Identifying Hydro-Geomorphological Conditions for State Shifts from Bare Tidal Flats to Vegetated Tidal Marshes

 , , , ,
, , , ,
Abstract
1. Introduction
- (1)
- Do stable states in vegetation biomass and elevation co-occur? To answer this question, we tested (1a) whether both vegetation biomass and elevation have a bimodal frequency distribution in intertidal zones; (1b) whether the spatial variation in vegetation biomass as a function of elevation shows an abrupt change from a bare to vegetated state (above a threshold elevation); and (1c) whether the temporal change in vegetation biomass is the largest in areas with an unstable intermediate elevation state;
- (2)
- Does hydrodynamics abruptly change between different states? We tested whether spatial variations in (2a) tidal currents and (2b) waves change abruptly between different biomass and elevation states, in order to identify potential mechanisms causing the two stable states, i.e., the biogeomorphic interactions between elevation, biomass, tidal currents, and waves;
- (3)
- Can we identify the location of state shifts? We examined whether we can identify the locations where shifts from bare state to vegetated state occur, based on integrating the spatially explicit data on elevation, tidal currents, and waves.
2. Materials and Methods
2.1. Study Area
2.2. Materials and Data Preprocessing
2.2.1. Aerial Images and Vegetation Maps
2.2.2. Elevation Data and Intertidal Zones
2.2.3. Tidal Current Velocity from a TELEMAC 2D Model
2.2.4. Wave Orbital Velocity from a SWAN Model
2.3. Data Analysis
2.3.1. Spatial Distribution of Vegetation and Intertidal Elevation
2.3.2. Spatial Variation of Currents and Waves
2.3.3. Identification of the State Shift
3. Results
3.1. Spatial Distribution of Biomass and Elevation
3.1.1. Bimodality in Biomass Distribution and Elevation Distribution
3.1.2. Relations between Variations in Elevation and NDVI
3.1.3. Abrupt Temporal Increase in Biomass in Intermediate Unstable Elevations
3.2. Spatial Variation of Hydrodynamics in between the Different States
3.2.1. Variation of Tidal Current and Wave Orbital Velocity in Relation with NDVI
3.2.2. Variation of Tidal Current and Wave Orbital Velocity in Relation with Elevation
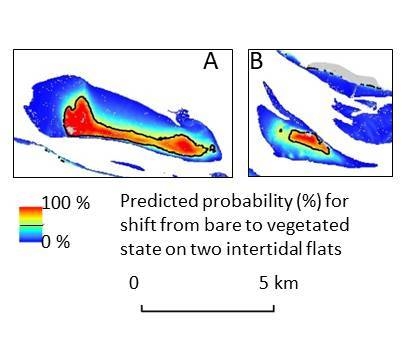
3.3. Identification of State Shift
4. Discussion
4.1. Do Stable States in Vegetation Biomass and Elevation Co-Occur?
4.2. Does Hydrodynamics Abruptly Change between Different States?
4.3. Can We Predict the Location of State Shifts?
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- May, R.M. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 1977, 269, 471–477. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M. Complex systems: Foreseeing tipping points. Nature 2010, 467, 411–412. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Moore, J.C. Predicting tipping points in complex environmental systems. Proc. Natl. Acad. Sci. USA 2018, 115, 635–636. [Google Scholar] [CrossRef]
- D’Alpaos, A.; Lanzoni, S.; Marani, M.; Rinaldo, A. Landscape evolution in tidal embayments: Modeling the interplay of erosion, sedimentation, and vegetation dynamics. J. Geophys. Res. 2007, 112, F01008. [Google Scholar] [CrossRef]
- Temmerman, S.; Bouma, T.J.; van de Koppel, J.; van der Wal, D.D.; de Vries, M.B.; Herman, P.M.J. Vegetation causes channel erosion in a tidal landscape. Geology 2007, 35, 631–634. [Google Scholar] [CrossRef]
- Bouma, T.J.; van Belzen, J.; Balke, T.; van Dalen, J.; Klaassen, P.; Hartog, A.M.; Callaghan, D.P.; Hu, Z.; Stive, M.J.F.; Temmerman, S.; et al. Short-term mudflat dynamics drive long-term cyclic salt marsh dynamics. Limnol. Oceanogr. 2016, 61, 2261–2275. [Google Scholar] [CrossRef]
- van Belzen, J.; van de Koppel, J.V.; Kirwan, M.L.; van der Wal, D.; Herman, P.M.J.; Dakos, V.; Kefi, S.; Scheffer, M.; Guntenspergen, G.R.; Bouma, T.J. Vegetation recovery in tidal marshes reveals critical slowing down under increased inundation. Nat. Commun. 2017, 8, 15811. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Stark, J.; van Oyen, T.; Meire, P.; Temmerman, S. Observations of tidal and storm surge attenuation in a large tidal marsh. Limnol. Oceanogr. 2015, 60, 1371–1381. [Google Scholar] [CrossRef]
- Leonardi, N.; Carnacina, I.; Donatelli, C.; Ganju, N.K.; Plater, A.J.; Schuerch, M.; Temmerman, S. Dynamic interactions between coastal storms and salt marshes: A review. Geomorphology 2018, 301, 92–107. [Google Scholar] [CrossRef]
- Van Coppenolle, R.; Schwarz, C.; Temmerman, S. Contribution of mangroves and salt marshes to nature-based mitigation of coastal flood risks in major deltas of the world. Estuar. Coasts 2018, 41, 1699–1711. [Google Scholar] [CrossRef]
- Vuik, V.; Jonkman, S.N.; Borsje, B.W.; Suzuki, T. Nature-based flood protection: The efficiency of vegetated foreshores for reducing wave loads on coastal dikes. Coast. Eng. 2016, 116, 42–56. [Google Scholar] [CrossRef]
- Möller, I.; Kudella, M.; Rupprecht, F.; Spencer, T.; Paul, M.; van Wesenbeeck, B.K.; Wolters, G.; Jensen, K.; Bouma, T.J.; Miranda-Lange, M.; et al. Wave attenuation over coastal salt marshes under storm surge conditions. Nat. Geosci. 2014, 7, 727–731. [Google Scholar] [CrossRef]
- Schoutens, K.; Heuner, M.; Minden, V.; Schulte Ostermann, T.; Silinski, A.; Belliard, J.-P.; Temmerman, S. How effective are tidal marshes as nature-based shoreline protection throughout seasons? Limnol. Oceanogr. 2019, 64, 1750–1762. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Temmerman, S.; Skeehan, E.E.; Guntenspergen, G.R.; Fagherazzi, S. Overestimation of marsh vulnerability to sea level rise. Nat. Clim. Chang. 2016, 6, 253–260. [Google Scholar] [CrossRef]
- Schuerch, M.; Spencer, T.; Temmerman, S.; Kirwan, M.L.; Wolff, C.; Lincke, D.; McOwen, C.J.; Pickering, M.D.; Reef, R.; Vafeidis, A.T.; et al. Future response of global coastal wetlands to sea-level rise. Nature 2018, 561, 231–234. [Google Scholar] [CrossRef]
- Fagherazzi, S.; Carniello, L.; D’Alpaos, L.; Defina, A. Critical bifurcation of shallow microtidal landforms in tidal flats and salt marshes. Proc. Natl. Acad. Sci. USA 2006, 103, 8337–8341. [Google Scholar] [CrossRef]
- Wang, C.; Temmerman, S. Does biogeomorphic feedback lead to abrupt shifts between alternative landscape states?: An empirical study on intertidal flats and marshes. J. Geophys. Res. Earth Surf. 2013, 118, 229–240. [Google Scholar] [CrossRef]
- Van Wesenbeeck, B.K.; van de Koppel, J.; Herman, P.M.; Bertness, M.D.; van der Wal, D.; Bakker, J.P.; Bouma, T.J. Potential for sudden shifts in transient systems: Distinguishing between local and landscape-scale processes. Ecosystems 2008, 11, 1133–1141. [Google Scholar] [CrossRef]
- Carniello, L.; Defina, A.; D’Alpaos, L. Morphological evolution of the Venice lagoon: Evidence from the past and trend for the future. J. Geophys. Res. Earth 2009, 114, F04002. [Google Scholar] [CrossRef]
- Defina, A.; Carniello, L.; Fagherazzi, S.; D’Alpaos, L. Self-organization of shallow basins in tidal flats and salt marshes. J. Geophys. Res. 2007, 112, F03001. [Google Scholar] [CrossRef]
- Fagherazzi, S.; Palermo, C.; Rulli, M.C.; Carniello, L.; Defina, A. Wind waves in shallow microtidal basins and the dynamic equilibrium of tidal flats. J. Geophys. Res. 2007, 112, F02024. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Murray, A.B. A coupled geomorphic and ecological model of tidal marsh evolution. Proc. Natl. Acad. Sci. USA 2007, 104, 6118–6122. [Google Scholar] [CrossRef]
- Marani, M.; D’Alpaos, A.; Lanzoni, S.; Carniello, L.; Rinaldo, A. Biologically-controlled multiple equilibria of tidal landforms and the fate of the Venice lagoon. Geophys Res. Lett. 2007, 34, L11402. [Google Scholar] [CrossRef]
- Marani, M.; D’Alpaos, A.; Lanzoni, S.; Carniello, L.; Rinaldo, A. The importance of being coupled: Stable states and catastropic shifts in tidal biomorphodynamics. J. Geophys. Res. Earth 2010, 115, F04004. [Google Scholar] [CrossRef]
- Balke, T.; Herman, P.M.J.; Bouma, T.J.; Nilsson, C. Critical transitions in disturbance-driven ecosystems: Identifying windows of opportunity for recovery. J. Ecol. 2014, 102, 700–708. [Google Scholar] [CrossRef]
- Hu, Z.; van Belzen, J.; van der Wal, D.; Balke, T.; Wang, Z.B.; Stive, M.; Bouma, T.J. Windows of opportunity for salt marsh vegetation establishment on bare tidal flats: The importance of temporal and spatial variability in hydrodynamic forcing. J. Geophys. Res. Biogeosci. 2015, 120, 1450–1469. [Google Scholar] [CrossRef]
- Balke, T.; Stock, M.; Jensen, K.; Bouma, T.J.; Kleyer, M. A global analysis of the seaward salt marsh extent: The importance of tidal range. Water Resour. Res. 2016, 52, 3775–3786. [Google Scholar] [CrossRef]
- Willemsen, P.; Borsje, B.W.; Hulscher, S.; Wal, D.; Zhu, Z.; Oteman, B.; Evans, B.; Möller, I.; Bouma, T. Quantifying bed level change at the transition of tidal flat and salt marsh: Can we understand the lateral location of the marsh edge? J. Geophys. Res. Earth Surf. 2018, 123, 2509–2524. [Google Scholar] [CrossRef]
- Friess, D.A.; Krauss, K.W.; Horstman, E.M.; Balke, T.; Bouma, T.J.; Galli, D.; Webb, E.L. Are all intertidal wetlands naturally created equal? Bottlenecks, thresholds and knowledge gaps to mangrove and saltmarsh ecosystems. Biol. Rev. 2012, 87, 346–366. [Google Scholar] [CrossRef]
- Spencer, K.; Harvey, G. Understanding system disturbance and ecosystem services in restored saltmarshes: Integrating physical and biogeochemical processes. Estuar. Coast. Shelf Sci. 2012, 106, 23–32. [Google Scholar] [CrossRef]
- Meire, P.; Ysebaert, T.; Damme, S.V.; Bergh, E.V.d.; Maris, T.; Struyf, E. The Scheldt estuary: A description of a changing ecosystem. Hydrobiologia 2005, 540, 1–11. [Google Scholar] [CrossRef]
- Reitsma, J.M. Toelichting bij de Vegetatiekartering Westerschelde 2004 op Basis Vanfalse Colour-Luchtfoto’s 1:5000/1:10000; Ministerie van verkeer en Waterstaat, Rijksinstituut, Adviesdienst Geo-Informatie & ICT: Den Haag-Delft, The Netherlands, 2006. [Google Scholar]
- Rijkswaterstaat. Kwaliteitsdocument Laseraltimetrie Projectgebied Westerschelde 2011; Ministerie van Verkeer en Waterstaat Rijkswaterstaat: Delft, The Netherlands, 2011. [Google Scholar]
- Lobo, A.; Moloney, K.; Chic, O.; Chiariello, N. Analysis of fine-scale spatial pattern of a grassland from remotely-sensed imagery and field collected data. Landsc. Ecol. 1998, 13, 111–131. [Google Scholar] [CrossRef]
- Price, J.C.; Bausch, W.C. Leaf area index estimation from visible and near-infrared reflectance data. Remote Sens. Environ. 1995, 52, 55–65. [Google Scholar] [CrossRef]
- Tolman, M.E.; Pranger, D.P. Toelichting bij de Vegetatiekartering Westerschelde 2010 op Basis Van False Colour-Luchtfoto’s 1:5000; Ministerie van verkeer en Waterstaat, Rijksinstituut, Adviesdienst Geo-Informatie & ICT: Delft, The Netherlands, 2012. [Google Scholar]
- Alkemade, I.S.W. Kwaliteitsdocument Laseraltimetrie, Projectgebied Westerschelde; Ministerie van Verkeer en Waterstaat, Rijkswaterstaat: Delft, The Netherlands, 2004. [Google Scholar]
- Hervouet, J.-M. Hydrodynamics of Free Surface Flows: Modelling with the Finite Element Method; Wiley Online Library: London, UK, 2007; Volume 360. [Google Scholar]
- Smolders, S.; Ides, S.; Plancke, Y.; Meire, P.; Temmerman, S. Calibrating discharges in a 2D hydrodynamic model of the Scheldt Estuary: Which parameters can be used and what is their sensitivity? In Proceedings of the 10th International Conference on Hydroinformatics, Hamburg, Germany, 14–18 July 2012; p. 8. [Google Scholar]
- Smolders, S.; Plancke, Y.; Ides, S.; Meire, P.; Temmerman, S. Role of intertidal wetlands for tidal and storm tide attenuation along a confined estuary: A model study. Nat. Hazards Earth Syst. Sci. 2015, 3, 3181–3224. [Google Scholar] [CrossRef]
- Booij, N.; Ris, R.C.; Holthuijsen, L.H. A third-generation wave model for coastal regions 1. model description and validation. J. Geophys. Res. 1999, 104, 7649–7666. [Google Scholar] [CrossRef]
- Ris, R.C.; Holthuijsen, L.H.; Booij, N. A third-generation wave model for coastal regions 2. verification. J. Geophys. Res. 1999, 104, 7667–7681. [Google Scholar] [CrossRef]
- Holthuijsen, L.H. Waves in Oceanic and Coastal Waters; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Callaghan, D.P.; Bouma, T.; Klaassen, P.; van der Wal, D.; Stive, M.J.F.; Herman, P.M.J. Hydrodynamic forcing on salt-marsh development: Distinguishing the relative importance of waves and tidal flows. Estuar. Coast. Shelf Sci. 2010, 89, 73–88. [Google Scholar] [CrossRef]
- Holthuijsen, L.H.; Herman, A.; Booij, N. Phase-decoupled refraction-diffraction for spectral wave models. Coast. Eng. 2003, 49, 291–305. [Google Scholar] [CrossRef]
- Cao, H.B.; Zhu, Z.C.; Balke, T.; Zhang, L.Q.; Bouma, T.J. Effects of sediment disturbance regimes on Spartina seedling establishment: Implications for salt marsh creation and restoration. Limnol. Oceanogr. 2017, 62, 2348–2357. [Google Scholar] [CrossRef]
- Silinski, A.; van Belzen, J.; Fransen, E.; Bouma, T.J.; Troch, P.; Meire, P.; Temmerman, S. Quantifying critical conditions for seaward expansion of tidal marshes: A transplantation experiment. Estuar. Coast. Shelf Sci. 2016, 169, 227–237. [Google Scholar] [CrossRef]
- Van Wesenbeeck, B.K.; van de Koppel, J.; Herman, P.M.J.; Bouma, T.J. Does scale-dependent feedback explain spatial complexity in salt-marsh ecosystems? Oikos 2008, 117, 152–159. [Google Scholar] [CrossRef]
- Balke, T.; Bouma, T.J.; Horstman, E.M.; Webb, E.L.; Erftemeijer, P.L.A.; Herman, P.M.J. Windows of opportunity: Thresholds to mangrove seedling establishment on tidal flats. Mar. Ecol. Prog. Ser. 2011, 440, 1–9. [Google Scholar] [CrossRef]
- Davy, A.J.; Brown, M.J.H.; Mossman, H.L.; Grant, A. Colonization of a newly developing salt marsh: Disentangling independent effects of elevation and redox potential on halophytes. J. Ecol. 2011, 99, 1350–1357. [Google Scholar] [CrossRef]
- McGlathery, K.J.; Reidenbach, M.A.; D’Odorico, P.; Fagherazzi, S.; Pace, M.L.; Porter, J.H. Nonlinear dynamics and alternative stable states in shallow coastal systems. Oceanography 2013, 26, 220–231. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Guntenspergen, G.R. Influence of tidal range on the stability of coastal marshland. J. Geophys. Res. 2010, 115, F02009. [Google Scholar] [CrossRef]
- D’Alpaos, A. The mutual influence of biotic and abiotic components on the long-term ecomorphodynamic evolution of salt-marsh ecosystems. Geomorphology 2011, 126, 269–278. [Google Scholar] [CrossRef]
- Bruno, J.F. Facilitation of cobble beach plant communities through habitat modification by Spartina alterniflora. Ecology 2000, 81, 1179–1192. [Google Scholar] [CrossRef]
- Van de Koppel, J.; Herman, P.M.J.; Thoolen, P.; Heip, C.H.R. Do alternate stable states occur in natural ecosystems? Evidence from a tidal flat. Ecology 2001, 82, 3449–3461. [Google Scholar] [CrossRef]
- Mudd, S.M.; D’Alpaos, A.; Morris, J.T. How does vegetation affect sedimentation on tidal marshes? Investigating particle capture and hydrodynamic controls on biologically mediated sedimentation. J. Geophys. Res. Earth Surf. 2010, 115, F03029. [Google Scholar] [CrossRef]
- Vandenbruwaene, W.; Temmerman, S.; Bouma, T.J.; Klaassen, P.C.; De Vries, M.B.; Callaghan, D.P.; Van Steeg, P.; Dekker, F.; Van Duren, L.A.; Martini, E.; et al. Flow interaction with dynamic vegetation patches: Implications for bio-geomorphic evolution of a tidal landscape. J. Geophys. Res. Earth 2011, 116, F01008. [Google Scholar] [CrossRef]
- Pethick, J.S. Long-term accretion rates on tidal salt marshes. J. Sediment. Petrol. 1981, 51, 571–577. [Google Scholar] [CrossRef]
- Temmerman, S.; Govers, G.; Meire, P.; Wartel, S. Modelling long-term tidal marsh growth under changing tidal conditions and suspended sediment concentrations, Scheldt estuary, Belgium. Mar. Geol. 2003, 193, 151–169. [Google Scholar] [CrossRef]
- Neumeier, U.; Amos, C.L. The influence of vegetation on turbulence and flow velocities in European salt-marshes. Sedimentology 2006, 53, 259–277. [Google Scholar] [CrossRef]
- Yang, S.L.; Shi, B.W.; Bouma, T.J.; Ysebaert, T.; Luo, X.X. Wave attenuation at a salt marsh margin: A case study of an exposed coast on the Yangtze Estuary. Estuaries Coasts 2012, 35, 169–182. [Google Scholar] [CrossRef]
- Bouma, T.J.; van Duren, L.A.; Temmerman, S.; Claverie, T.; Blanco-Garcia, A.; Ysebaert, T.; Herman, P.M.J. Spatial flow and sedimentation patterns within patches of epibenthic structures: Combining field, flume and modelling experiments. Cont. Shelf Res. 2007, 27, 1020–1045. [Google Scholar] [CrossRef]
- D’Alpaos, A.; Da Lio, C.; Marani, M. Biogeomorphology of tidal landforms: Physical and biological processes shaping the tidal landscape. Ecohydrology 2012, 5, 550–562. [Google Scholar] [CrossRef]
- Burd, F.; Clifton, J.; Murphy, B. Sites of Historical Sea Defence Failure. Phase II study; Institute of Estuarine and Coastal Studies: Hull, UK, 1994.
- Blott, S.J.; Pye, K. Application of lidar digital terrain modelling to predict intertidal habitat development at a managed retreat site: Abbotts Hall, Essex, UK. Earth Surf. Process. Landf. 2004, 29, 893–905. [Google Scholar] [CrossRef]
- Bouma, T.J.; Friedrichs, M.; Klaassen, P.; van Wesenbeeck, B.K.; Brun, F.G.; Temmerman, S.; van Katwijk, M.M.; Graf, G.; Herman, P.M.J. Effects of shoot stiffness, shoot size and current velocity on scouring sediment from around seedlings and propagules. Mar. Ecol. Prog. Ser. 2009, 388, 293–297. [Google Scholar] [CrossRef]
- Acacio, V.; Holmgren, M.; Rego, F.; Moreira, F.; Mohren, G.M.J. Are drought and wildfires turning Mediterranean cork oak forests into persistent shrublands? Agrofor. Syst. 2009, 76, 389–400. [Google Scholar] [CrossRef]
- Hirota, M.; Holmgren, M.; van Nes, E.H.; Scheffer, M. Global resilience of tropical forest and savanna to critical transitions. Science 2011, 334, 232–235. [Google Scholar] [CrossRef]
- Kachergis, E.; Rocca, M.E.; Fernandez-Gimenez, M.E. Indicators of ecosystem function identify alternate states in the Sagebrush steppe. Ecol. Appl. 2011, 21, 2781–2792. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Wang, Z.; Wang, C.; Mao, D.; Zhang, Y. A new vegetation index to detect periodically submerged Mangrove forest using single-tide sentinel-2 imagery. Remote Sens. 2019, 11, 2043. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Z.; Zhang, Y.; Mao, D.; Wang, C. Monitoring loss and recovery of mangrove forests during 42 years: The achievements of mangrove conservation in China. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 535–545. [Google Scholar] [CrossRef]
- Balke, T.; Friess, D.A. Geomorphic knowledge for mangrove restoration: A pan-tropical categorization. Earth Surf. Process. Landf. 2016, 41, 231–239. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grid | Origin | Rotation | Length | Grid Spacing | |||
|---|---|---|---|---|---|---|---|
| E [m] | N [m] | [°] | x [km] | y [km] | x [km] | y [km] | |
| 0 | −113,200 | 271,906 | 38.303 | 293 | 148 | 2.9 | 3 |
| 1 | 16,023 | 376,356 | 352.014 | 62.4 | 19.5 | 0.19 | 0.19 |
| 2 | 25,226 | 375,297 | 352.014 | 52 | 16.5 | 0.1 | 0.1 |
| 3 | 35,934 | 376,418 | 326.089 | 5.69 | 2.4 | 0.04 | 0.02 |
| 4 | 45,632 | 376,523 | 8.787 | 6.84 | 3.45 | 0.05 | 0.02 |
| 5 | 53,603 | 374,887 | 37.622 | 4.64 | 2.11 | 0.04 | 0.02 |
| 6 | 29,873 | 378,285 | 347.418 | 8.19 | 4.04 | 0.04 | 0.04 |
| 7 | 37,522 | 378,999 | 327.858 | 12.4 | 3.3 | 0.04 | 0.04 |
| 8 | 54,806 | 377,049 | 69.927 | 8.08 | 2.99 | 0.04 | 0.04 |
| 9 | 55,874 | 380,301 | 350.768 | 4.8 | 3.67 | 0.04 | 0.04 |
| 10 | 61,203 | 377,403 | 347.418 | 8.2 | 3.33 | 0.04 | 0.04 |
| 11 | 68,404 | 369,852 | 34.401 | 10.5 | 6.93 | 0.04 | 0.02 |
| No. | Parameters | Equations | Nagelkerke R2 | Overall Correct Percentage | Correct Percentage for New Marshes | Correct Percentage for Stable Bare Flats |
|---|---|---|---|---|---|---|
| 1 | Elevation (E) | 0.304 | 85.4 | 22.4 | 97.7 | |
| 2 | Current velocity (C) | 0.320 | 84.2 | 23.1 | 96.1 | |
| 3 | Wave velocity (W) | 0.168 | 84.7 | 11.2 | 99.1 | |
| 4 | Elevation, current velocity | 0.435 | 88.8 | 47.3 | 96.9 | |
| 5 | Elevation, wave velocity | 0.359 | 86.1 | 34.5 | 96.2 | |
| 6 | Current velocity, wave velocity | 0.412 | 87.1 | 38.4 | 96.6 | |
| 7 | Elevation, current velocity, wave velocity | 0.475 | 89.0 | 50.1 | 96.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Smolders, S.; Callaghan, D.P.; van Belzen, J.; Bouma, T.J.; Hu, Z.; Wen, Q.; Temmerman, S. Identifying Hydro-Geomorphological Conditions for State Shifts from Bare Tidal Flats to Vegetated Tidal Marshes. Remote Sens. 2020, 12, 2316. https://doi.org/10.3390/rs12142316
Wang C, Smolders S, Callaghan DP, van Belzen J, Bouma TJ, Hu Z, Wen Q, Temmerman S. Identifying Hydro-Geomorphological Conditions for State Shifts from Bare Tidal Flats to Vegetated Tidal Marshes. Remote Sensing. 2020; 12(14):2316. https://doi.org/10.3390/rs12142316
Chicago/Turabian StyleWang, Chen, Sven Smolders, David P. Callaghan, Jim van Belzen, Tjeerd J. Bouma, Zhan Hu, Qingke Wen, and Stijn Temmerman. 2020. "Identifying Hydro-Geomorphological Conditions for State Shifts from Bare Tidal Flats to Vegetated Tidal Marshes" Remote Sensing 12, no. 14: 2316. https://doi.org/10.3390/rs12142316
APA StyleWang, C., Smolders, S., Callaghan, D. P., van Belzen, J., Bouma, T. J., Hu, Z., Wen, Q., & Temmerman, S. (2020). Identifying Hydro-Geomorphological Conditions for State Shifts from Bare Tidal Flats to Vegetated Tidal Marshes. Remote Sensing, 12(14), 2316. https://doi.org/10.3390/rs12142316