
Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices


Abstract
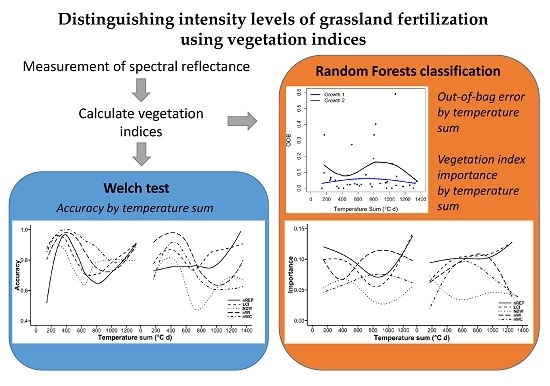
:
1. Introduction
- Each plant community on grassland is characterized by a unique VI development throughout the growing season.
- The performance of each VI for distinguishing such plant communities varies throughout the growing season.
- VIs sensitive to a certain plant property (e.g., chlorophyll content) may allow a successful classification at times when VIs sensitive to other plant properties (e.g., water content, biomass, etc.) fail to classify correctly.
- Combining several VIs for grassland classification allows a temporally more stable classification of grassland communities than one single VI because for distinguishing communities at a point in time, a set of optimal VIs is selected.
2. Materials and Methods
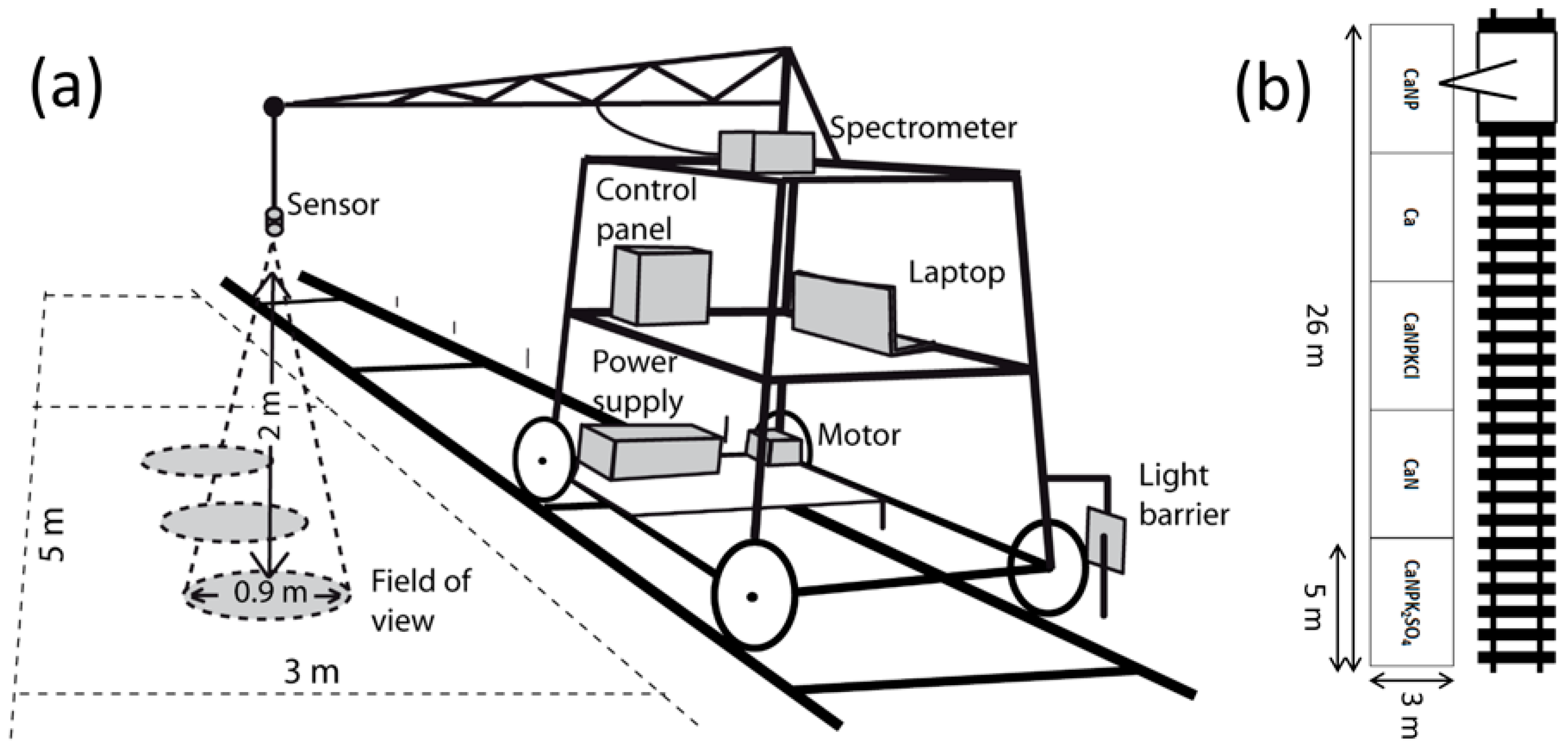
2.1. Study Area
2.2. Spectral Measurements
2.3. Calculation of the Temperature Sum
2.4. Calculation of the VIs
- VIs sensitive to green vegetation, biomass and leaf area,
- VIs sensitive to plant chlorophyll content,
- VIs sensitive to the plants’ content in lignin, nitrogen, water, pigments or to plant physiological performance and phaeophytization (environmental stress).
2.5. Welch Test
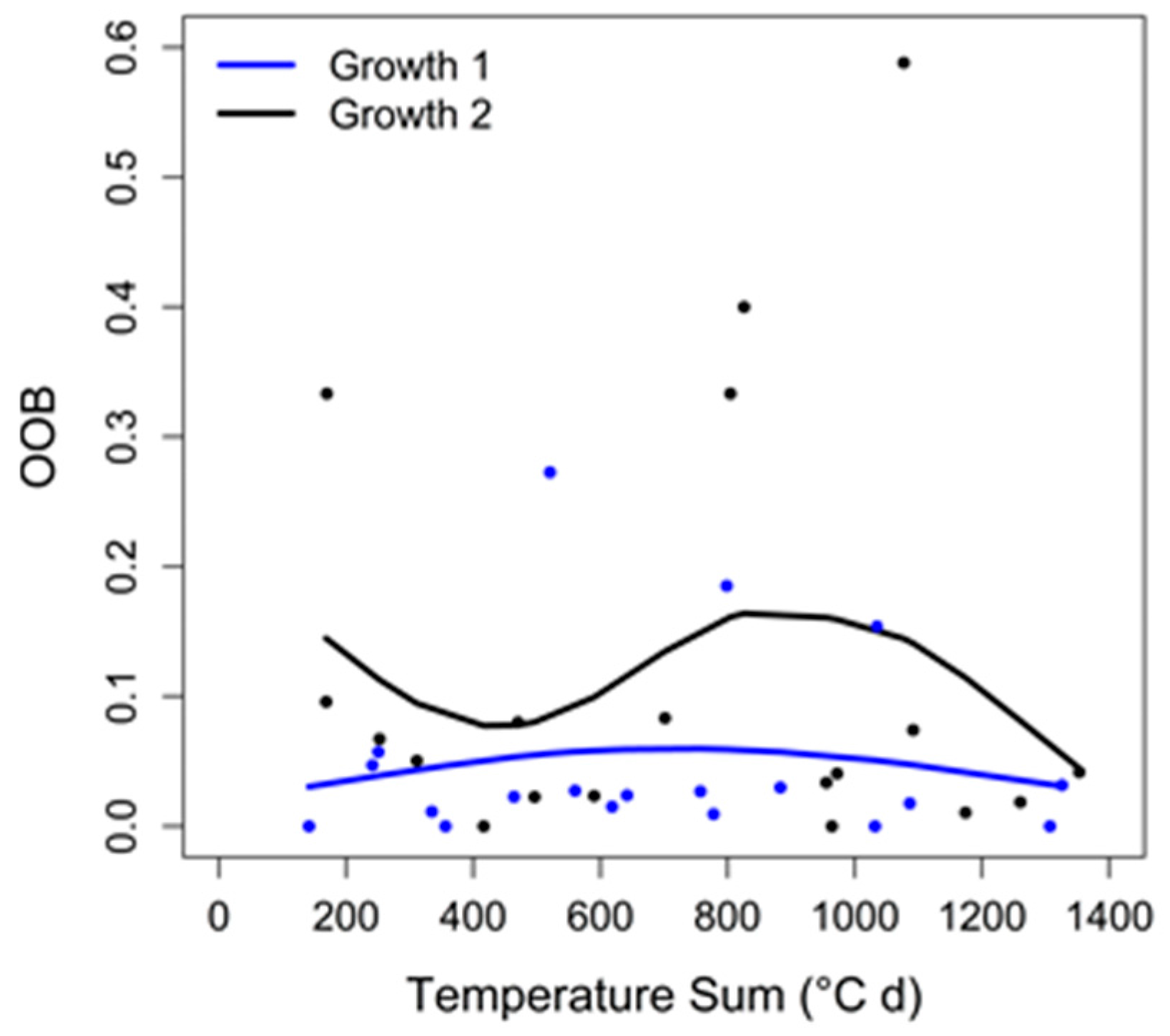
2.6. Random Forests Classification
3. Results
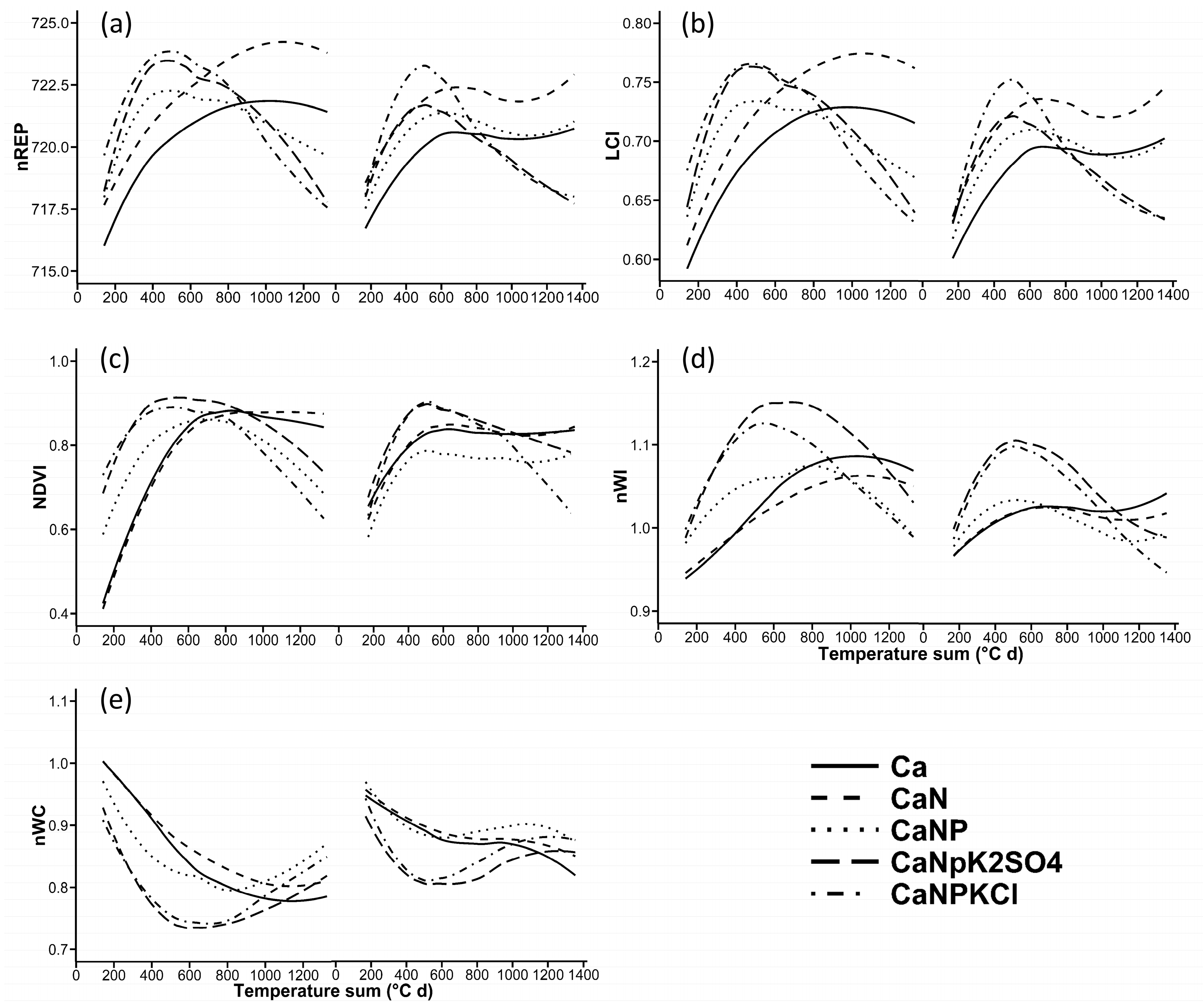
3.1. Seasonal Curves of the VIs
3.2. Distinguishing the Grassland Communities Using the Welch Test
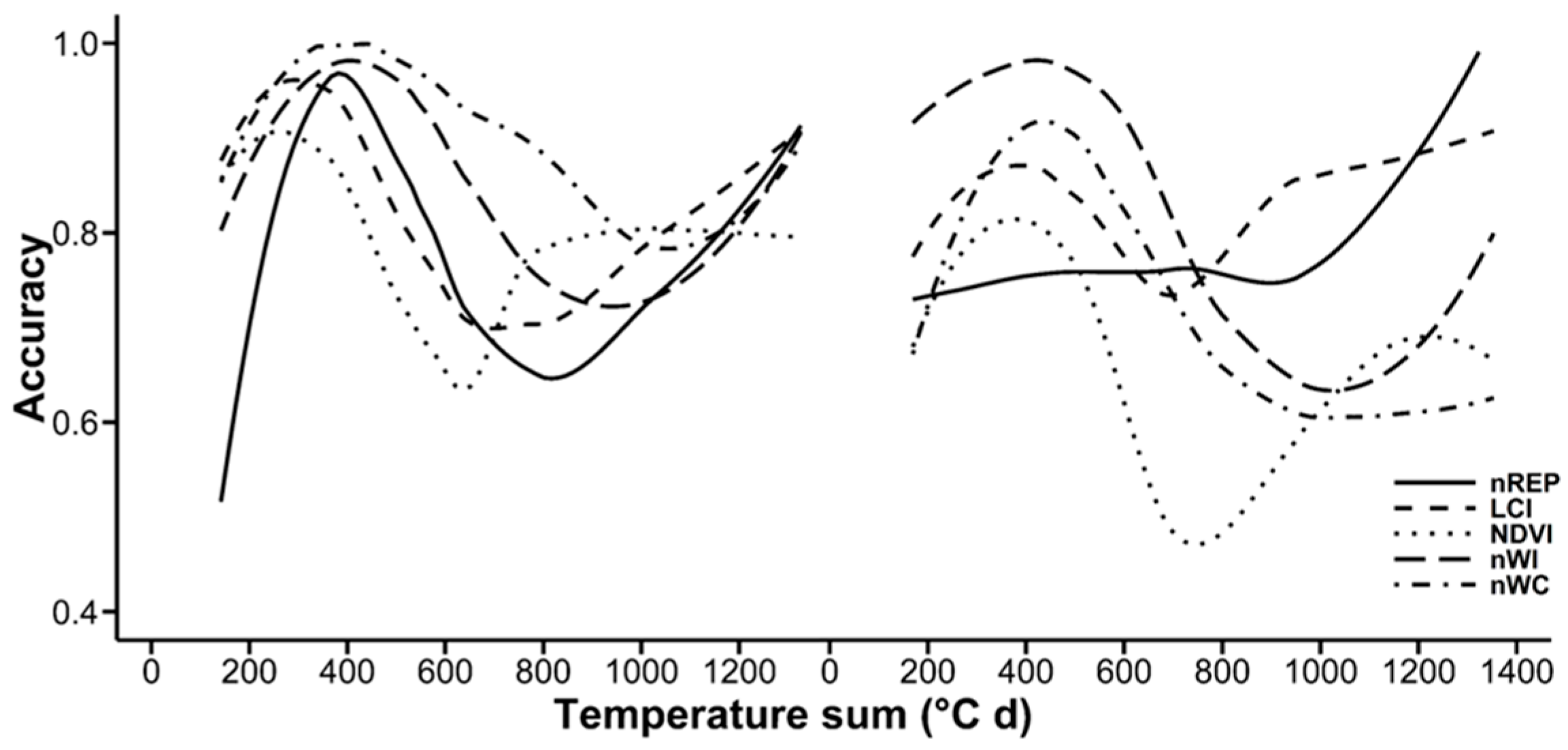
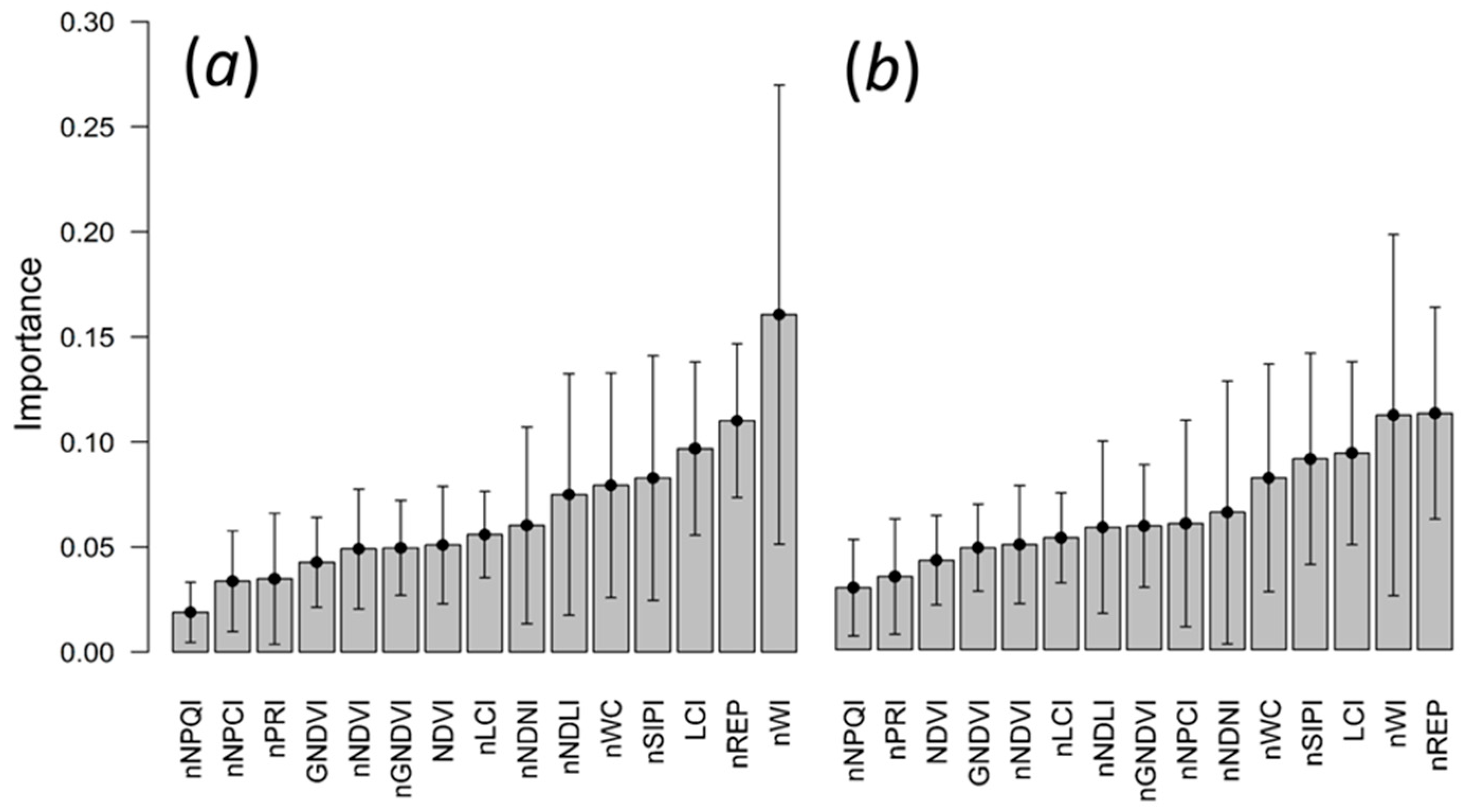
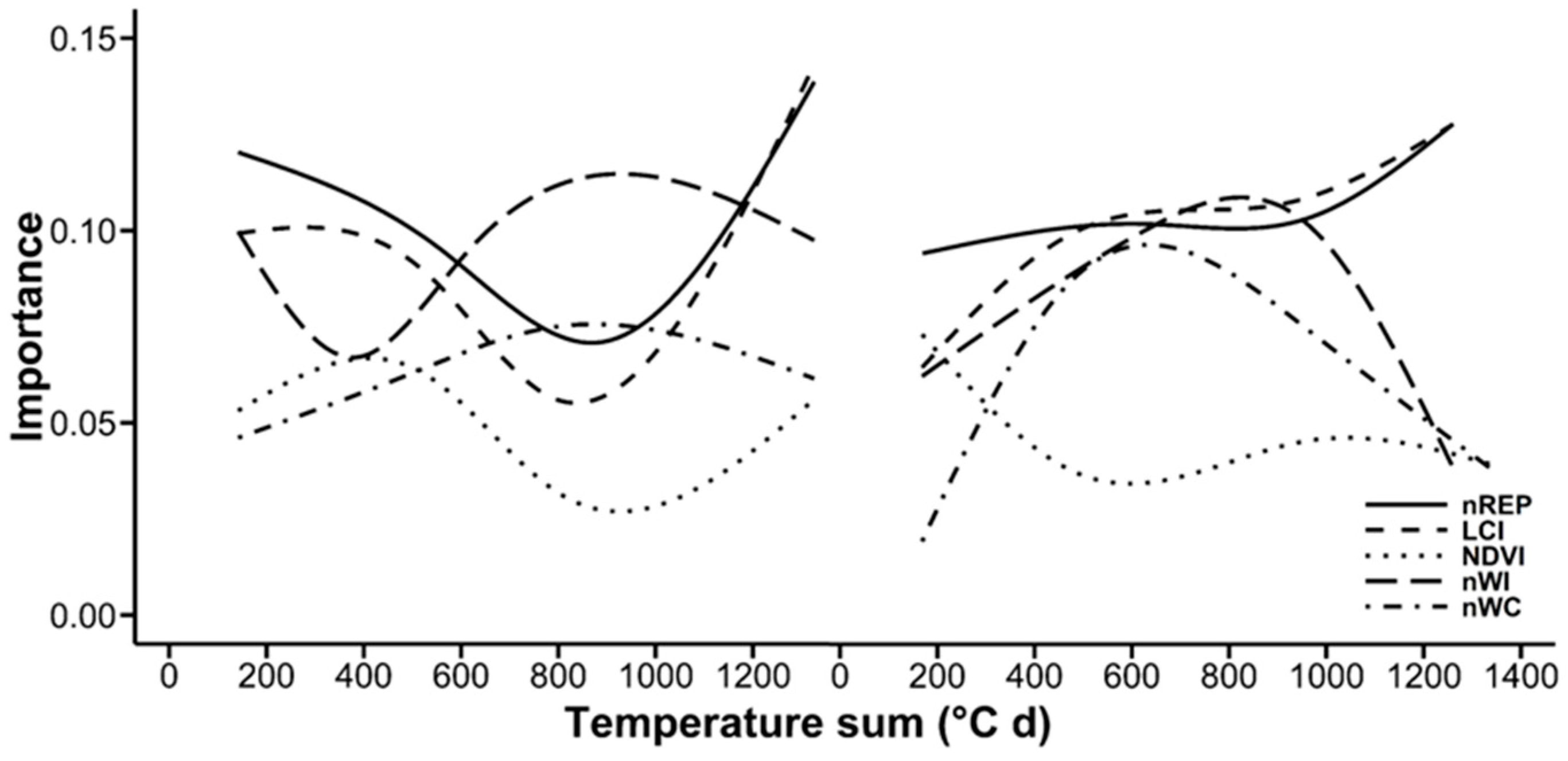
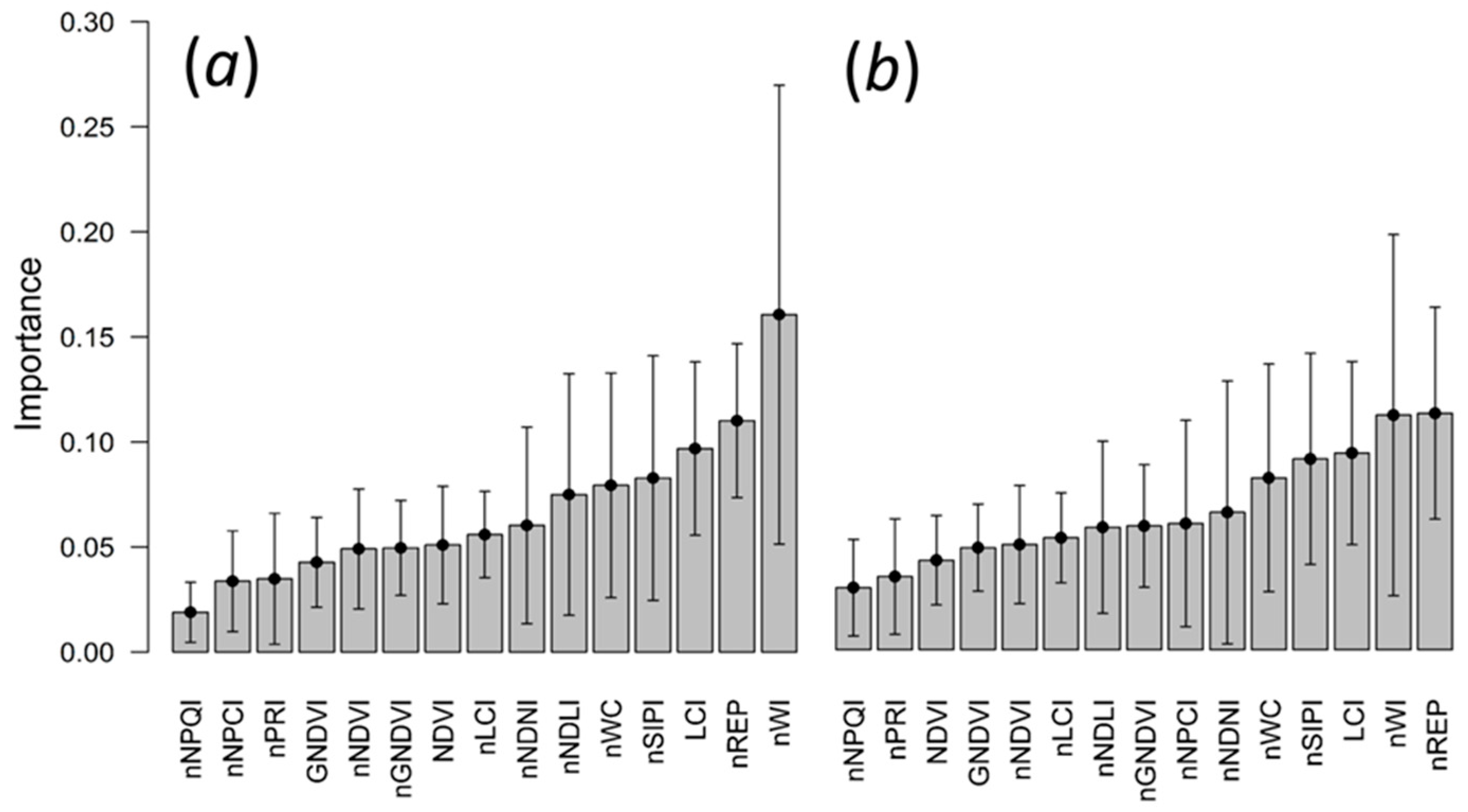
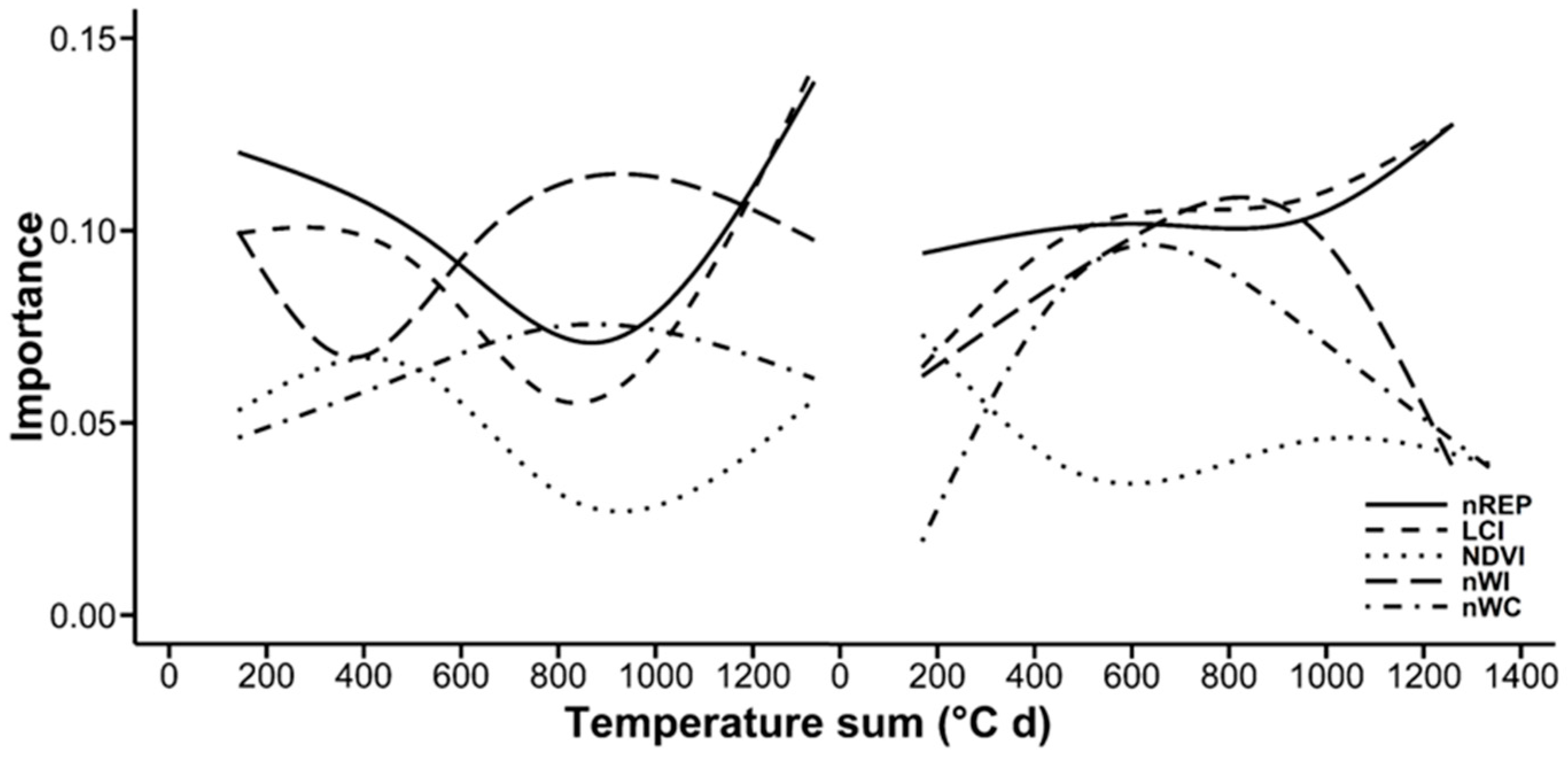
3.3. Random Forests Classification
4. Discussion
4.1. Critical Reflection on the Experimental Settings
4.2. Seasonal Curves of the VIs
4.3. Testing the Classification Accuracy of the Fifteen VIs Using the Welch Test
4.4. Random Forests Classification
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth 1 | nWI | nREP | LCI | nSIPI | nWC | nNDLI | nNDNI | nLCI | NDVI | nGNDVI | nNDVI | GNDVI | nPRI | nNPCI | nNPQI | |
| nWI | - | - | ** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| nREP | - | - | - | * | ** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| LCI | ** | - | - | - | - | - | ** | *** | *** | *** | *** | *** | *** | *** | *** | |
| nSIPI | ** | * | - | - | - | - | - | ** | ** | ** | ** | *** | *** | *** | *** | |
| nWC | *** | ** | - | - | - | - | - | - | * | * | * | ** | *** | *** | *** | |
| nNDLI | *** | ** | - | - | - | - | - | - | * | * | * | ** | *** | *** | *** | |
| nNDNI | *** | *** | ** | - | - | - | - | - | - | - | - | - | * | ** | *** | |
| nLCI | *** | *** | *** | ** | - | - | - | - | - | - | - | - | ** | ** | *** | |
| NDVI | *** | *** | *** | ** | * | * | - | - | - | - | - | - | * | ** | *** | |
| nGNDVI | *** | *** | *** | ** | * | * | - | - | - | - | - | - | * | * | *** | |
| nNDVI | *** | *** | *** | ** | * | * | - | - | - | - | - | - | - | ** | *** | |
| GNDVI | *** | *** | *** | *** | ** | ** | - | - | - | - | - | - | - | ** | *** | |
| nPRI | *** | *** | *** | *** | *** | *** | * | ** | * | * | - | - | - | - | - | |
| nNPCI | *** | *** | *** | *** | *** | *** | ** | ** | ** | * | ** | ** | - | - | - | |
| nNPQI | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | - | - | - | |
| Growth 2 | nREP | nWI | LCI | nSIPI | nWC | nNDNI | nNPCI | nGNDVI | nNDLI | nLCI | nNDVI | GNDVI | NDVI | nPRI | nNPQI | |
| nREP | - | - | - | - | * | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| nWI | - | - | - | - | - | * | ** | ** | ** | ** | ** | *** | *** | *** | *** | |
| LCI | - | - | - | - | - | * | *** | ** | ** | *** | *** | *** | *** | *** | *** | |
| nSIPI | - | - | - | - | - | - | ** | ** | ** | *** | *** | *** | *** | *** | *** | |
| nWC | * | - | - | - | - | - | * | - | - | ** | ** | ** | *** | *** | *** | |
| nNDNI | ** | * | ** | - | - | - | - | - | - | - | - | - | - | ** | ** | |
| nNPCI | *** | ** | *** | ** | * | - | - | - | - | - | - | - | - | *** | *** | |
| nGNDVI | *** | ** | ** | ** | - | - | - | - | - | - | - | - | - | * | ** | |
| nNDLI | *** | ** | ** | ** | - | - | - | - | - | - | - | - | - | * | * | |
| nLCI | *** | ** | *** | *** | ** | - | - | - | - | - | - | - | - | ** | *** | |
| nNDVI | *** | ** | *** | *** | ** | - | - | - | - | - | - | - | - | *** | *** | |
| GNDVI | *** | *** | *** | *** | ** | - | - | - | - | - | - | - | - | * | ** | |
| NDVI | *** | *** | *** | *** | *** | - | - | - | - | - | - | - | - | * | * | |
| nPRI | *** | *** | *** | *** | *** | ** | *** | * | * | ** | *** | * | * | - | - | |
| nNPQI | *** | *** | *** | *** | *** | ** | *** | ** | * | *** | *** | ** | * | - | - |
| Year | Growth | T∑ (°C d) | CSH (cm) | SD (CSH) |
|---|---|---|---|---|
| 2013 | 1 | 947 | 20.59 | 10.63 |
| 2 | 964 | 6.49 | 2.51 | |
| 1077 | 4.62 | 2.15 | ||
| 1378 | 6.98 | 2.95 | ||
| 2014 | 1 | 464 | 5.52 | 2.93 |
| 560 | 7.75 | 4.23 | ||
| 757 | 18.12 | 8.47 | ||
| 883 | 17.72 | 10.09 | ||
| 1017 | 17.18 | 9.64 | ||
| 1307 | 24.5 | 18.21 | ||
| 2 | 210 | 5.17 | 1.18 | |
| 804 | 10.54 | 5.37 | ||
| 972 | 12.94 | 4.90 | ||
| 1174 | 13.28 | 4.97 | ||
| 1353 | 11.21 | 4.57 |
References
- Price, K.P.; Crooks, T.J.; Martinko, E.A. Grasslands across time and scale: A remote sensing perspective. Photogramm. Eng. Remote Sens. 2001, 67, 414–420. [Google Scholar]
- Schut, A.G.T.; Ketelaars, J.J.M.H.; Meulemann, J.; Kornet, J.G.; Lockhorst, C. Novel imaging spectroscopy for grass sward characterization. Biosyst. Eng. 2002, 82, 131–141. [Google Scholar] [CrossRef]
- Gianelle, D.; Vescovo, L. Determination of green herbage ratio in grasslands using spectral reflectance. Methods and ground measurements. Int. J. Remote Sens. 2007, 28, 931–942. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Aragón, R.; Oesterheld, M. Linking vegetation heterogeneity and functional attributes of temperate grasslands through remote sensing. Appl. Veg. Sci. 2008, 11, 117–130. [Google Scholar] [CrossRef]
- Hill, M.J. Vegetation index suites as indicators of vegetation state in grassland and savanna: An analysis with simulated SENTINEL 2 data for a North American transect. Remote Sens. Environ. 2013, 137, 94–111. [Google Scholar] [CrossRef]
- Wardlow, B.D.; Egbert, S.L.; Kastens, J.H. Analysis of time-series MODIS 250 m vegetation index data for crop classification in the U.S. Central Great Plains. Remote Sens. Environ. 2007, 108, 290–310. [Google Scholar] [CrossRef]
- Psomas, A.; Zimmermann, N.E.; Kneubühler, M.; Kellenberger, T.; Itten, K. Seasonal variability in spectral reflectance for discriminating grasslands along a dry-mesic gradient in Switzerland. In Proceedings of the 4th EARSeL Workshop on Imaging Spectroscopy, New Quality in Environmental Studies, Warsaw, Poland, 27–30 April 2005.
- Poças, I.; Cunha, M.; Pereira, L.S. Dynamics of mountain semi-natural grassland meadows inferred from SPOT-VEGETATION and field spectroradiometer data. Int. J. Remote Sens. 2012, 33, 4334–4355. [Google Scholar] [CrossRef]
- Fava, F.; Colombo, R.; Bocchi, S.; Meroni, M.; Sitzia, M.; Fois, N.; Zucca, C. Identification of hyperspectral vegetation indices for Mediterranean pasture characterization. Int. J. Appl. Earth Obs. Geoinf. 2009, 11, 233–243. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; Brown de Colstoun, E.; McMurtey, J.E. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- He, Y.; Guo, X.; Wilmshurst, J. Studying mixed grassland ecosystems I: Suitable hyperspectral vegetation indices. Can. J. Remote Sens. 2006, 32, 98–107. [Google Scholar] [CrossRef]
- Asner, G.P. Biophysical and Biochemical Sources of Variability in Canopy Reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- Kumar, L.; Schmidt, K.; Dury, S.; Skidmore, A. Imaging spectrometry and vegetation science. In Imaging Spectrometry; van de Meer, F., de Jong, S.M., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 111–155. [Google Scholar]
- Lucas, R.M.; Honzak, M.; Foody, G.M.; Curran, P.J.; Corves, C. Characterizing tropical secondary forests using multi-temporal Landsat sensor imagery. Int. J. Remote Sens. 1993, 14, 3061–3067. [Google Scholar] [CrossRef]
- Roberts, D.A.; Numata, I.; Holmes, K.; Batista, G.; Krug, T.; Monteiro, A.; Powell, B.; Chadwick, O.A. Large area mapping of land-cover change in Rondonia using multitemporal spectral mixture analysis and decision tree classifiers. J. Geophys. Res. Atmos. 2002, 107, D20. [Google Scholar] [CrossRef]
- Adams, J.B.; Sabol, D.E.; Kapos, V.; Almeida Filho, R.; Roberts, D.A.; Smith, M.O.; Gillespie, A.R. Classification of multispectral images based on fractions of endmembers: Application to land-cover change in the Brazilian Amazon. Remote Sens. Environ. 1995, 52, 137–154. [Google Scholar] [CrossRef]
- Numata, I.; Roberts, D.A.; Chadwick, O.A.; Schimel, J.P.; Galvao, L.S.; Soares, J.V. Evaluation of hyperspectral data for pasture estimate in the Brazilian Amazon using field and imaging spectrometers. Remote Sens. Environ. 2008, 112, 1569–1583. [Google Scholar] [CrossRef]
- Welch, B.L. The significance of the difference between two means when the population variances are unequal. Biometrika 1938, 29, 350–362. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Schellberg, J.; Möseler, B.M.; Kühbauch, W.; Rademacher, I.F. Long-term effects of fertilizer on soil nutrient concentration, yield, forage quality and floristic composition of a hay meadow in the Eifel mountains, Germany. Grass Forage Sci. 1999, 54, 195–207. [Google Scholar] [CrossRef]
- Hejcman, M.; Češková, M.; Schellberg, J.; Pätzold, S. The Rengen Grassland Experiment: Effect of Soil Chemical Properties on Biomass Production, Plant Species Composition and Species Richness. Folia Geobot. 2010, 45, 125–142. [Google Scholar] [CrossRef]
- Hejcman, M.; Klaudisová, M.; Schellberg, J.; Honsová, D. The Rengen Grassland Experiment: Plant species composition after 64 years of fertilizer application. Agric. Ecosyst. Environ. 2007, 122, 259–266. [Google Scholar] [CrossRef]
- Chytrý, M.; Hejcman, M.; Hennekens, S.M.; Schellberg, J. Changes in vegetation types and Ellenberg indicator values after 65 years of fertilizer application in the Rengen Grassland Experiment, Germany. Appl. Veg. Sci. 2009, 12, 167–176. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Smith, R.B.; De Pauw, E. Evaluation of narrowband and broadband vegetation indices for determining optimal hyperspectral wavebands for agricultural crop characterization. Photogramm. Eng. Remote Sens. 2002, 68, 607–622. [Google Scholar]
- Gamon, J.A.; Cheng, Y.; Claudio, H.; MacKinney, L.; Sims, D.A. A mobile tram system for systematic sampling of ecosystem optical properties. Remote Sens. Environ. 2006, 103, 246–254. [Google Scholar] [CrossRef]
- Ritchie, J.T.; NeSmith, D.S. Temperature and crop development. In Modeling Plant and Soil Systems; Hanks, J., Ritchie, J.T., Eds.; Agron. Monogr. ASA; CSSA; SSSA: Madison, WI, USA, 1991; pp. 5–29. [Google Scholar]
- DWD [Deutscher Wetterdienst] 1 × 1 km Wetterdaten. Available online: http://www.dwd.de/DE/leistungen/webwerdis/webwerdis.html (accessed on 15 May 2016).
- Ernst, P.; Loeper, E.G. Temperaturentwicklung und Vegetationsbeginn auf dem Grünland. Wirtschaftseigene Futter 1976, 22, 5–11. [Google Scholar]
- BlackBridge (Ed.) Spectral Response curves of the RapidEye Sensor. Available online: http://blackbridge.com/rapideye/upload/Spectral_Response_Curves.pdf (accessed on 20 January 2016).
- Cleveland, W.S.; Grosse, E.; Shyu, W.M. Local regression models. In Statistical Models in S; Chambers, J.M., Hastie, T.J., Eds.; Chapman & Hall: Boca Raton, FL, USA, 1992; pp. 309–377. [Google Scholar]
- Wickham, H.; Chang, W. (Eds.) ggplot2. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 13 January 2017).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Rouse, J., Jr.; Haas, R.; Schell, J.; Deering, D. Monitoring vegetation systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309–317. [Google Scholar]
- Guyot, G.; Baret, F.; Major, D.J. High spectral resolution: Determination of spectral shifts between the red and the near infrared. Int. Arch. Photogramm. Remote Sens. 1988, 11, 750–760. [Google Scholar]
- Datt, B. A new reflectance index for remote sensing of chlorophyll content in higher plants: Tests using eucalyptus leaves. J. Plant Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Peñuelas, J.; Gamon, J.A.; Fredeen, A.L.; Merino, J.; Field, C.B. Reflectance indices associated with physiological changes in nitrogen- and water-limited sunflower leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Underwood, E.; Ustin, S.; Dipietro, D. Mapping nonnative plants using hyperspectral imagery. Remote Sens. Environ. 2003, 86, 150–161. [Google Scholar] [CrossRef]
- Peñuelas, J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance Water Index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Peñuelas, P.; Filella, I.; Lloret, P.; Munoz, F.; Vilajeliu, M. Reflectance assessment of mite effects on apple trees. Int. J. Remote Sens. 1995, 16, 2727–2733. [Google Scholar] [CrossRef]
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Chambers, J.M.; Hastie, T.J. (Eds.) Statistical Models in S; Chapman & Hall: Boca Raton, FL, USA, 1992.
- Stagakis, S.; Markos, N.; Sykioti, O.; Kyparissis, A. Monitoring canopy biophysical and biochemical parameters in ecosystem scale using satellite hyperspectral imagery: An application on a Phlomis fruticosa Mediterranean ecosystem using multiangular CHRIS/PROBA observations. Remote Sens. Environ. 2010, 114, 977–994. [Google Scholar] [CrossRef]
- Roelofsen, H.; van Bodegom, P.; Kooistra, L.; Witte, J.P. Trait Estimation in Herbaceous Plant Assemblages from in situ Canopy Spectra. Remote Sens. 2013, 5, 6323–6345. [Google Scholar] [CrossRef]
- Jensen, J.R. (Ed.) Remote Sensing of the Environment. An Earth Resource Perspective, 2nd ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2007.
- Rossini, M.; Cogliati, S.; Meroni, M.; Migliavacca, M.; Galvagno, M.; Busetto, L.; Cremonese, E.; Juliatta, T.; Siniscalco, C.; Morra di Cella, U.; et al. Remote sensing-based estimation of gross primary production in a subalpine grassland. Biogeosciences 2012, 9, 2565–2584. [Google Scholar] [CrossRef]
- Harmoney, K.R.; Moore, K.J.; George, J.R.; Brummer, E.C.; Russell, J.R. Determination of pasture biomass using four indirect methods. Agron. J. 1997, 89, 665–672. [Google Scholar] [CrossRef]
- Norman, J.M.; Welles, J.M.; Walter, E.A. Contrasts among bidirectional reflectance of leaves, canopies, and soils. IEEE Trans. Geosci. Remote Sens. 1985, 23, 659–667. [Google Scholar] [CrossRef]
- Sánchez-Azofeifa, G.A.; Castro, K.; Wright, S.J.; Gamon, J.; Kalacska, M.; Rivard, B.; Schnitzler, S.A.; Feng, J.L. Differences in leaf traits, leaf internal structure, and spectral reflectance between two communities of lianas and trees: Implications for remote sensing in tropical environments. Remote Sens. Environ. 2009, 113, 2076–2088. [Google Scholar] [CrossRef]
- Lorenzen, B.; Jensen, A. Reflectance of blue, green, red and near infrared radiation from wetland vegetation used in a model discriminating live and dead above ground biomass. New Phytol. 1998, 108, 345–355. [Google Scholar] [CrossRef]
- Wang, Q.; Adiku, A.; Tenhunen, J.; Granier, A. On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens. Environ. 2005, 94, 244–255. [Google Scholar] [CrossRef]
- Cho, M.A.; Skidmore, A.; Corsi, F.; Van Wieren, S.E.; Sobhan, I. Estimation of green grass/herb biomass from airborne hyperspectral imagery using spectral indices and partial least squares regression. Int. J. Appl. Earth Obs. Geoinf. 2007, 9, 414–424. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Enclona, E.A.; Ashton, M.S.; Van der Meer, B. Accuracy assessments of hyperspectral waveband performance for vegetation analysis applications. Remote Sens. Environ. 2004, 91, 354–376. [Google Scholar] [CrossRef]
- Idso, S.B.; Pinter, P.J., Jr.; Jackson, R.D.; Reginato, R.J. Estimation of grain yields by remote sensing of crop senescence rates. Remote Sens. Environ. 1980, 9, 87–91. [Google Scholar] [CrossRef]
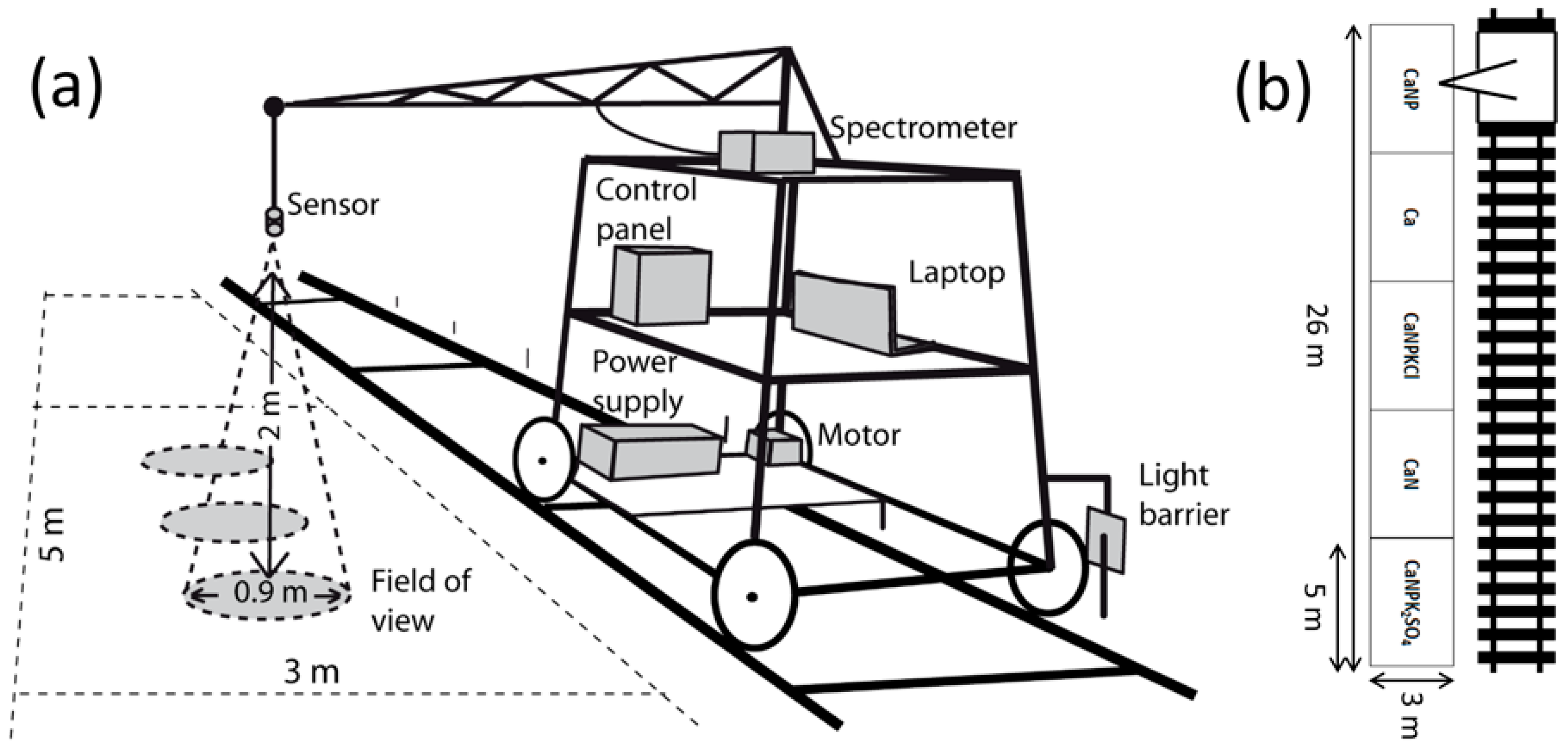
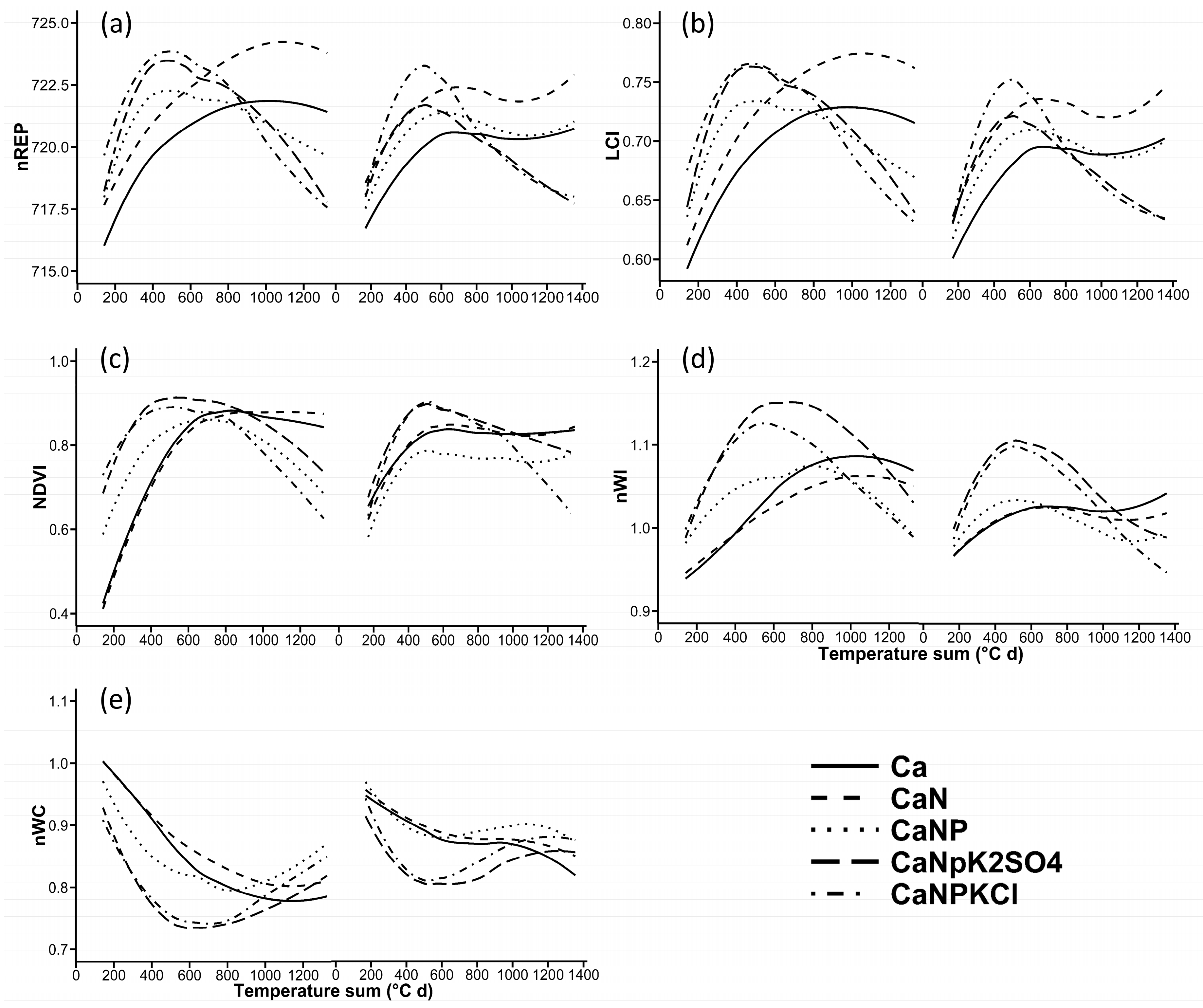
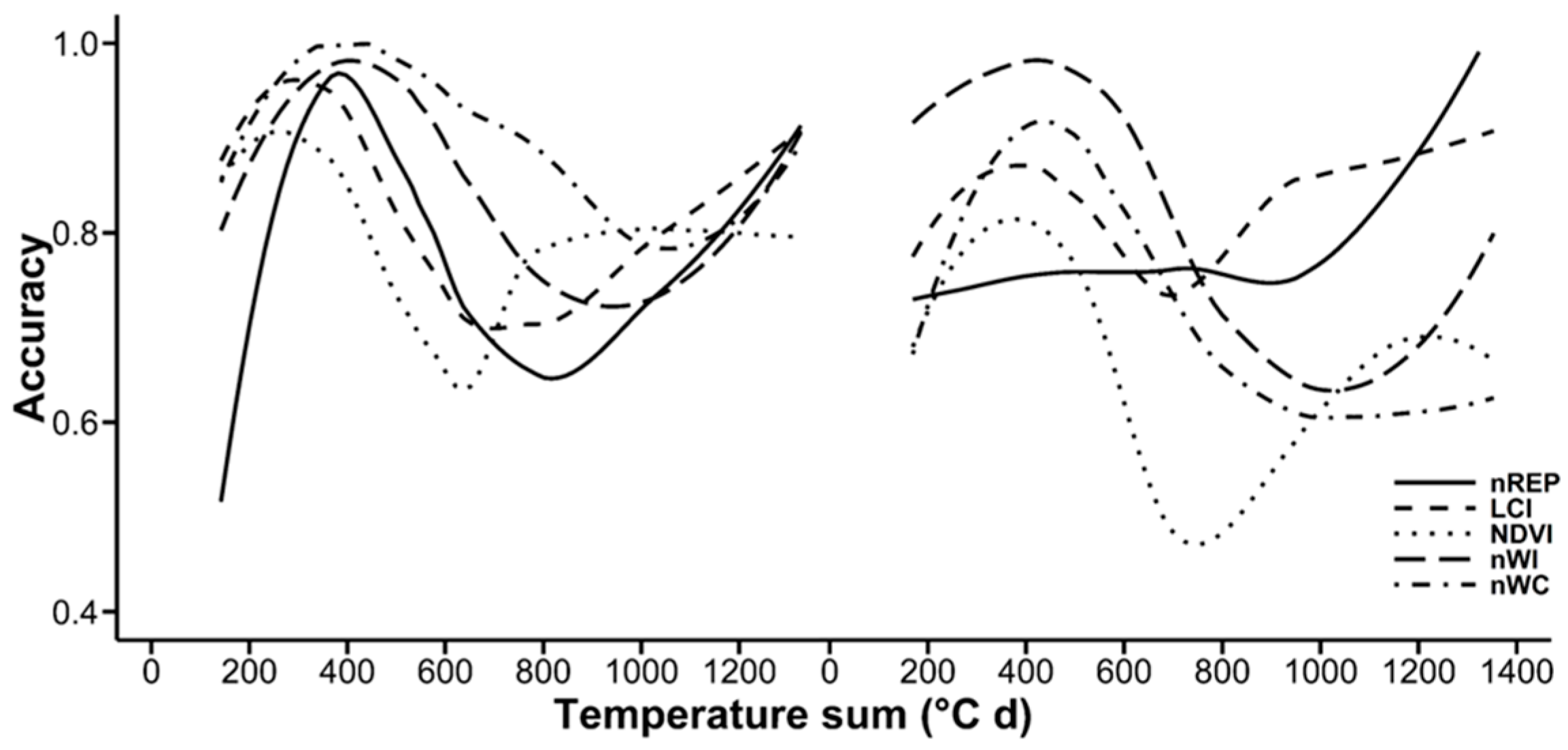
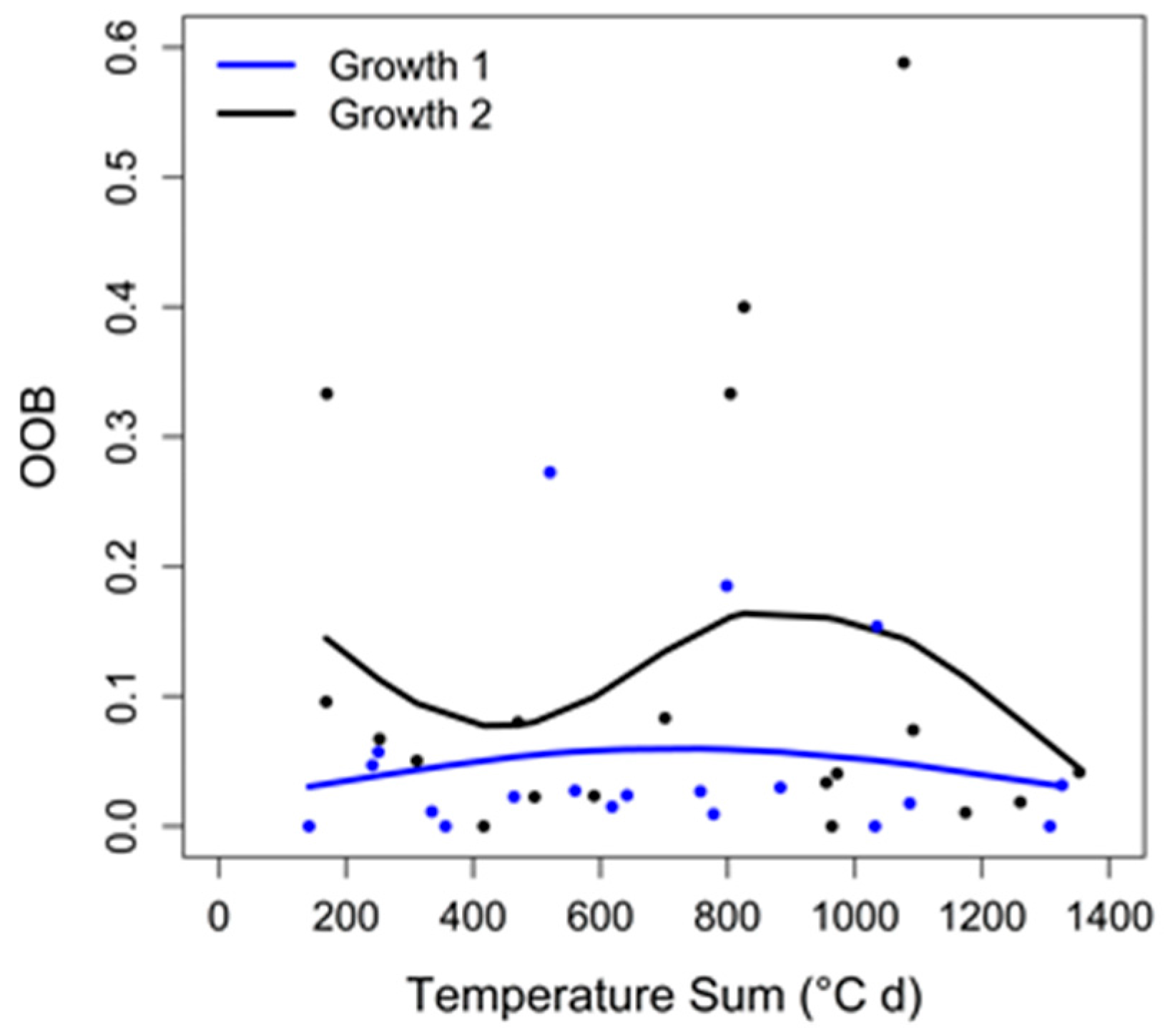
| No. of Acquisition Days | |||
|---|---|---|---|
| 2012 | 2013 | 2014 | |
| 1st growth | 4 | 7 | 8 |
| 2nd growth | 7 | 6 | 6 |
| VI | VI Full Name | Formula | Sensitivity | Source |
|---|---|---|---|---|
| GNDVI | Green Normalized Difference Vegetation Index | Green vegetation/biomass, LAI | [34] | |
| nGNDVI | narrowband Green Normalize Difference Vegetation Index | Green vegetation/biomass, LAI | [34] | |
| NDVI | Normalized Difference Vegetation Index | Green vegetation/biomass, LAI | [35] | |
| nNDVI | narrowband Normalized Difference Vegetation Index | Green vegetation/biomass, LAI | [35] | |
| nREP | narrowband Red Edge Position | Chlorophyll | [36] | |
| LCI | Leaf Chlorophyll Index | Chlorophyll | [37] | |
| nLCI | narrowband Leaf Chlorophyll Index | Chlorophyll | [37] | |
| nNPCI | narrowband Normalized Pigment Chlorophyll Index | Chlorophyll | [38] | |
| nNDLI | narrowband Normalized Difference Lignin Index | Lignin content | [39] | |
| nNDNI | narrowband Normalized Difference Nitrogen Index | Nitrogen content | [39] | |
| nPRI | narrowband Photochemical Reflectance Index | Physiology (photosynthesis, pigments) | [38] | |
| nWC | narrowband Water Content | Water content/water stress | [40] | |
| nWI | narrowband Water Index | Water content | [41] | |
| nSIPI | narrowband Structure Intensive Pigment Index | Pigments | [42] | |
| nNPQI | narrowband Normalized Phaeophytization Index | Phaeophytization | [43] |
| 1st growth | Index | nWC | nWI | nLCI | nSIPI | LCI | nNDLI | NDVI | nNDNI | |
| Accuracy | 0.91 | 0.85 | 0.84 | 0.84 | 0.83 | 0.83 | 0.80 | 0.80 | ||
| Index | nNDVI | nREP | GNDVI | nGNDVI | nNPCI | nPRI | nNPQI | Average | ||
| Accuracy | 0.79 | 0.79 | 0.76 | 0.75 | 0.73 | 0.68 | 0.41 | 0.77 | ||
| 2nd growth | Index | LCI | nWI | nREP | nSIPI | nWC | nLCI | nNDLI | nNDNI | |
| Accuracy | 0.76 | 0.75 | 0.72 | 0.71 | 0.71 | 0.67 | 0.64 | 0.64 | ||
| Index | nGNDVI | nNDVI | NDVI | GNDVI | nNPCI | nNPQI | nPRI | Average | ||
| Accuracy | 0.62 | 0.62 | 0.61 | 0.61 | 0.46 | 0.46 | 0.44 | 0.63 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hollberg, J.L.; Schellberg, J. Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices. Remote Sens. 2017, 9, 81. https://doi.org/10.3390/rs9010081
Hollberg JL, Schellberg J. Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices. Remote Sensing. 2017; 9(1):81. https://doi.org/10.3390/rs9010081
Chicago/Turabian StyleHollberg, Jens L., and Jürgen Schellberg. 2017. "Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices" Remote Sensing 9, no. 1: 81. https://doi.org/10.3390/rs9010081
APA StyleHollberg, J. L., & Schellberg, J. (2017). Distinguishing Intensity Levels of Grassland Fertilization Using Vegetation Indices. Remote Sensing, 9(1), 81. https://doi.org/10.3390/rs9010081