
Comparing the Dry Season In-Situ Leaf Area Index (LAI) Derived from High-Resolution RapidEye Imagery with MODIS LAI in a Namibian Savanna


Abstract
:
1. Introduction
- (1).
- Assess ground-based dry-season LAI in a Namibian savanna ecosystem;
- (2).
- upscale LAI field measurements to high-resolution RapidEye imagery;
- (3).
- and compare an in situ-calibrated model of LAI with the MODIS LAI product (MOD15A2) in order to evaluate its performance.
2. Study Area
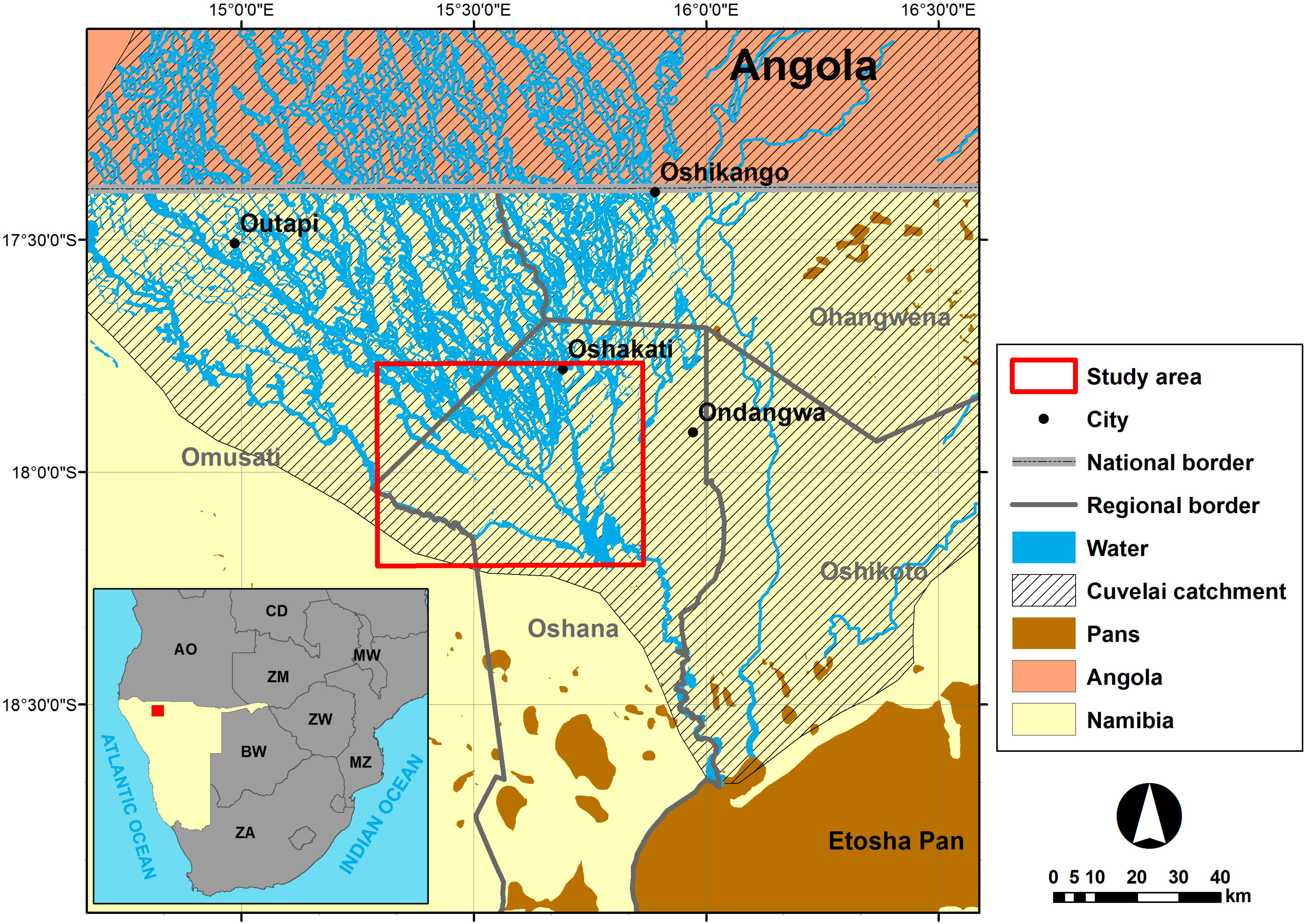
3. Materials and Methods
3.1. Remote Sensing Data
3.1.1. RapidEye
3.1.2. MODIS
3.2. Field Data
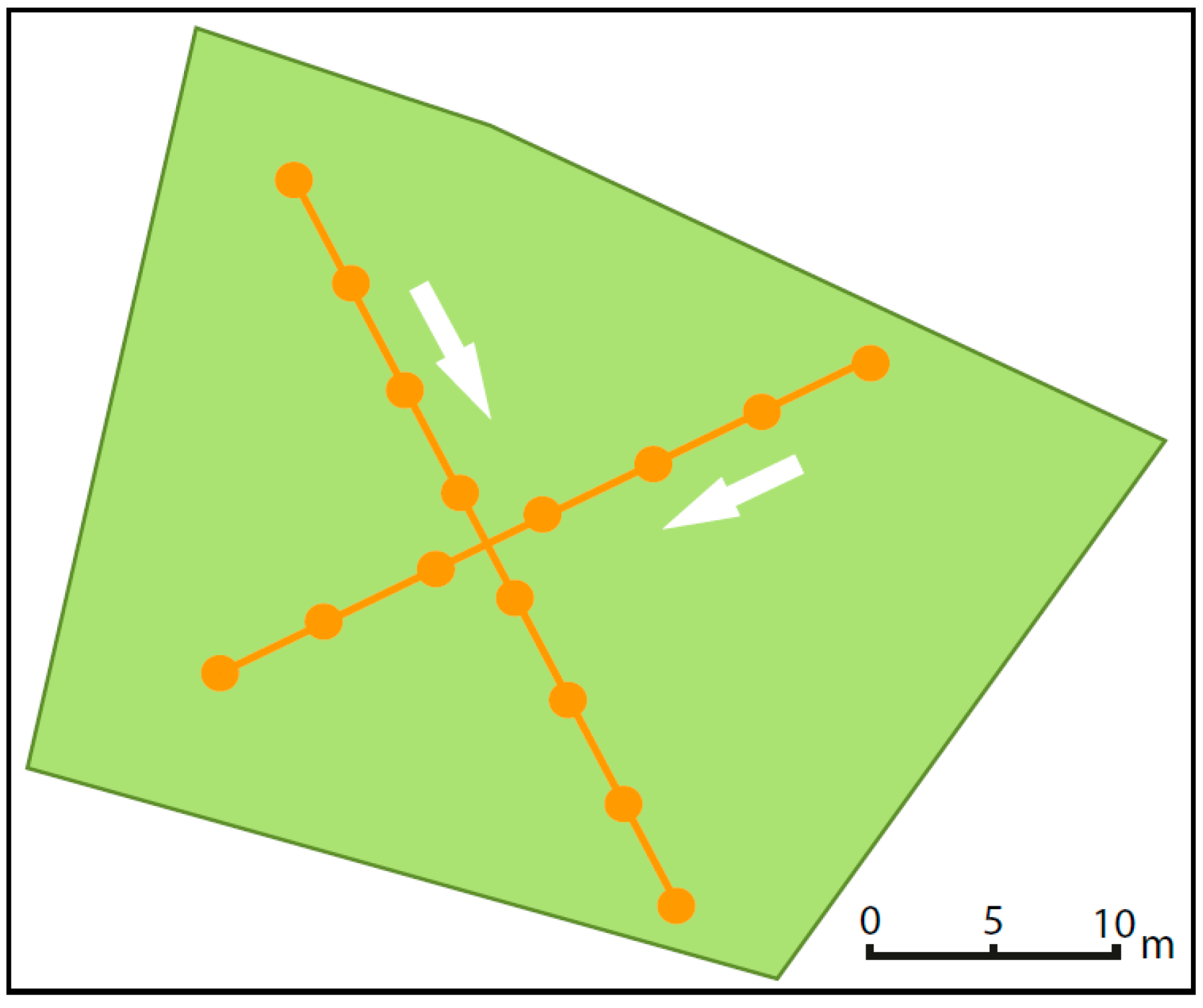
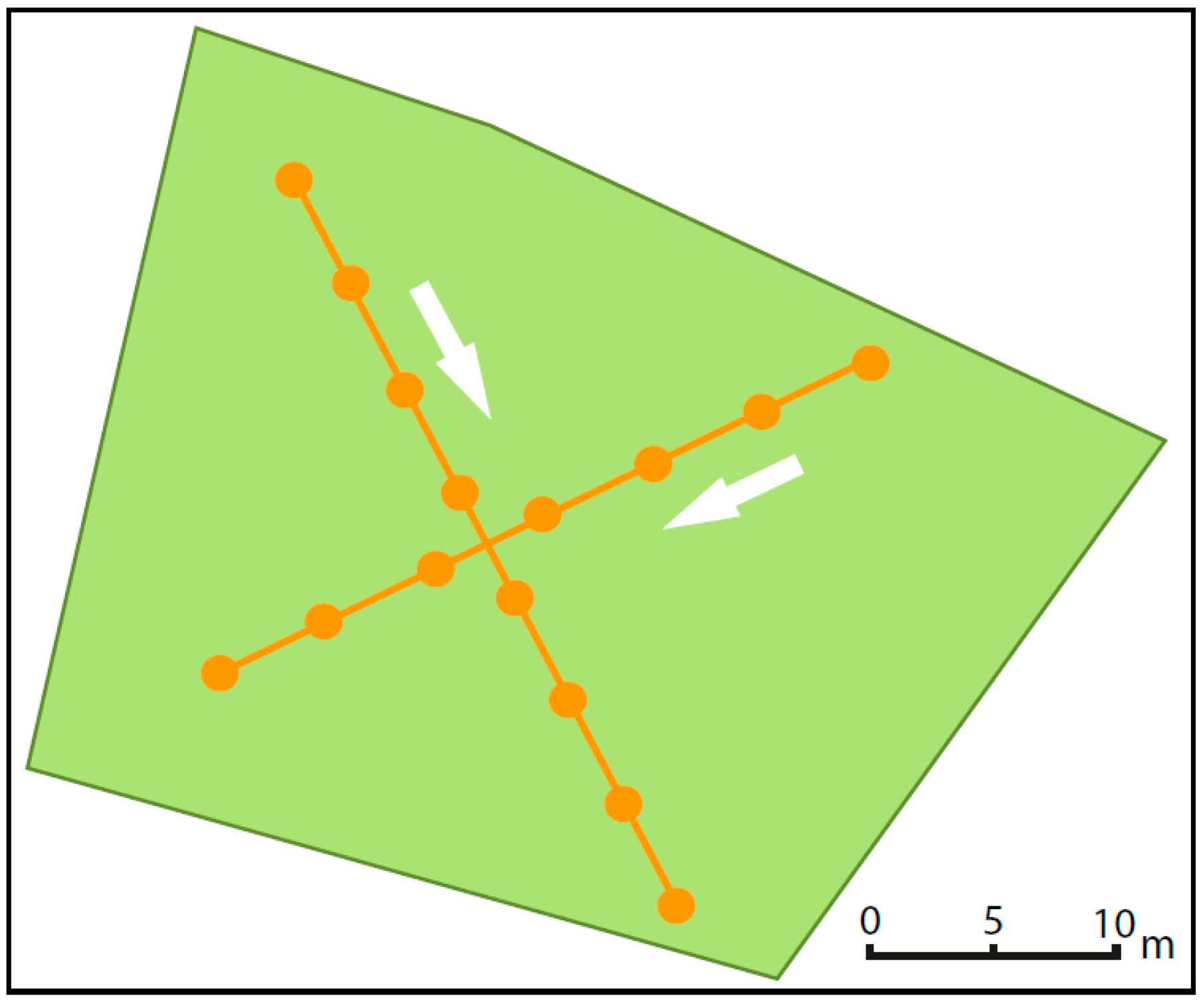
3.2.1. Site Selection
3.2.2. Recorded Parameters
3.2.3. Leaf Area Index
3.3. Empirical-Statistical Modeling of LAI

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral Vegetation Index | Equation | Reference |
|---|---|---|
| Simple Ratio (SR) | [50] | |
| Difference Vegetation Index (DVI) | [51] | |
| Normalized Difference Vegetation Index (NDVI) | [52] | |
| Transformed Vegetation Index (TVI) | [53] | |
| Soil-adjusted Vegetation Index (SAVI) | [49] | |
| Enhanced Vegetation Index (EVI) | [49] | |
| Simple Ratio (SR NIR/RE) | [54] | |
| Simple Ratio (SR RE/R) | [54] | |
| Normalized Difference Vegetation Index (NDVI NIR/RE) | [54] | |
| Normalized Difference Vegetation Index (NDVI RE/R) | - | |
| NOTE: SL = 0.1, L = 1.0, C1 = 6.0, and C2 = 7.5 | - | - |

3.4. Comparing the Empirical Model with MODIS LAI (MOD15A2)
- (1).
- the Coefficient of determination (R2) to specify the proportion of variance between two models explained by the predictor variable;
- (2).
- the Root-Mean-Square-Error (RMSE), which is calculated using:where is the predictor variable (i.e., the aggregated LAImodel), is the estimated variable (i.e., MOD15A2), and is the sample size (i.e., the number of pixels);
- (3).
- the Relative Predictive Error (RPE), which provides a directional measure from mean difference between and in percent and is defined as:with: , the mean of and , the mean of ;
- (4).
- the Modified Index of Agreement (mIOA) [58]:
4. Results
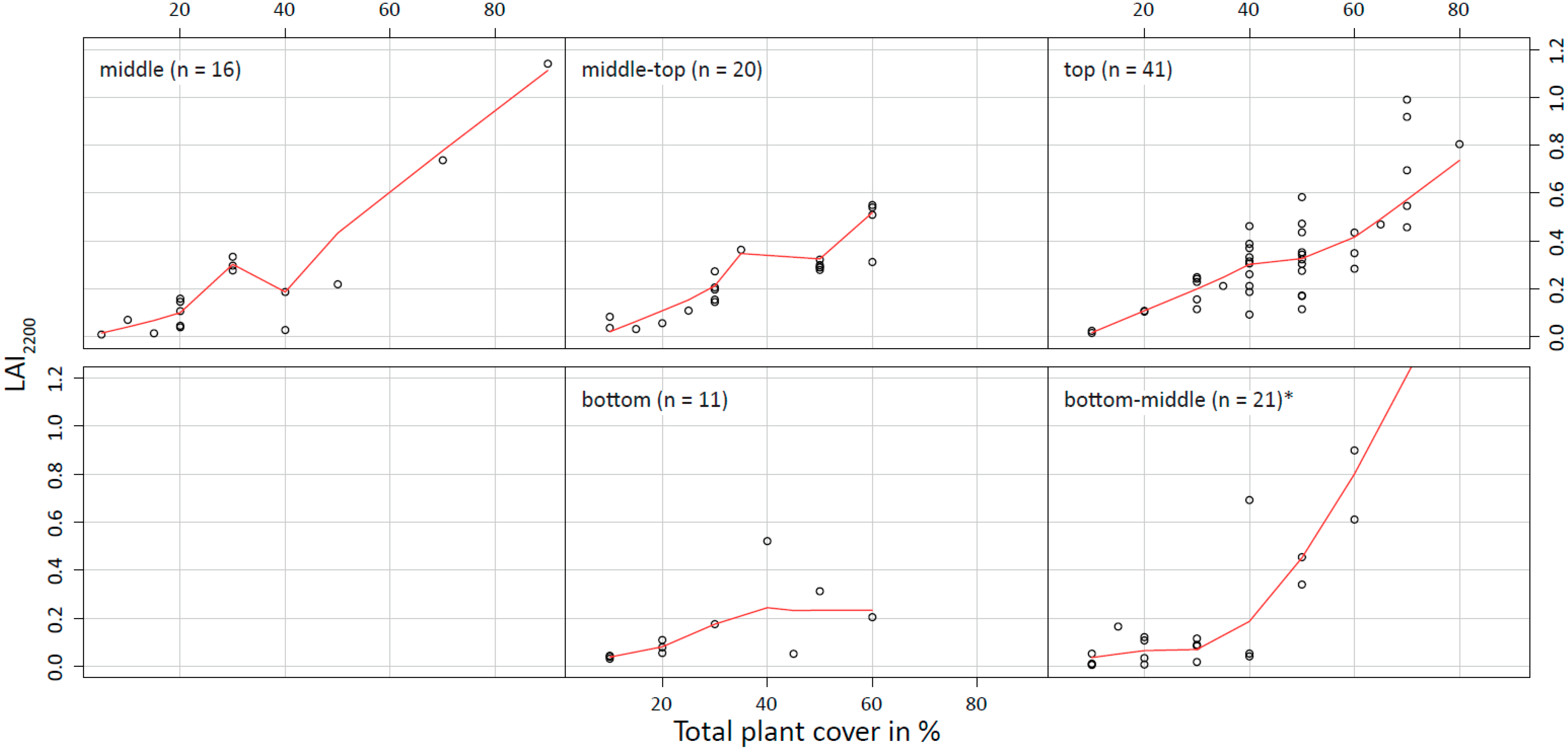
4.1. Field Data
| FLU | Description | Characteristic Species | Total Plant Cover (%) | Predominant Terrain Position | No. of ESUs |
|---|---|---|---|---|---|
| open woodland | tree cover (>5 m) >30% or trees are dominant | H. petersiana | 30–90 | middle-top | 9 |
| wooded shrub land | shrub cover >30% | Acacia arenaria, Acacia hebeclada tr. | 50–60 | top | 8 |
| P.-L. leubnitziae shrub land | shrub cover >30% | Pechuel-Loschea leubnitziae | 40–70 | top | 8 |
| C. mopane shrub land | shrub cover >30% | C. mopane | 40–70 | middle-top | 8 |
| grassland tufts | grassland at low elevation, tuft forming species | S. iocladus, Eragrostis lehmanniana | 10–40 | bottom-middle | 7 |
| grassland medium | grassland at medium elevation | A. stipoides, W. sarmentosa | 5–30 | middle | 11 |
| grassland high | grassland at high elevation | A. stipoides, O. paucinervis | 40–70 | top | 8 |
| shrub-wooded grassland | grassland with shrub cover <30% | A. stipoides, O. paucinervis, A arenaria, A. hebeclada tr. | 20–80 | middle-top | 11 |
| P.-L. leubnitziae grassland | grassland with shrub cover <30% | A. stipoides, O. paucinervis, P-L. leubnitziae | 20–50 | top | 7 |
| seasonally flooded grassland | grassland in minor depressions, tall-growing species vs. intense grazing | L. fusca, S. iocladus, Elytrophorus globularis | 10–90 | bottom-middle | 16 |
| R. limnophila forbs | R. limnophila dominant | R. limnophila | 10–20 | bottom-middle | 8 |
| agricultural land | remains of Pennisetum glaucum | - | 10 | top | 4 |
| wetland | co-existence of vegetation, water surface and bare soil | - | 40–60 | bottom-middle | 4 |
4.2. In Situ LAI (LAI2200)
| LAI2200 | SEL | ACF | No. of Samples | LAIeff | ||
|---|---|---|---|---|---|---|
| Overall (n = 109) | Mean | 0.28 | 0.05 | 0.90 | 19 | 0.24 |
| Min. | 0.01 | 0.00 | 0.63 | 14 | 0.01 | |
| Max. | 2.09 | 0.26 | 0.99 | 33 | 1.65 | |
| Median | 0.21 | 0.04 | 0.93 | 19 | 0.20 | |
| Green vegetation only (n = 17) | Mean | 0.47 | 0.10 | 0.83 | 18 | 0.37 |
| Min. | 0.22 | 0.04 | 0.66 | 15 | 0.20 | |
| Max. | 1.14 | 0.26 | 0.94 | 25 | 0.75 | |
| Median | 0.44 | 0.09 | 0.87 | 18 | 0.33 |

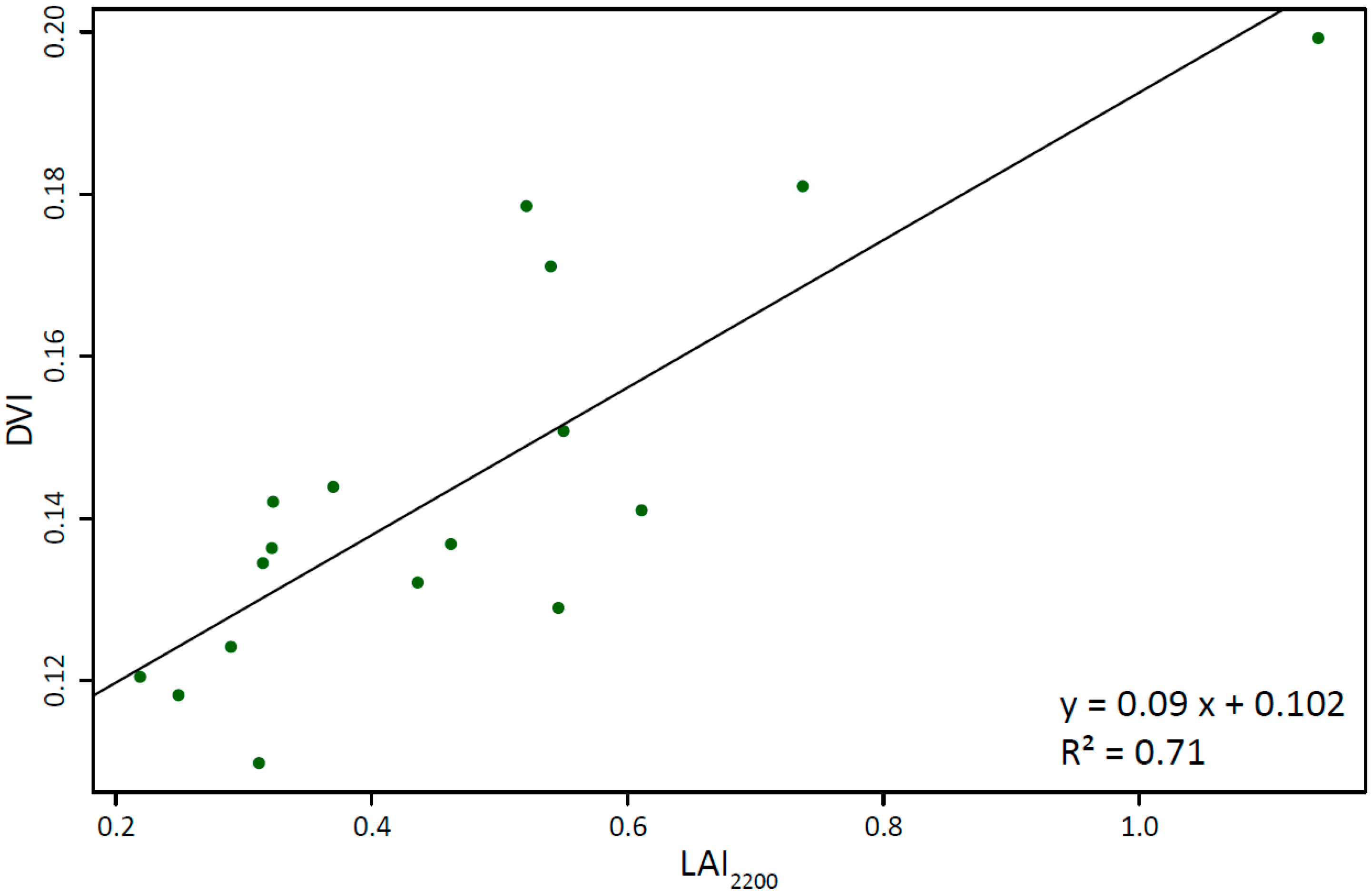
4.3. Empirical-Statistical Modeling
| SVI | SR | SRNIR/RE | SRRE/R | DVI | NDVI | NDVINIR/RE | NDVIRE/R | TVI | EVI | SAVI |
|---|---|---|---|---|---|---|---|---|---|---|
| R2 | 0.48 * | 0.55 * | 0.37 * | 0.71 * | 0.49 * | 0.56 * | 0.36 ** | 0.49 * | 0.32 ** | 0.57 * |
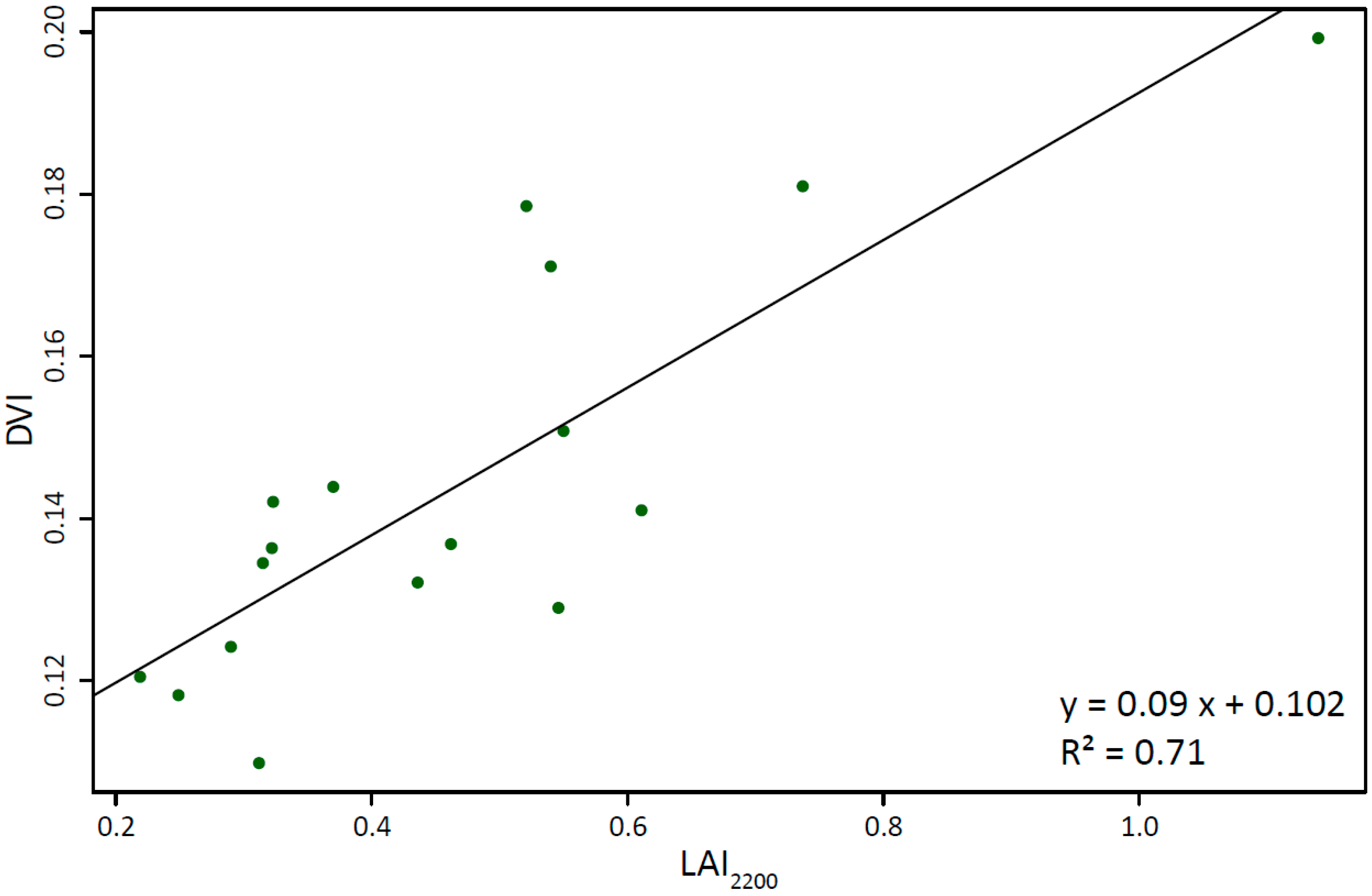
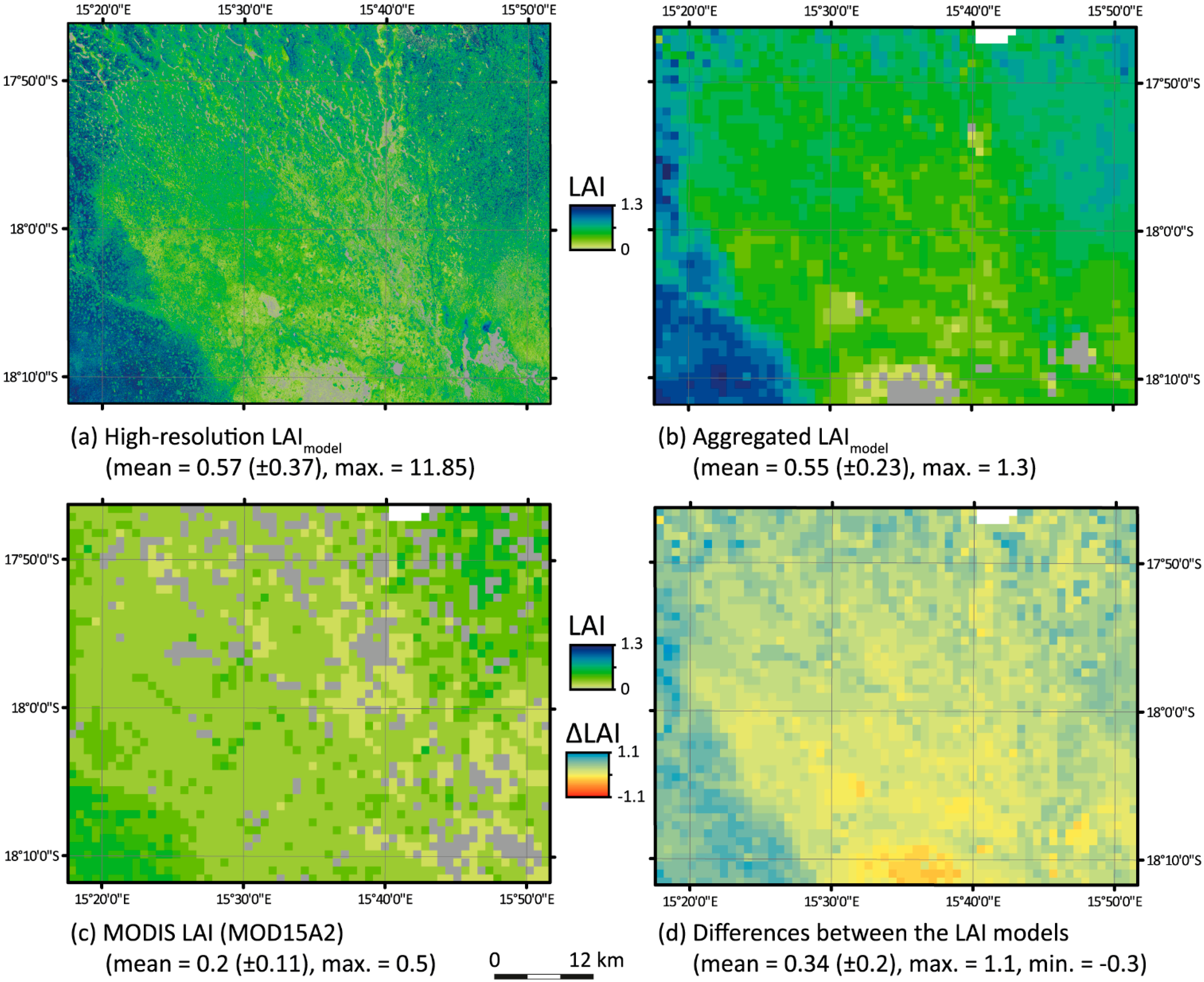
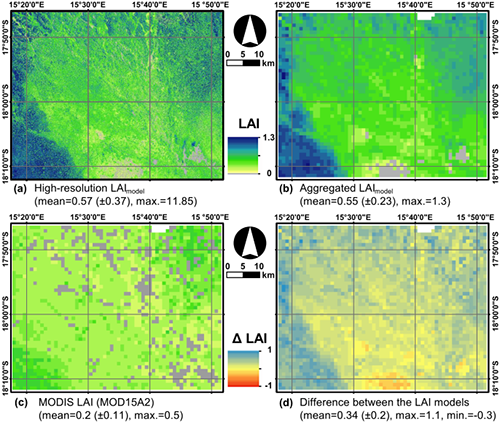
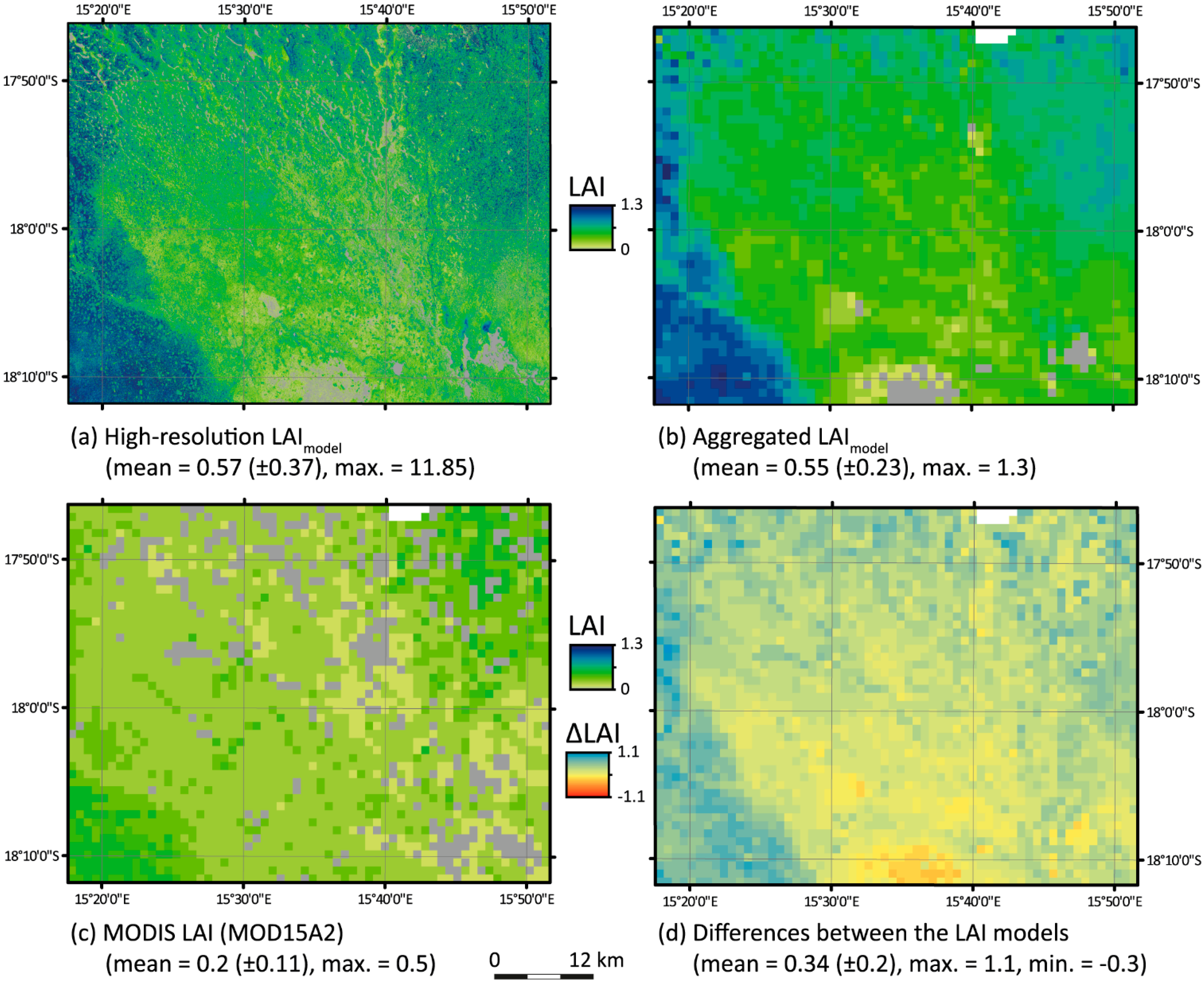
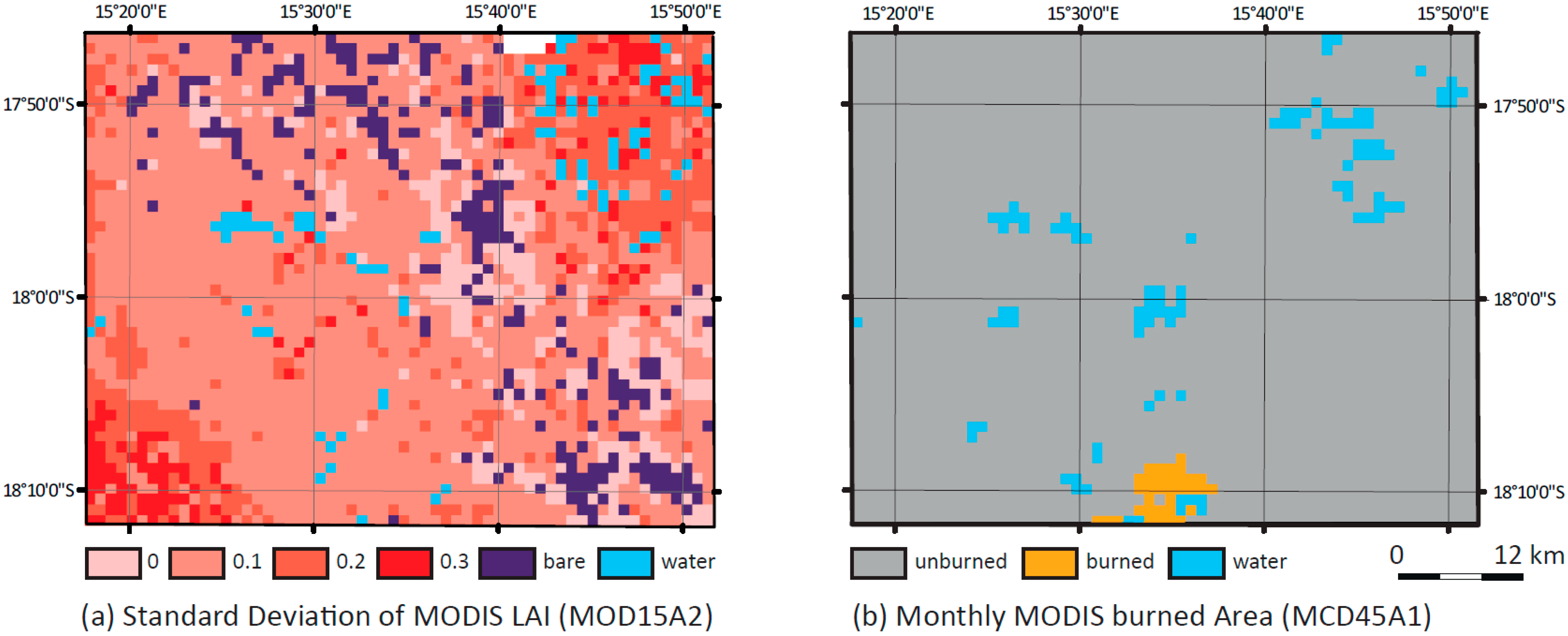
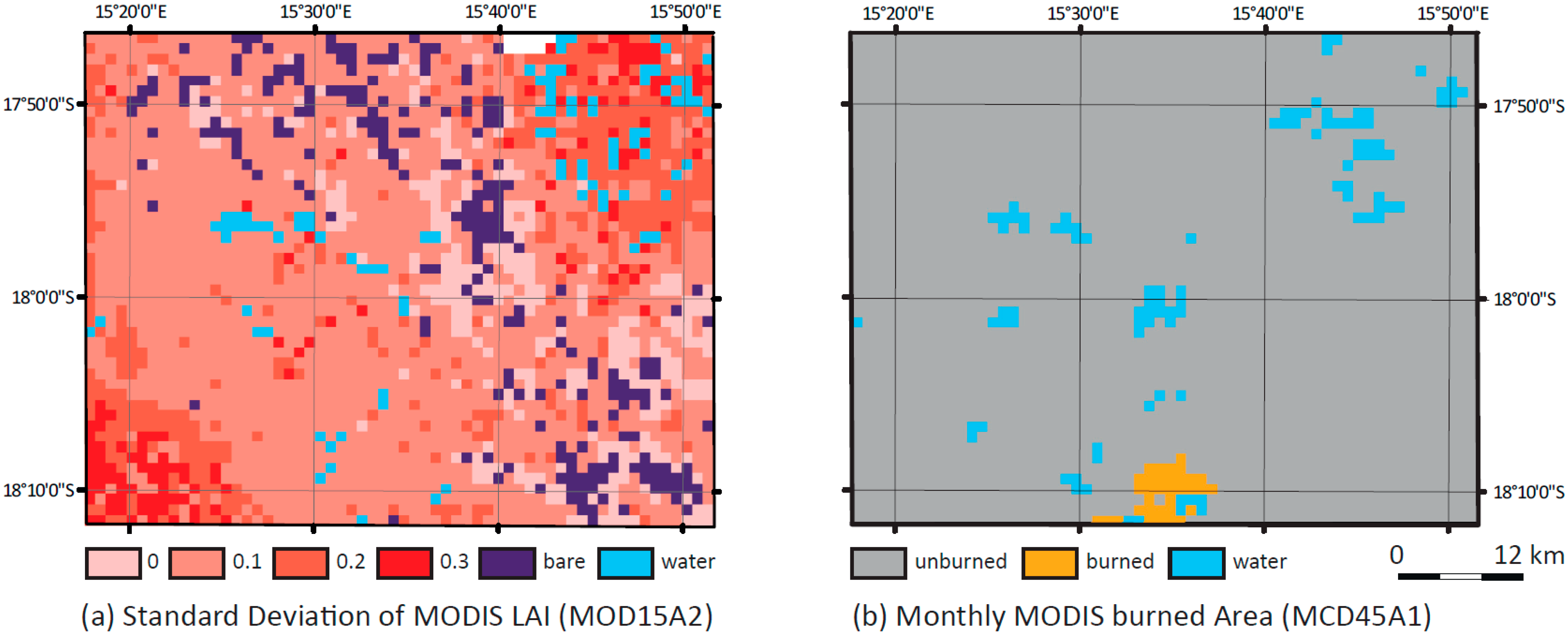
4.4. Comparison of the High-Resolution LAImodel with MOD15A2
| x | Pixels Valid (n) | Linear Model | R2 | RMSE | RPE | mIOA |
|---|---|---|---|---|---|---|
| MODIS LAI | 2811 | y = 0.8463x + 0.371 | 0.182 | 0.40 | −62.97% | 0.13 |
| MODIS LAI > 0 | 2448 | y = 1.3396x + 0.2521 | 0.293 | 0.39 | −59.18% | 0.42 |
5. Discussion
5.1. Sampling
5.2. In Situ LAI
5.3. Empirical-Statistical Modeling
5.4. Comparison with MOD15A2
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Seghieri, J.; Floret, C.; Pontanier, R. Plant phenology in relation to water availability: Herbaceous and woody species in the savannas of northern Cameroon. J. Trop. Ecol. 1995, 11, 237–254. [Google Scholar] [CrossRef]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in Savannas. Ann. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Wagenseil, H.; Samimi, C. Woody vegetation cover in Namibian savannahs: A modelling approach based on remote sensing. Erdkunde 2007, 61, 325–334. [Google Scholar] [CrossRef]
- Barclay, H.J. Conversion of total leaf area to projected leaf area in lodgepole pine and Douglas-fir. Tree Physiol. 1998, 18, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Baret, F.; Smith, G.J.; Jonckheere, I.; Coppin, P. Review of methods for in situ leaf area index (LAI) determination Part II. Estimation of LAI, errors and sampling. Agric. For. Meteorol. 2004, 121, 37–53. [Google Scholar] [CrossRef]
- Garrigues, S.; Shabanov, N.V.; Swanson, K.; Morisette, J.T.; Baret, F.; Myneni, R.B. Intercomparison and sensitivity analysis of Leaf Area Index retrievals from LAI-2000, AccuPAR, and digital hemispherical photography over croplands. Agric. For. Meteorol. 2008, 148, 1193–1209. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.M.; Cihlar, J.; Park, W.M. A process-based boreal ecosystem productivity simulator using remote sensing inputs. Remote Sens. Environ. 1997, 62, 158–175. [Google Scholar] [CrossRef]
- Novick, K.A.; Stoy, P.C.; Katul, G.G.; Ellsworth, D.S.; Siqueira, M.B.S.; Juang, J.; Oren, R. Carbon dioxide and water vapor exchange in a warm temperate grassland. Oecologia 2004, 138, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Chen, X.; Ju, W.; Geng, X. Distributed hydrological model for mapping evapotranspiration using remote sensing inputs. J. Hydrol. 2005, 305, 15–39. [Google Scholar] [CrossRef]
- Boulain, N.; Cappelaere, B.; Ramier, D.; Issoufou, H.B.A.; Halilou, O.; Seghieri, J.; Guillemin, F.; Oï, M.; Gignoux, J.; Timouk, F. Towards an understanding of coupled physical and biological processes in the cultivated Sahel-2. Vegetation and carbon dynamics. J. Hydrol. 2009, 375, 190–203. [Google Scholar] [CrossRef]
- Kraus, T. Ground-based Validation of the MODIS Leaf Area Index Product for East African Rain Forest Ecosystems. Ph.D. Thesis, Friedrich-Alexander University, Erlangen-Nuremberg, Germany, 10 October 2008. [Google Scholar]
- Bréda, N.J.J. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. Journal Exp. Bot. 2003, 54, 2403–2417. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Welles, J.M.; Cohen, S. Canopy structure measurement by gap fraction analysis using commercial instrumentation. J. Exp. Bot. 1996, 47, 1335–1342. [Google Scholar] [CrossRef]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and indirect estimation of leaf area index, f(APAR), and net primary production of terrestrial ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Van Gardingen, P.R.; Jackson, G.E.; Hernandez-Daumas, S.; Russell, G.; Sharp, L. Leaf area index estimates obtained for clumped canopies using hemispherical photography. Agric. For. Meteorol. 1999, 94, 243–257. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T.A. Defining leaf area index for non-flat leaves. Plant. Cell. Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Asner, G.P.; Scurlock, J.M.O.; Hicke, J.A. Global synthesis of leaf area index observations: Implications for ecological and remote sensing studies. Glob. Ecol. Biogeogr. 2003, 12, 191–205. [Google Scholar] [CrossRef]
- Dorigo, W.A.; Zurita-Milla, R.; de Wit, A.J.W.; Brazile, J.; Singh, R.; Schaepman, M.E. A review on reflective remote sensing and data assimilation techniques for enhanced agroecosystem modeling. Int. J. Appl. Earth Obs. Geoinf. 2007, 9, 165–193. [Google Scholar] [CrossRef]
- Qi, J.; Kerr, Y.H.; Moran, M.S.; Weltz, M.; Huete, A.R.; Sorooshian, S.; Bryant, R. Leaf area index estimates using remotely sensed data and BRDF models in a semiarid region. Remote Sens. Environ. 2000, 73, 18–30. [Google Scholar] [CrossRef]
- Privette, J.L.; Myneni, R.B.; Knyazikhin, Y.; Mukelabai, M.; Roberts, G.; Tian, Y.; Wang, Y.; Leblanc, S.G. Early spatial and temporal validation of MODIS LAI product in the Southern Africa Kalahari. Remote Sens. Environ. 2002, 83, 232–243. [Google Scholar] [CrossRef]
- Tian, Y.; Woodcock, C.E.; Wang, Y.; Privette, J.L.; Shabanov, N.V.; Zhou, L.; Zhang, Y.; Buermann, W.; Dong, J.; Veikkanen, B.; et al. Multiscale analysis and validation of the MODIS LAI product I. Uncertainty assessment. Remote Sens. Environ. 2002, 83, 414–430. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.S. Evaluation of MODIS LAI, fAPAR and the relation between fAPAR and NDVI in a semi-arid environment using in situ measurements. Remote Sens. Environ. 2004, 91, 490–507. [Google Scholar] [CrossRef]
- Hill, M.J.; Senarath, U.; Lee, A.; Zeppel, M.; Nightingale, J.M.; Williams, R.J.; McVicar, T.R. Assessment of the MODIS LAI product for Australian ecosystems. Remote Sens. Environ. 2006, 101, 495–518. [Google Scholar] [CrossRef]
- Kraus, T.; Schmidt, M.; Dech, S.W.; Samimi, C. The potential of optical high resolution data for the assessment of leaf area index in East African rainforest ecosystems. Int. J. Remote Sens. 2009, 30, 5039–5059. [Google Scholar] [CrossRef]
- Serbin, S.P.; Ahl, D.E.; Gower, S.T. Spatial and temporal validation of the MODIS LAI and FPAR products across a boreal forest wildfire chronosequence. Remote Sens. Environ. 2013, 133, 71–84. [Google Scholar] [CrossRef]
- Ryu, Y.; Sonnentag, O.; Nilson, T.; Vargas, R.; Kobayashi, H.; Wenk, R.; Baldocchi, D.D. How to quantify tree leaf area index in an open savanna ecosystem: A multi-instrument and multi-model approach. Agric. For. Meteorol. 2010, 150, 63–76. [Google Scholar] [CrossRef]
- Okin, G.S. The contribution of brown vegetation to vegetation dynamics. Ecology 2010, 91, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Bond, W.J.; van Wilgen, B.W. Fire and Plants, 1st ed.; Chapman & Hall: London, UK, 1996. [Google Scholar]
- Mendelsohn, J.M.; El Obeid, S.; Roberts, C. A Profile of North-Central Namibia; Gamsberg Macmillan Publishers: Windhoek, Namibia, 2000. [Google Scholar]
- Kluge, T.; Liehr, S.; Lux, A.; Moser, P.; Niemann, S.; Umlauf, N.; Urban, W. IWRM concept for the Cuvelai Basin in northern Namibia. Phys. Chem. Earth 2008, 33, 48–55. [Google Scholar] [CrossRef]
- Mendelsohn, J.M.; Jarvis, A.; Roberts, C.; Robertson, T. Atlas of Namibia: A Portrait of the Land and Its People; David Philip: Cape Town, South Africa, 2002. [Google Scholar]
- RapidEye AG. In RapidEye Satellite Imagery Product Specifications. Version 3.2.; RapidEye AG: Brandenburg An der Havel, Germany, 2011.
- Song, C.; Woodcock, C.E.; Seto, K.C.; Lenney, M.P.; Macomber, S.A. Classification and change detection using Landsat TM data: When and how to correct atmospheric effects? Remote Sens. Environ. 2001, 75, 230–244. [Google Scholar] [CrossRef]
- Mather, P.M.; Koch, M. Computer Processing of Remotely-Sensed Images: An. Introduction, 4th ed.; Wiley-Blackwell: Chichester, UK, 2011. [Google Scholar]
- Yang, W.; Shabanov, N.V.; Huang, D.; Wang, W.; Dickinson, R.E.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Analysis of leaf area index products from combination of MODIS Terra and Aqua data. Remote Sens. Environ. 2006, 104, 297–312. [Google Scholar] [CrossRef]
- Shabanov, N.V.; Huang, D.; Yang, W.; Tan, B.; Knyazikhin, Y.; Myneni, R.B.; Ahl, D.E.; Gower, S.T.; Huete, A.R.; Aragão, L.E.O.C.; et al. Analysis and optimization of the MODIS leaf area index algorithm retrievals over broadleaf forests. IEEE Trans. Geosci. Remote Sens. 2005, 43, 1855–1865. [Google Scholar] [CrossRef]
- Fang, H.; Jiang, C.; Li, W.; Wei, S.; Baret, F.; Chen, J.M.; Garcia-Haro, J.; Liang, S.; Liu, R.; Myneni, R.B.; et al. Characterization and intercomparison of global moderate resolution leaf area index (LAI) products: Analysis of climatologies and theoretical uncertainties. J. Geophys. Res.: Biogeosci. 2013, 118, 529–548. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Giladi, I.; Ziv, Y.; May, F.; Jeltsch, F. Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem. J. Veg. Sci. 2011, 22, 983–996. [Google Scholar] [CrossRef]
- LI-COR Inc. LAI-2200 Plant. Canopy Analyzer Instruction Manual; Li-Cor Inc.: Lincoln, NE, USA, 2009. [Google Scholar]
- Nilson, T. A theoretical analysis of the frequency of gaps in plant stands. Agr. Meteorol. 1971, 8, 25–38. [Google Scholar] [CrossRef]
- Jones, H.G.; Vaughan, R.A. Remote Sensing of Vegetation: Principles, Techniques, and Applications; Oxford University Press: Oxford, UK and New York, NY, USA, 2010. [Google Scholar]
- Kobayashi, H.; Ryu, Y.; Baldocchi, D.D.; Welles, J.M.; Norman, J.M. On the correct estimation of gap fraction: How to remove scattered radiation in gap fraction measurements? Agric. For. Meteorol. 2013, 174–175, 170–183. [Google Scholar] [CrossRef]
- White, M.A.; Asner, G.P.; Nemani, R.R.; Privette, J.L.; Running, S.W. Measuring fractional cover and leaf area index in arid ecosystems: Digital camera, radiation transmittance, and laser altimetry methods. Remote Sens. Environ. 2000, 74, 45–57. [Google Scholar] [CrossRef]
- Garrigues, S.; Allard, D.; Weiss, M.; Baret, F. Comparing VALERI Sampling Schemes to Better Represent High Spatial Resolution Satellite Pixel from Ground Measurements: How to Characterize an ESU. Available online: http://www.avignon.inra.fr/valeri/methodology/samplingschemes.pdf (accessed on 25 January 2015).
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Huete, A.R.; Justice, C.; van Leeuwen, W. MODIS Vegetation Index (MOD 13)—Algorithm Theoretical Basis Document (Version 3). Available online: http://modis.gsfc.nasa.gov/data/atbd/atbd_mod13.pdf (accessed on 25 January 2015).
- Birth, G.S.; McVey, G.R. Measuring the color of growing turf with a reflectance spectrophotometer. Agron. J. 1968, 60, 640–643. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Rouse, J.W., Jr.; Haas, R.H.; Deering, D.W.; Schell, J.A. Monitoring the Vernal Advancement and Retrogradiation (Green Wave Effect) of Natural Vegetation. Available online: http://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/19740004927.pdf (accessed on 10 April 2015).
- Deering, D.W.; Haas, R.H. Using Landsat Digital Data for Estimating Green Biomass. Available online: http://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/19800024311.pdf (accessed on 25 January 2015).
- Ehammer, A.; Fritsch, S.; Conrad, C.; Lamers, J.; Dech, S. Statistical derivation of fPAR and LAI for irrigated cotton and rice in arid Uzbekistan by combining multi-temporal RapidEye data and ground measurements. Proc. SPIE 2010, 782409. [Google Scholar] [CrossRef]
- Morisette, J.T.; Baret, F.; Privette, J.L.; Myneni, R.B.; Nickeson, J.E.; Garrigues, S.; Shabanov, N.V.; Weiss, M.; Fernandes, R.A.; Leblanc, S.G.; et al. Validation of global moderate-resolution LAI products: A framework proposed within the CEOS land product validation subgroup. IEEE Trans. Geosci. Remote Sens. 2006, 44, 1804–1814. [Google Scholar] [CrossRef]
- Yang, W.; Tan, B.; Huang, D.; Rautiainen, M.; Shabanov, N.V.; Wang, Y.; Privette, J.L.; Huemmrich, K.F.; Fensholt, R.; Sandholt, I.; et al. MODIS leaf area index products: From validation to algorithm improvement. IEEE Trans. Geosci. Remote Sens. 2006, 44, 1885–1896. [Google Scholar] [CrossRef]
- Kanniah, K.D.; Beringer, J.; Hutley, L.B.; Tapper, N.J.; Zhu, X. Evaluation of Collections 4 and 5 of the MODIS Gross Primary Productivity product and algorithm improvement at a tropical savanna site in northern Australia. Remote Sens. Environ. 2009, 113, 1808–1822. [Google Scholar] [CrossRef]
- Willmott, C.J.; Ackleson, S.G.; Davis, R.E.; Feddema, J.J.; Klink, K.M.; Legates, D.R.; O’Donnell, J.; Rowe, C.M. Statistics for the evaluation and comparison of models. J. Geophys. Res. 1985, 90, 8995–9005. [Google Scholar] [CrossRef]
- Willmott, C.J.; Robeson, S.M.; Matsuura, K. A refined index of model performance. Int. J. Climatol. 2012, 32, 2088–2094. [Google Scholar] [CrossRef]
- Zandler, H.; Brenning, A.; Samimi, C. Quantifying dwarf shrub biomass in an arid environment: Comparing empirical methods in a high dimensional setting. Remote Sens. Environ. 2015, 158, 140–155. [Google Scholar] [CrossRef]
- Law, B.E.; Campbell, J.L.; Chen, J.M.; Sun, O.; Schwartz, M.; van Ingen, C.; Verma, S. Terrestrial Carbon Observations: Protocols for Vegetation Sampling and Data Submission, Report of the Global Terrestrial Observing System (GTOS). Available online: http://www.fao.org/gtos/doc/pub55.pdf (accessed on 10 April,2015).
- Zhang, J.; Zhang, C. Sampling and sampling strategies for environmental analysis. Int. J. Environ. Anal. Chem. 2012, 92, 466–478. [Google Scholar] [CrossRef]
- Roleček, J.; Chytrý, M.; Hájek, M.; Lvončík, S.; Tichý, L. Sampling design in large-scale vegetation studies: Do not sacrifice ecological thinking to statistical purism! Folia Geobot. 2007, 42, 199–208. [Google Scholar] [CrossRef]
- Scholes, R.J.; Frost, P.G.H.; Tian, Y. Canopy structure in savannas along a moisture gradient on Kalahari sands. Glob. Chang. Biol. 2004, 10, 292–302. [Google Scholar] [CrossRef]
- Ribeiro, N.S.; Saatchi, S.S.; Shugart, H.H.; Washington-Allen, R.A. Aboveground biomass and leaf area index (LAI) mapping for Niassa Reserve, northern Mozambique. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Sea, W.B.; Choler, P.; Beringer, J.; Weinmann, R.A.; Hutley, L.B.; Leuning, R. Documenting improvement in leaf area index estimates from MODIS using hemispherical photos for Australian savannas. Agric. For. Meteorol. 2011, 151, 1453–1461. [Google Scholar] [CrossRef]
- Hill, M.J.; Román, M.O.; Schaaf, C.B.; Hutley, L.; Brannstrom, C.; Etter, A.; Hanan, N.P. Characterizing vegetation cover in global savannas with an annual foliage clumping index derived from the MODIS BRDF product. Remote Sens. Environ. 2011, 115, 2008–2024. [Google Scholar] [CrossRef]
- Ryu, Y.; Nilson, T.; Kobayashi, H.; Sonnentag, O.; Law, B.E.; Baldocchi, D.D. On the correct estimation of effective leaf area index: Does it reveal information on clumping effects? Agric. For. Meteorol. 2010, 150, 463–472. [Google Scholar] [CrossRef]
- Meza-Díaz, B.; Blackburn, G.A. Remote sensing of mangrove biophysical properties: Evidence from a laboratory simulation of the possible effects of background variation on spectral vegetation indices. Int. J. Remote Sens. 2003, 24, 53–73. [Google Scholar] [CrossRef]
- Van Leeuwen, W.; Huete, A.R. Effects of standing litter on the biophysical interpretation of plant canopies with spectral indices. Remote Sens. Environ. 1996, 55, 123–138. [Google Scholar] [CrossRef]
- Pinty, B.; Lavergne, T.; Widlowski, J.-L.; Gobron, N.; Verstraete, M.M. On the need to observe vegetation canopies in the near-infrared to estimate visible light absorption. Remote Sens. Environ. 2009, 113, 10–23. [Google Scholar] [CrossRef]
- Marsett, R.C.; Qi, J.; Heilman, P.; Biedenbender, S.H.; Watson, M.C.; Amer, S.; Weltz, M.; Goodrich, D.; Marsett, R. Remote sensing for grassland management in the arid Southwest. Rangeland Ecol. Manage. 2006, 59, 530–540. [Google Scholar] [CrossRef]
- Guerschman, J.P.; Hill, M.J.; Renzullo, L.J.; Barrett, D.J.; Marks, A.S.; Botha, E.J. Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and bare soil in the Australian tropical savanna region upscaling the EO-1 Hyperion and MODIS sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
- Fang, H.; Li, W.; Myneni, R.B. The impact of potential land cover misclassification on MODIS Leaf Area Index (LAI) estimation: A statistical perspective. Remote Sens. 2013, 5, 830–844. [Google Scholar] [CrossRef]
- Myneni, R.B.; Boston University, Boston, MA, USA. Personal communication, 2013.
- Boschetti, L.; Roy, D.; Hoffmann, A.A.; Humber, M. MODIS Collection 5.1 Burned Area Product—MCD45,User’s Guide Version 3.0.1. Available online: https://earthdata.nasa.gov/sites/default/files/field/document/MODIS_Burned_Area_Collection51_User_Guide_3.0.pdf (accessed on 26 October 2013).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayr, M.J.; Samimi, C. Comparing the Dry Season In-Situ Leaf Area Index (LAI) Derived from High-Resolution RapidEye Imagery with MODIS LAI in a Namibian Savanna. Remote Sens. 2015, 7, 4834-4857. https://doi.org/10.3390/rs70404834
Mayr MJ, Samimi C. Comparing the Dry Season In-Situ Leaf Area Index (LAI) Derived from High-Resolution RapidEye Imagery with MODIS LAI in a Namibian Savanna. Remote Sensing. 2015; 7(4):4834-4857. https://doi.org/10.3390/rs70404834
Chicago/Turabian StyleMayr, Manuel J., and Cyrus Samimi. 2015. "Comparing the Dry Season In-Situ Leaf Area Index (LAI) Derived from High-Resolution RapidEye Imagery with MODIS LAI in a Namibian Savanna" Remote Sensing 7, no. 4: 4834-4857. https://doi.org/10.3390/rs70404834
APA StyleMayr, M. J., & Samimi, C. (2015). Comparing the Dry Season In-Situ Leaf Area Index (LAI) Derived from High-Resolution RapidEye Imagery with MODIS LAI in a Namibian Savanna. Remote Sensing, 7(4), 4834-4857. https://doi.org/10.3390/rs70404834