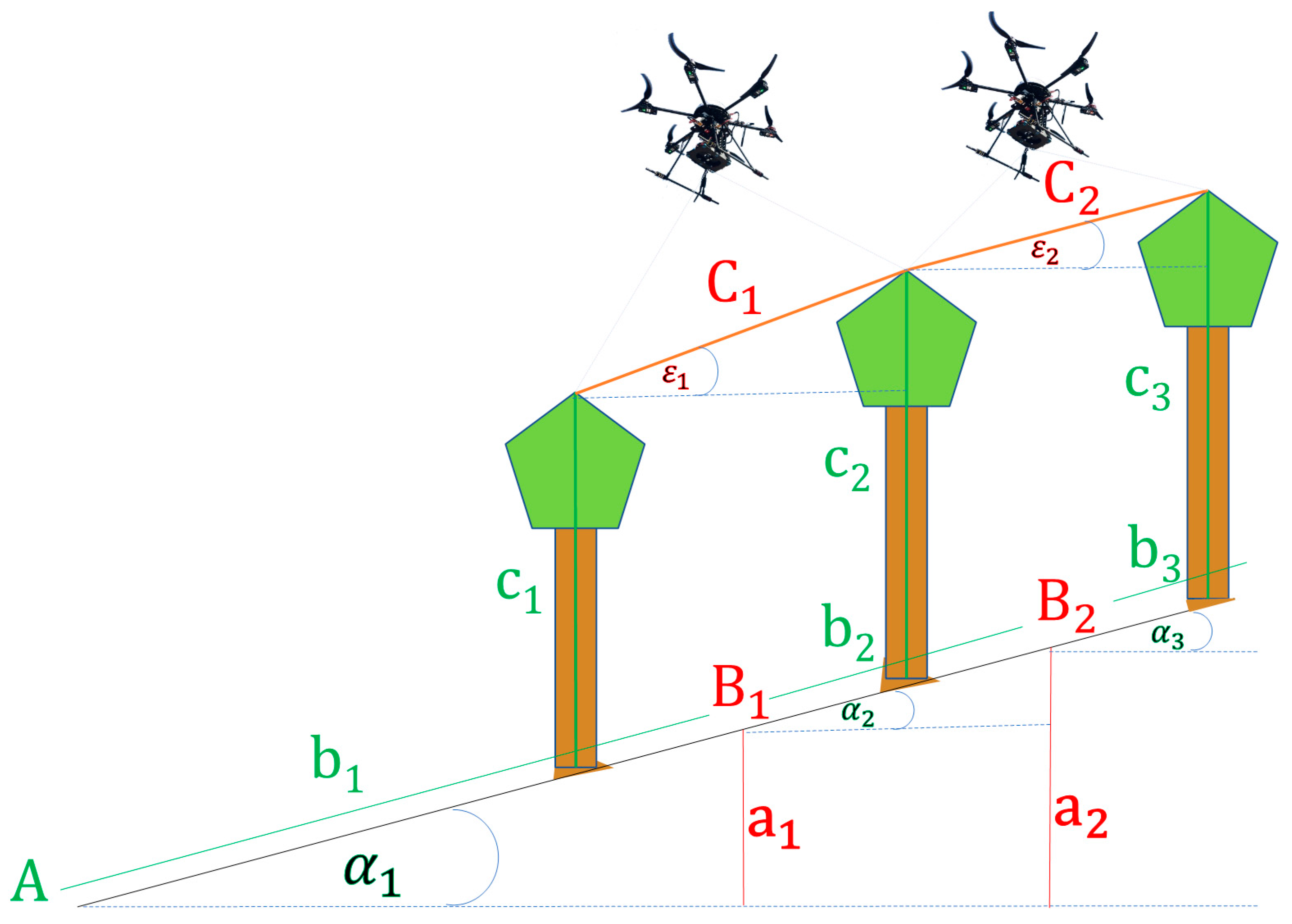
2.2.3. Flower Trees (UAV and Field Data Collection)
2.2.4. Image Analysis
2.2.5. Models: Calculating Flower Number
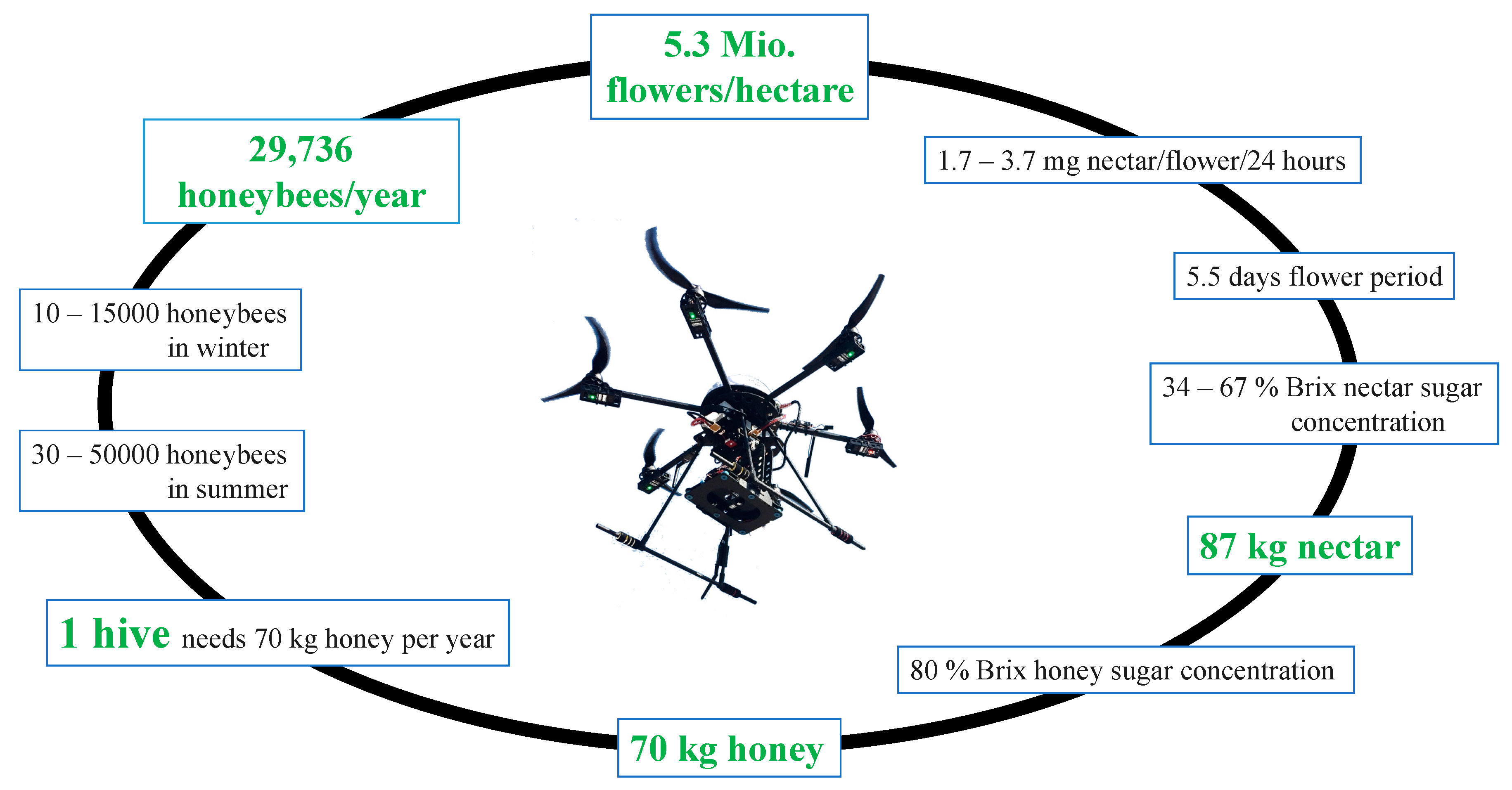
2.2.6. Nectar, Honey, and Honeybees
3. Results
3.1. Vertical Analysis
3.2. Horizontal Analysis
3.3. Flower Trees, Flower Surface, and Volume (2- and 3-Dimensional)
3.4. Flowers, Honey, Apies Mellifera
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 2D | Two-Dimensional, flower surface area |
| 3D | Three-Dimensional, flower volume |
| FA | Flying Altitude |
| FoA | Flower Area |
| FOV | Field of View |
| FSU | Average flower Surface detected by UAV |
| GSD | Ground Sample Distance |
| LAI | Leaf Area Index |
| RGB | Red-Green-Blue |
| UAV | Unmanned Aerial Vehicles |
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Threshold | Overall Accuracy (%) | Producers Accuracy (Flower) | Producers Accuracy (Biomass) | Users Accuracy (Flower) | Users Accuracy (Biomass) | Mean Accuracy (%) | Kappa Coefficient |
|---|---|---|---|---|---|---|---|
| >110 | 98.6 | 100 | 98.5 | 66.2 | 100 | 90.8 | 0.789 |
| >120 | 99.1 | 99.4 | 99.1 | 75.5 | 99.9 | 93.4 | 0.854 |
| >130 | 99.4 | 97.0 | 99.5 | 84.3 | 99.9 | 95.7 | 0.899 |
| >140 | 99.5 | 89.2 | 99.8 | 94.0 | 99.7 | 98.2 | 0.913 |
| >150 | 99.2 | 70.7 | 99.9 | 99.2 | 99.1 | 99.2 | 0.822 |
| >160 | 98.7 | 53.4 | 100 | 100 | 98.7 | 99.0 | 0.690 |
References
- European Commission under the Sixth Framework Programme through the DAISIE Project. Delivering Alien Invasive Species Inventories for Europe—Species Factsheet Robinia Pseudoacacia. Available online: http://www.europe-aliens.org/speciesFactsheet.do?speciesId=11942 (accessed on 29 June 2017).
- Roloff, A.; Weisgerber, H.; Lang, U.; Stimm, B. Bäume Nordamerikas—Von Alligator-Wachholder bis Zuckerahorn. Alle Charakteristischen Arten im Porträt; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2010; ISBN 978-3-527-32825-3. [Google Scholar]
- Rédei, K. Black Locust (Robinia pseudoacacia L.) Growing in Hungary; Hungarian Forest Research Institute: Sarvar, Hungary, 2013; pp. 72–73. [Google Scholar]
- Deutscher Imkerverbund e.V. Honigsorten-Bezeichnungen. Available online: http://deutscherimkerbund.de/userfiles/downloads/satzung_richtlinien/Merkblatt_Sorten_3_4_neu.pdf (accessed on 19 July 2017).
- Rogan, J.; Franklin, J.; Roberts, D.A. A comparison of methods for monitoring multitemporal vegetation change using Thematic Mapper imagery. Remote Sens. Environ. 2002, 80, 143–156. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Knipling, E.B. Physical and physiological basis for the reflectance of visible and near-infrared radiation from vegetation. Remote Sens. Environ. 1970, 1, 155–159. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel algorithms for remote estimation of vegetation fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, S.; Li, D.; Wang, C.; Yang, J. Papaya Tree Detection with UAV Images Using a GPU-Accelerated Scale-Space Filtering Method. Remote Sens. 2017, 9, 721. [Google Scholar] [CrossRef]
- Asner, G.P. Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Baret, F.; Clevers, J.G.P.W.; Steven, M.D. The robustness of canopy gap fraction estimates from red and near-infrared reflectances: A comparison of approaches. Remote Sens. Environ. 1995, 54, 141–151. [Google Scholar] [CrossRef]
- Le Maire, G.; François, C.; Soudani, K.; Berveiller, D.; Pontailler, J.Y.; Bréda, N.; Genet, H.; Davi, H.; Dufrêne, E. Calibration and validation of hyperspectral indices for the estimation of broadleaved forest leaf chlorophyll content, leaf mass per area, leaf area index and leaf canopy biomass. Remote Sens. Environ. 2008, 112, 3846–3864. [Google Scholar] [CrossRef]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Hu, J.; Su, Y.; Tan, B.; Huang, D.; Yang, W.; Schull, M.; Bull, M.A.; Martonchik, J.V.; Diner, D.J.; Knyazikhin, Y.; et al. Analysis of the MISR LAI/FPAR product for spatial and temporal coverage, accuracy and consistency. Remote Sens. Environ. 2007, 107, 334–347. [Google Scholar] [CrossRef]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between leaf area index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]
- North, P.R. Estimation of f APAR, LAI, and vegetation fractional cover from ATSR-2 imagery. Remote Sens. Environ. 2002, 80, 114–121. [Google Scholar] [CrossRef]
- Demarez, V.; Gastellu-Etchegorry, J.P. A modeling approach for studying forest chlorophyll content. Remote Sens. Environ. 2000, 71, 226–238. [Google Scholar] [CrossRef]
- Houborg, R.; Boegh, E. Mapping leaf chlorophyll and leaf area index using inverse and forward canopy reflectance modeling and SPOT reflectance data. Remote Sens. Environ. 2008, 112, 186–202. [Google Scholar] [CrossRef]
- Healey, S.P.; Cohen, W.B.; Zhiqiang, Y.; Krankina, O.N. Comparison of Tasseled Cap-based Landsat data structures for use in forest disturbance detection. Remote Sens. Environ. 2005, 97, 301–310. [Google Scholar] [CrossRef]
- Hilker, T.; Wulder, M.A.; Coops, N.C.; Linke, J.; McDermid, G.; Masek, J.G.; Gao, F.; White, J.C. A new data fusion model for high spatial-and temporal-resolution mapping of forest disturbance based on Landsat and MODIS. Remote Sens. Environ. 2009, 113, 1613–1627. [Google Scholar] [CrossRef]
- Meddens, A.J.; Hicke, J.A.; Vierling, L.A. Evaluating the potential of multispectral imagery to map multiple stages of tree mortality. Remote Sens. Environ. 2011, 115, 1632–1642. [Google Scholar] [CrossRef]
- Bright, B.C.; Hicke, J.A.; Hudak, A.T. Estimating aboveground carbon stocks of a forest affected by mountain pine beetle in Idaho using lidar and multispectral imagery. Remote Sens. Environ. 2012, 124, 270–281. [Google Scholar] [CrossRef]
- Cheng, T.; Rivard, B.; Sánchez-Azofeifa, G.A.; Feng, J.; Calvo-Polanco, M. Continuous wavelet analysis for the detection of green attack damage due to mountain pine beetle infestation. Remote Sens. Environ. 2010, 114, 899–910. [Google Scholar] [CrossRef]
- Coops, N.C.; Johnson, M.; Wulder, M.A.; White, J.C. Assessment of QuickBird high spatial resolution imagery to detect red attack damage due to mountain pine beetle infestation. Remote Sens. Environ. 2006, 103, 67–80. [Google Scholar] [CrossRef]
- Dennison, P.E.; Brunelle, A.R.; Carter, V.A. Assessing canopy mortality during a mountain pine beetle outbreak using GeoEye-1 high spatial resolution satellite data. Remote Sens. Environ. 2010, 114, 2431–2435. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N.; Ramsey, R.D. Combining dendrochronological data and the disturbance index to assess Engelmann spruce mortality caused by a spruce beetle outbreak in southern Utah, USA. Remote Sens. Environ. 2011, 115, 2342–2349. [Google Scholar] [CrossRef]
- Meddens, A.J.; Hicke, J.A.; Vierling, L.A.; Hudak, A.T. Evaluating methods to detect bark beetle-caused tree mortality using single-date and multi-date Landsat imagery. Remote Sens. Environ. 2013, 132, 49–58. [Google Scholar] [CrossRef]
- Meigs, G.W.; Kennedy, R.E.; Cohen, W.B. A Landsat time series approach to characterize bark beetle and defoliator impacts on tree mortality and surface fuels in conifer forests. Remote Sens. Environ. 2011, 115, 3707–3718. [Google Scholar] [CrossRef]
- Skakun, R.S.; Wulder, M.A.; Franklin, S.E. Sensitivity of the thematic mapper enhanced wetness difference index to detect mountain pine beetle red-attack damage. Remote Sens. Environ. 2003, 86, 433–443. [Google Scholar] [CrossRef]
- White, J.C.; Wulder, M.A.; Brooks, D.; Reich, R.; Wheate, R.D. Detection of red attack stage mountain pine beetle infestation with high spatial resolution satellite imagery. Remote Sens. Environ. 2005, 96, 340–351. [Google Scholar] [CrossRef]
- Landmann, T.; Piiroinen, R.; Makori, D.M.; Abdel-Rahman, E.M.; Makau, S.; Pellikka, P.; Raina, S.K. Application of hyperspectral remote sensing for flower mapping in African savannas. Remote Sens. Environ. 2015, 166, 50–60. [Google Scholar] [CrossRef]
- Abdel-Rahman, E.M.; Makori, D.M.; Landmann, T.; Piiroinen, R.; Gasim, S.; Pellikka, P.; Raina, S.K. The utility of AISA eagle hyperspectral data and random forest classifier for flower mapping. Remote Sens. 2015, 7, 13298–13318. [Google Scholar] [CrossRef]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-surface remote sensing of spatial and temporal variation in canopy phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Walter, A.; Finger, R.; Huber, R.; Buchmann, N. Opinion: Smart farming is the key to developing sustainable agriculture. Proc. Natl. Acad. Sci. USA 2017, 114, 6148–6150. [Google Scholar] [CrossRef] [PubMed]
- Calvario, G.; Sierra, B.; Alarcón, T.E.; Hernandez, C.; Dalmau, O. A Multi-Disciplinary Approach to Remote Sensing through Low-Cost UAVs. Sensors 2017, 17, 1411. [Google Scholar] [CrossRef] [PubMed]
- Severtson, D.; Callow, N.; Flower, K.; Neuhaus, A.; Olejnik, M.; Nansen, C. Unmanned aerial vehicle canopy reflectance data detects potassium deficiency and green peach aphid susceptibility in canola. Precis. Agric. 2016, 17, 659–677. [Google Scholar] [CrossRef]
- Fang, S.; Tang, W.; Peng, Y.; Gong, Y.; Dai, C.; Chai, R.; Liu, K. Remote estimation of vegetation fraction and flower fraction in oilseed rape with unmanned aerial vehicle data. Remote Sens. 2016, 8, 416. [Google Scholar] [CrossRef]
- Gnädinger, F.; Schmidhalter, U. Digital Counts of Maize Plants by Unmanned Aerial Vehicles (UAVs). Remote Sens. 2017, 9, 544. [Google Scholar] [CrossRef]
- Maresma, Á.; Ariza, M.; Martínez, E.; Lloveras, J.; Martínez-Casasnovas, J.A. Analysis of vegetation indices to determine nitrogen application and yield prediction in maize (Zea mays L.) from a standard UAV service. Remote Sens. 2016, 8, 973. [Google Scholar] [CrossRef]
- Roosjen, P.P.; Suomalainen, J.M.; Bartholomeus, H.M.; Kooistra, L.; Clevers, J.G. Mapping Reflectance Anisotropy of a Potato Canopy Using Aerial Images Acquired with an Unmanned Aerial Vehicle. Remote Sens. 2017, 9, 417. [Google Scholar] [CrossRef]
- Roosjen, P.P.; Suomalainen, J.M.; Bartholomeus, H.M.; Clevers, J.G. Hyperspectral Reflectance Anisotropy Measurements Using a Pushbroom Spectrometer on an Unmanned Aerial Vehicle—Results for Barley, Winter Wheat, and Potato. Remote Sens. 2016, 8, 909. [Google Scholar] [CrossRef]
- Schirrmann, M.; Giebel, A.; Gleiniger, F.; Pflanz, M.; Lentschke, J.; Dammer, K.H. Monitoring Agronomic Parameters of Winter Wheat Crops with Low-Cost UAV Imagery. Remote Sens. 2016, 8, 706. [Google Scholar] [CrossRef]
- Du, M.; Noguchi, N. Monitoring of Wheat Growth Status and Mapping of Wheat Yield’s within-Field Spatial Variations Using Color Images Acquired from UAV-camera System. Remote Sens. 2017, 9, 289. [Google Scholar] [CrossRef]
- Holman, F.H.; Riche, A.B.; Michalski, A.; Castle, M.; Wooster, M.J.; Hawkesford, M.J. High throughput field phenotyping of wheat plant height and growth rate in field plot trials using UAV based remote sensing. Remote Sens. 2016, 8, 1031. [Google Scholar] [CrossRef]
- Yue, J.; Yang, G.; Li, C.; Li, Z.; Wang, Y.; Feng, H.; Xu, B. Estimation of Winter Wheat Above-ground Biomass Using Unmanned Aerial Vehicle-based Snapshot Hyperspectral Sensor and Crop Height Improved Models. Remote Sens. 2017, 9, 708. [Google Scholar] [CrossRef]
- Burkart, A.; Aasen, H.; Alonso, L.; Menz, G.; Bareth, G.; Rascher, U. Angular dependency of hyperspectral measurements over wheat characterized by a novel UAV based goniometer. Remote Sens. 2015, 7, 725–746. [Google Scholar] [CrossRef]
- Luna, I.; Lobo, A. Mapping Crop Planting Quality in Sugarcane from UAV Imagery: A Pilot Study in Nicaragua. Remote Sens. 2016, 8, 500. [Google Scholar] [CrossRef]
- Mathews, A.J.; Jensen, J.L. Visualizing and quantifying vineyard canopy LAI using an unmanned aerial vehicle (UAV) collected high density structure from motion point cloud. Remote Sens. 2013, 5, 2164–2183. [Google Scholar] [CrossRef]
- Matese, A.; Toscano, P.; Di Gennaro, S.F.; Genesio, L.; Vaccari, F.P.; Primicerio, J.; Belli, C.; Zaldei, A.; Bianconi, A.; Gioli, B. Intercomparison of UAV, aircraft and satellite remote sensing platforms for precision viticulture. Remote Sens. 2015, 7, 2971–2990. [Google Scholar] [CrossRef]
- Horton, R.; Cano, E.; Bulanon, D.; Fallahi, E. Peach Flower Monitoring Using Aerial Multispectral Imaging. J. Imaging 2017, 3, 2. [Google Scholar] [CrossRef]
- Fernández, T.; Pérez, J.L.; Cardenal, J.; Gómez, J.M.; Colomo, C.; Delgado, J. Analysis of landslide evolution affecting olive groves using uav and photogrammetric techniques. Remote Sens. 2016, 8, 837. [Google Scholar] [CrossRef]
- Ortega-Farías, S.; Ortega-Salazar, S.; Poblete, T.; Kilic, A.; Allen, R.; Poblete-Echeverría, C.; Ahumada-Orellana, L.; Zuñiga, M.; Sepúlveda, D. Estimation of energy balance components over a drip-irrigated olive orchard using thermal and multispectral cameras placed on a helicopter-based unmanned aerial vehicle (UAV). Remote Sens. 2016, 8, 638. [Google Scholar] [CrossRef]
- Díaz-Varela, R.A.; de la Rosa, R.; León, L.; Zarco-Tejada, P.J. High-resolution airborne UAV imagery to assess olive tree crown parameters using 3D photo reconstruction: Application in breeding trials. Remote Sens. 2015, 7, 4213–4232. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Hexapilots, Unmanned Aerial Services. Available online: http://wm1bc4730.homepage.t-online.de/hexawp/ (accessed on 13 May 2017).
- Apus Systems Intelligent Geocoding. Available online: https://www.apus-systems.com (accessed on 25 July 2017).
- Mund, J.-P.; Cremer, T.; Krause, S. Potenzial und Perspektive: Drohnen in der Forstwirtschaft. AFZ DerWald 2017, 17, 43–46. [Google Scholar]
- Festmeter—Ihr Wald. Available online: http://www.festmeter.at (accessed on 12 May 2017).
- Näsi, R.; Honkavaara, E.; Lyytikäinen-Saarenmaa, P.; Blomqvist, M.; Litkey, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Tanhuanpää, T.; Holopainen, M. Using UAV-based photogrammetry and hyperspectral imaging for mapping bark beetle damage at tree-level. Remote Sens. 2015, 7, 15467–15493. [Google Scholar] [CrossRef]
- Lehmann, J.R.K.; Nieberding, F.; Prinz, T.; Knoth, C. Analysis of unmanned aerial system-based CIR images in forestry—A new perspective to monitor pest infestation levels. Forests 2015, 6, 594–612. [Google Scholar] [CrossRef]
- Rucon Engineering, Über RUCON Engineering. Available online: http://rucon.de (accessed on 20 March 2017).
- Duan, F.; Wan, Y.; Deng, L. A Novel Approach for Coarse-to-Fine Windthrown Tree Extraction Based on Unmanned Aerial Vehicle Images. Remote Sens. 2017, 9, 306. [Google Scholar] [CrossRef]
- Sohns, V. LIGNA 2017 zeigt Forsttechnik und Produktionskette. AFZ DerWald 2017, 13, 35–37. [Google Scholar]
- Eusemann, P.; Liesebach, M.; Liesebach, H. Mit Drohnen Ernteaussichten in Saatgutbeständen erkunden. AFZ DerWald 2017, 10, 28–30. [Google Scholar]
- Chianucci, F.; Disperati, L.; Guzzi, D.; Bianchini, D.; Nardino, V.; Lastri, C.; Rindinella, A.; Corona, P. Estimation of canopy attributes in beech forests using true colour digital images from a small fixed-wing UAV. Int. J. Appl. Earth Obs. Geoinform. 2016, 47, 60–68. [Google Scholar] [CrossRef]
- Cunliffe, A.M.; Brazier, R.E.; Anderson, K. Ultra-fine grain landscape-scale quantification of dryland vegetation structure with drone-acquired structure-from-motion photogrammetry. Remote Sens. Environ. 2016, 183, 129–143. [Google Scholar] [CrossRef]
- McNeil, B.E.; Pisek, J.; Lepisk, H.; Flamenco, E.A. Measuring leaf angle distribution in broadleaf canopies using UAVs. Agric. For. Meteorol. 2016, 218, 204–208. [Google Scholar] [CrossRef]
- MacInnis, G.; Forrest, J. Quantifying pollen deposition with macro photography and ‘stigmagraphs’. J. Pollinat. Ecol. 2017, 20, 13–21. [Google Scholar]
- Lino, A.C.L.; Sanches, J.; Moraes, G.; Dias-Tagliacozzo, I.M.D.; Augusto, F.; Lima, B.; Nascimento, T.S. Flower classification supported by digital imaging techniques. J. Inform. Technol. Agric. 2011, 4, 1–6. [Google Scholar]
- Deutscher Wetterdienst DWD. Archiv Monats- und Tageswerte. Available online: http://www.dwd.de (accessed on 10 June 2017).
- Landgraf, D.; Böcker, L.; Wiesner, S.; Kempe, K. Energiewald Kostebrau—Chancen und Risiken für die Stadt Lauchhammer. In Proceedings of the Tagungsband zur Fachtagung Anbau und Nutzung von Bäumen auf landwirtschaftlichen Flächen, Freiburg, Germany, 3 July 2007; pp. 39–45. [Google Scholar]
- yWorks, the Diagramming Company. Available online: http://www.yworks.com/products/yed/download (accessed on 20 January 2017).
- MAPIR—Survey2 Camera—Visible Light RGB. Available online: https://www.mapir.camera/products/survey2-camera-visible-light-rgb (accessed on 28 May 2017).
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Grouven, U.; Bender, R.; Ziegler, A.; Lange, S. Der kappa-koeffizient. Dtsch. Med. Wochenschr. 2007, 132, e65–e68. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. Available online: https://pdfs.semanticscholar.org/7e73/43a5608fff1c68c5259db0c77b9193f1546d.pdf (accessed on 18 October 2017). [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2016. Available online: https://www.R-project.org/ (accessed on 23 November 2016).
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2009. [Google Scholar]
- Urbanek, S. png: Read and Write PNG Images. R Package Version 0.1–7. Available online: https://CRAN.R-project.org/package=png (accessed on 3 July 2017).
- Murrell, P. gridGraphics: Redraw Base Graphics Using ‘Grid’ Graphics. R Package Version 0.2. 2017. Available online: https://CRAN.R-project.org/package=gridGraphics (accessed on 3 July 2017).
- Droege, G. Das Imkerbuch; VEB Deutscher Landwirtschaftsverlag: Berlin, Germany, 1989; p. 106. ISBN 3331002623. [Google Scholar]
- Dachmarke—Über Bienen und Honig. Available online: http://dachmarke.com/produkte/bienenwissen/ (accessed on 25 June 2017).
- Die Honigmacher. Winterbiene. Available online: http://www.die-honigmacher.de/kurs3/seite_15103.html (accessed on 20 June 2017).
- Imkerverein Büchertal. Biologie des Bienenvolkes. Available online: http://www.imkerverein-buechertal.de/Biologie_Bienenvolk.php (accessed on 20 May 2017).
- Deutsches Institut für Normung e. V. Untersuchung von Honig—Bestimmung der Relativen Pollenhaüfigkeit, DIN 10760. 2002, pp. 1–6. Available online: https://www.din.de/de/mitwirken/normenausschuesse/krdl/din-spec/wdc-beuth:din21:47633680 (accessed on 13 September 2017).
- Cruzan, M.B.; Weinstein, B.G.; Grasty, M.R.; Kohrn, B.F.; Hendrickson, E.C.; Arredondo, T.M.; Thompson, P.G. Small unmanned aerial vehicles (micro-UAVs, drones) in plant ecology. Appl. Plant Sci. 2016, 4, 1600041. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.R.; Potts, B.M.; Neilsen, W.A.; Joyce, K.R. The effect of tree spacing on the production of flowers in Eucalyptus nitens. Aust. For. 2006, 69, 299–304. [Google Scholar] [CrossRef]
- Yarra Ranges Shire Council, Eucalyptus Nitens. Available online: http://fe.yarraranges.vic.gov.au/Residents/Trees_Vegetation/Yarra_Ranges_Plant_Directory/Yarra_Ranges_Local_Plant_Directory/Upper_Storey/Trees_5m/Eucalyptus_nitens (accessed on 13 September 2017).
- Hart, S.J.; Veblen, T.T. Detection of spruce beetle-induced tree mortality using high-and medium-resolution remotely sensed imagery. Remote Sens. Environ. 2015, 168, 134–145. [Google Scholar] [CrossRef]
- Knoth, C.; Klein, B.; Prinz, T.; Kleinebecker, T. Unmanned aerial vehicles as innovative remote sensing platforms for high-resolution infrared imagery to support restoration monitoring in cut-over bogs. Appl. Veg. Sci. 2013, 16, 509–517. [Google Scholar] [CrossRef]
- Bálint, M.; Domisch, S.; Engelhardt, C.H.M.; Haase, P.; Lehrian, S.; Sauer, J.; Theissinger, K.; Pauls, S.U.; Nowak, C. Cryptic biodiversity loss linked to global climate change. Nat. Clim. Chang. 2011, 1, 313. [Google Scholar] [CrossRef]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, A.; McCallum, B. A World without Bees, The Mysterious Decline of the Honeybee-and What is Means for Us; Guardian Books: London, UK, 2009; ISBN 9780852651315. [Google Scholar]






| FA Above Starting Point (m) | FA Above Ground (m) | FA Above Crowns (m) |
|---|---|---|
| 20.0 | 8.2 | 2.6 |
| 25.0 | 13.2 | 7.6 |
| 30.0 | 18,2 | 12.6 |
| 35.0 | 23.2 | 17.6 |
| 40.0 | 28.2 | 22.6 |
| 45.0 | 33.2 | 27.6 |
| 50.0 | 38.2 | 32.6 |
| 60.0 | 48.2 | 42.6 |
| 70.0 | 58.2 | 52.6 |
| 80.0 | 68.2 | 62.6 |
| 90.0 | 78.2 | 72.6 |
| 100.0 | 88.2 | 82.6 |
| 110.0 | 98.2 | 92.6 |
| Flight Line (South-North) | FA Above Starting Point (m) | FA Above Ground (m) | FA Above Crowns (m) | FOV (Above Ground) (m) | (m) |
|---|---|---|---|---|---|
| A | 17.0 | 10.8 | 5.5 | 16.8 | 12.6 |
| B | 18.0 | 10.5 | 5.3 | 16.3 | 12.3 |
| C | 19.0 | 10.0 | 4.5 | 15.6 | 11.7 |
| D | 19.0 | 8.5 | 3.0 | 13.2 | 9.9 |
| E | 20.0 | 8.2 | 2.6 | 12.8 | 9.6 |
| F | 22.0 | 9.1 | 4.4 | 14.2 | 10.6 |
| G | 24.0 | 10.0 | 4.5 | 15.6 | 11.7 |
| H | 24.0 | 9.2 | 4.2 | 14.3 | 10.7 |
| Mean | 20.4 | 9.5 | 4.2 | 14.9 | 11.1 |
| Tree Number | Inflorescence per Tree | Flower per Inflorescence | Flower per Tree | Flower Surface 2D (cm2) | Flower Mass (Total) | Flower Mass (1.0 cm2) | Flower Volume 3D (cm3) |
|---|---|---|---|---|---|---|---|
| 1 | 197 | 17 (1–34) | 3409 | 2.44 (1.64–3.26) | 133 (93–168) | 24 (14–35) | 5.78 (4.27–9.26) |
| 2 | 658 | 24 (1–41) | 15,559 | 1.75 (0.81–2.49) | 100 (60–142) | 18 (11–25) | 5.69 (3.77–8.13) |
| 3 | 324 | 19 (2–30) | 6001 | 1.84 (1.21–2.57) | 119 (82–215) | 18 (10–24) | 6.73 (4.66–11.46) |
| 4 | 198 | 12 (1–25) | 2427 | 1.90 (1.32–2.59) | 90 (60–127) | 18 (10–31) | 5.15 (3.29–8.53) |
| 5 | 204 | 28 (6–37) | 5641 | 1.82 (1.29–2.26) | 115 (77–226) | 19 (8–27) | 6.18 (4.31–12.19) |
| 6 | 84 | 23 (2–38) | 1913 | 2.12 (1.57–2.99) | 105 (80–130) | 16 (9–30) | 6.77 (3.63–10.52) |
| 7 | 118 | 16 (7–31) | 1896 | 1.58 (1.22–2.10) | 95 (72–123) | 18 (11–25) | 5.42 (3.98–7.82) |
| Mean | 255 | 20 (3–34) | 5264 | 1.92 (0.81–3.26) | 108 (60–226) | 19 (8–35) | 5.96 (3.29–12.19) |
| Plants | Classification | Honey Sample 1 | Honey Sample 2 |
|---|---|---|---|
| Nectar-giving | >45% | – | Brassicaceae |
| 45–15% | R. pseudoacacia, Brassicaceae | – | |
| <15–3% | Rubus, Salix, Cyanus segetum Hill | Pyrinae, R. pseudoacacia, Rubus, Salix | |
| <3% | Aesculus hippocastanum, Acer, Sinapis, Trifolium, Asteraceae, Pyrinae, Cornus, Gleditsia | Asteraceae, Aesculus hippocastanum, Liliaceae, Tilia, Weigela, Lytherum, Carduoideae, Frangula alnus | |
| Nectarless | 13% | Poaceae, Papaver, Asteraceae, Pyrinae, Cornus | – |
| 6.4% | – | Papaver, Quercus, Betula, Pinus, Sambucus, Filipendula ulmaria, Rosaceae |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carl, C.; Landgraf, D.; Van der Maaten-Theunissen, M.; Biber, P.; Pretzsch, H. Robinia pseudoacacia L. Flower Analyzed by Using An Unmanned Aerial Vehicle (UAV). Remote Sens. 2017, 9, 1091. https://doi.org/10.3390/rs9111091
Carl C, Landgraf D, Van der Maaten-Theunissen M, Biber P, Pretzsch H. Robinia pseudoacacia L. Flower Analyzed by Using An Unmanned Aerial Vehicle (UAV). Remote Sensing. 2017; 9(11):1091. https://doi.org/10.3390/rs9111091
Chicago/Turabian StyleCarl, Christin, Dirk Landgraf, Marieke Van der Maaten-Theunissen, Peter Biber, and Hans Pretzsch. 2017. "Robinia pseudoacacia L. Flower Analyzed by Using An Unmanned Aerial Vehicle (UAV)" Remote Sensing 9, no. 11: 1091. https://doi.org/10.3390/rs9111091
APA StyleCarl, C., Landgraf, D., Van der Maaten-Theunissen, M., Biber, P., & Pretzsch, H. (2017). Robinia pseudoacacia L. Flower Analyzed by Using An Unmanned Aerial Vehicle (UAV). Remote Sensing, 9(11), 1091. https://doi.org/10.3390/rs9111091